
Marine-Sulfated Polysaccharides Extracts Exhibit Contrasted Time-Dependent Immunomodulatory and Antiviral Properties on Porcine Monocytes and Alveolar Macrophages
 ,
, 
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Monocytes
2.2. Alveolar Macrophages
2.3. Virus Productions
2.4. Marine-Sulfated Polysaccharides
2.5. Treatments
2.6. Marine-Sulfated Polysaccharide Treatment of the Porcine Cells and Virus Infections
2.7. RNA Extractions and Reverse Transcription (RT)-Quantitative Polymerase Chain Reaction (qPCR)
2.8. Statistics
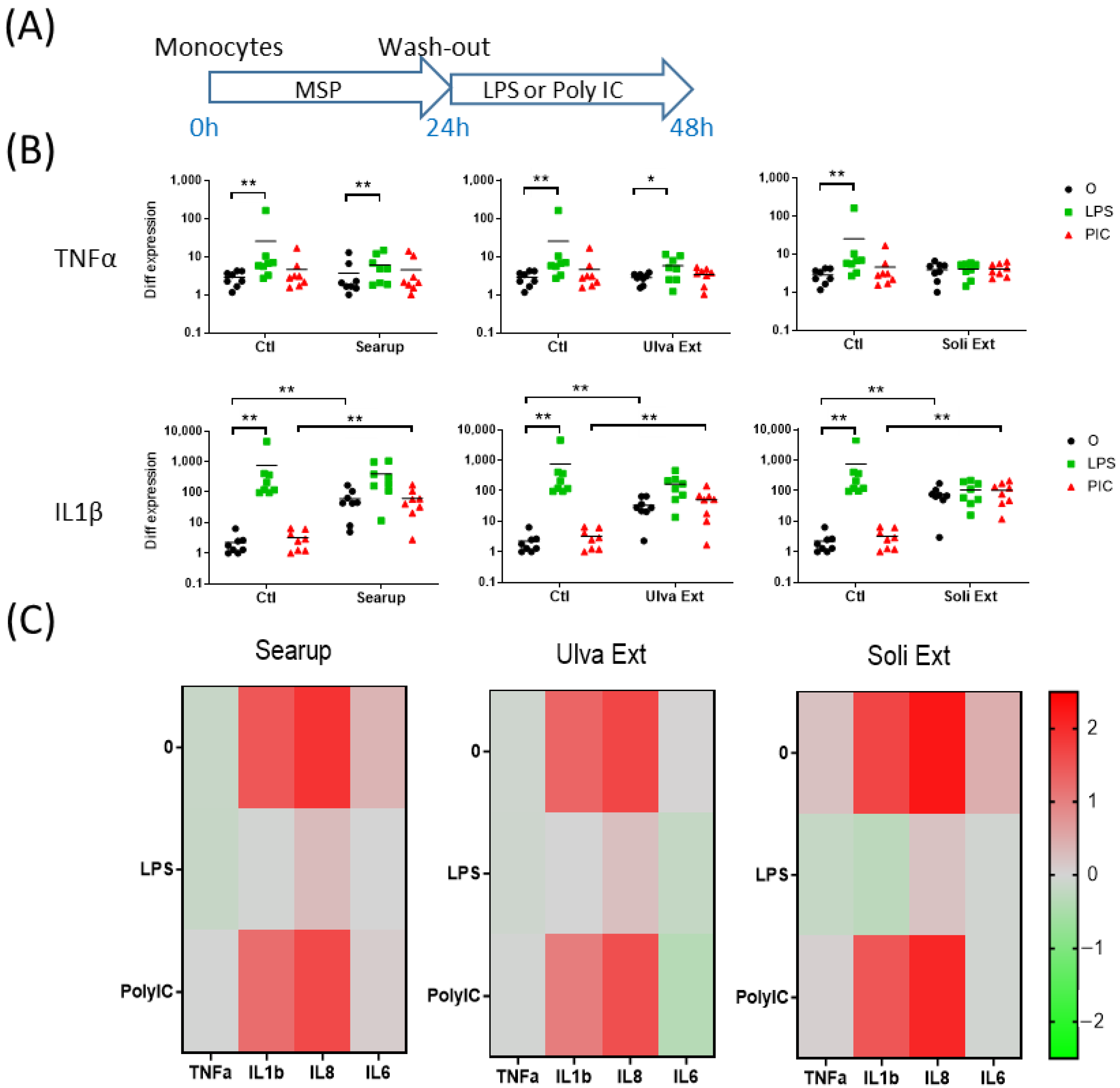
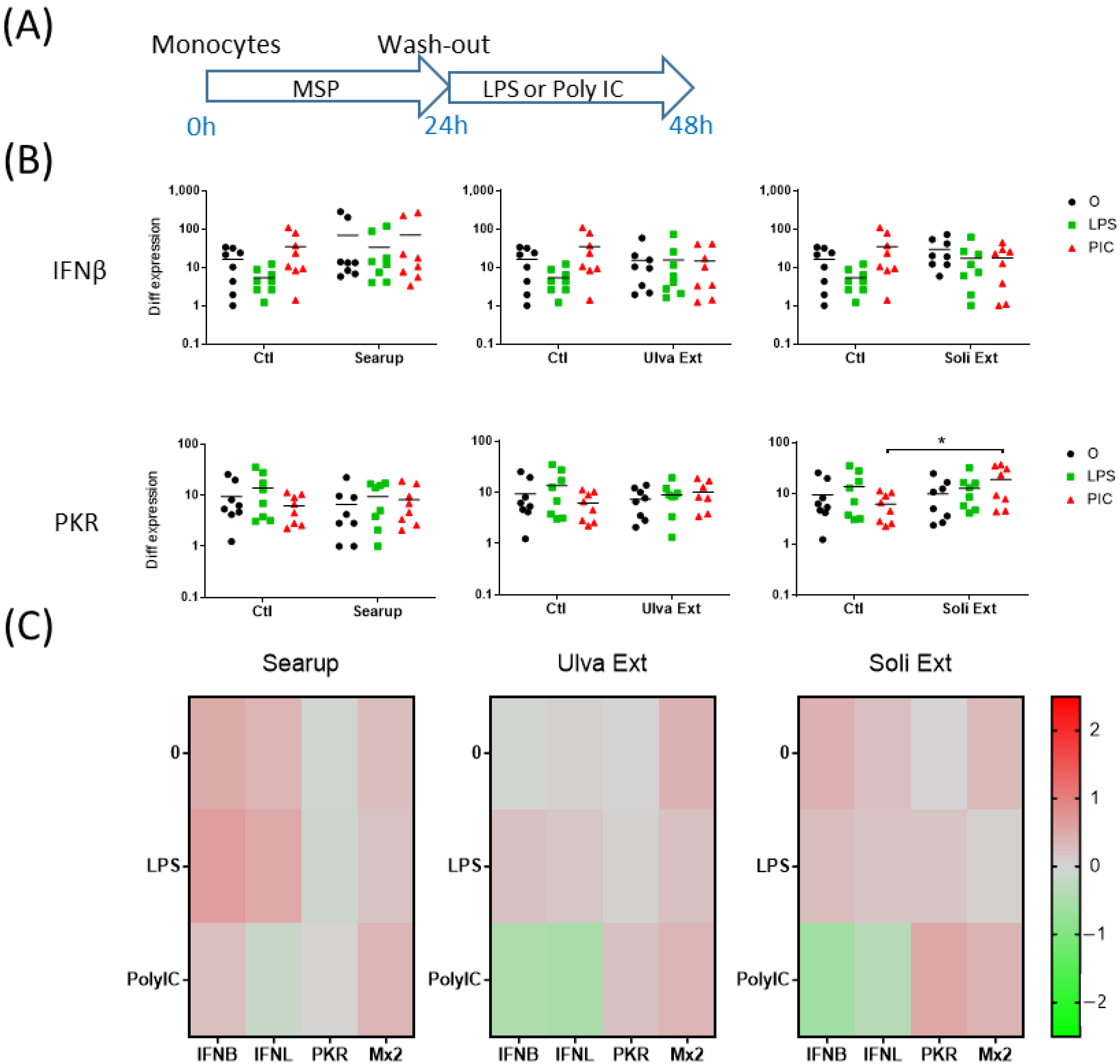
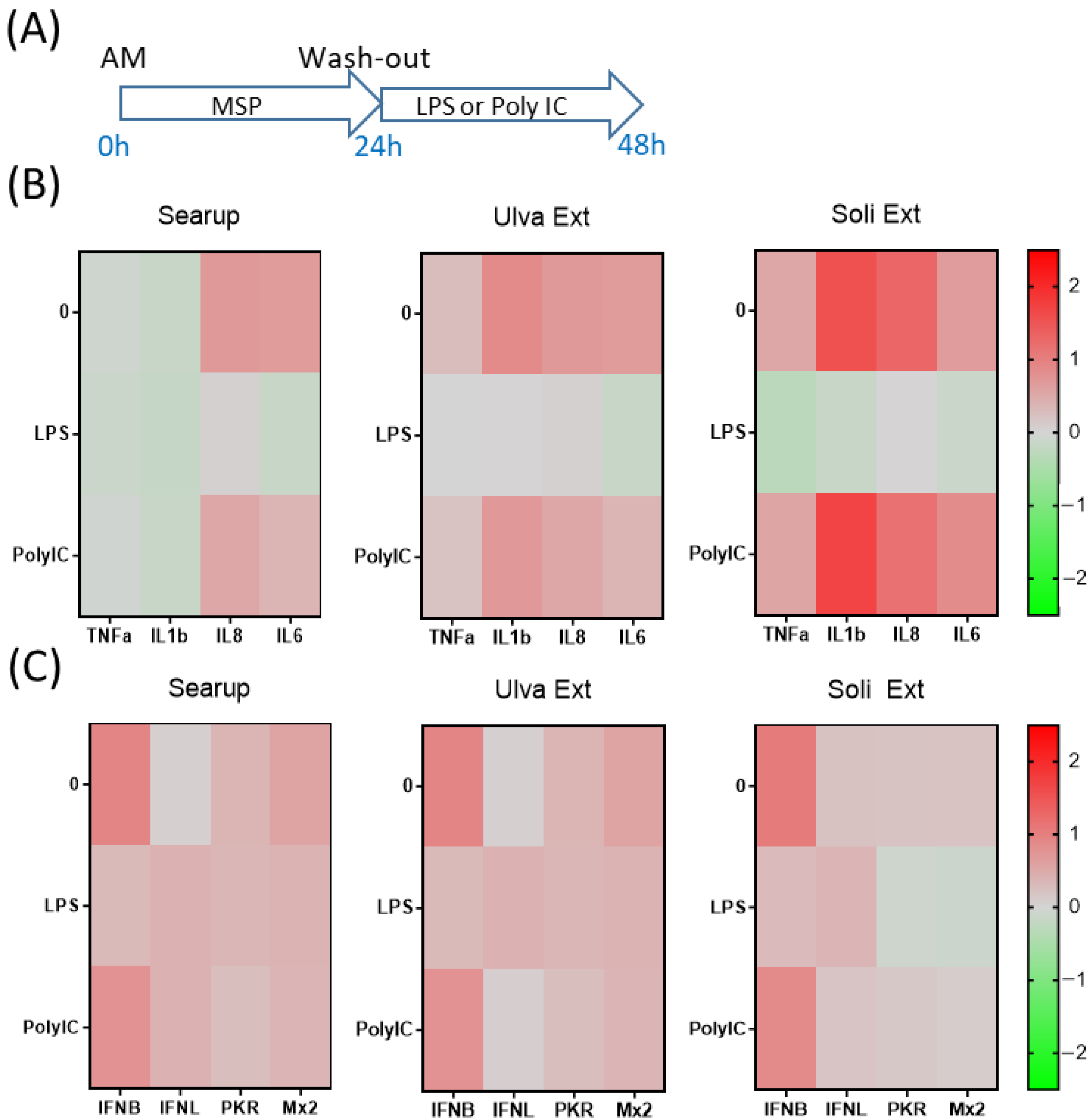
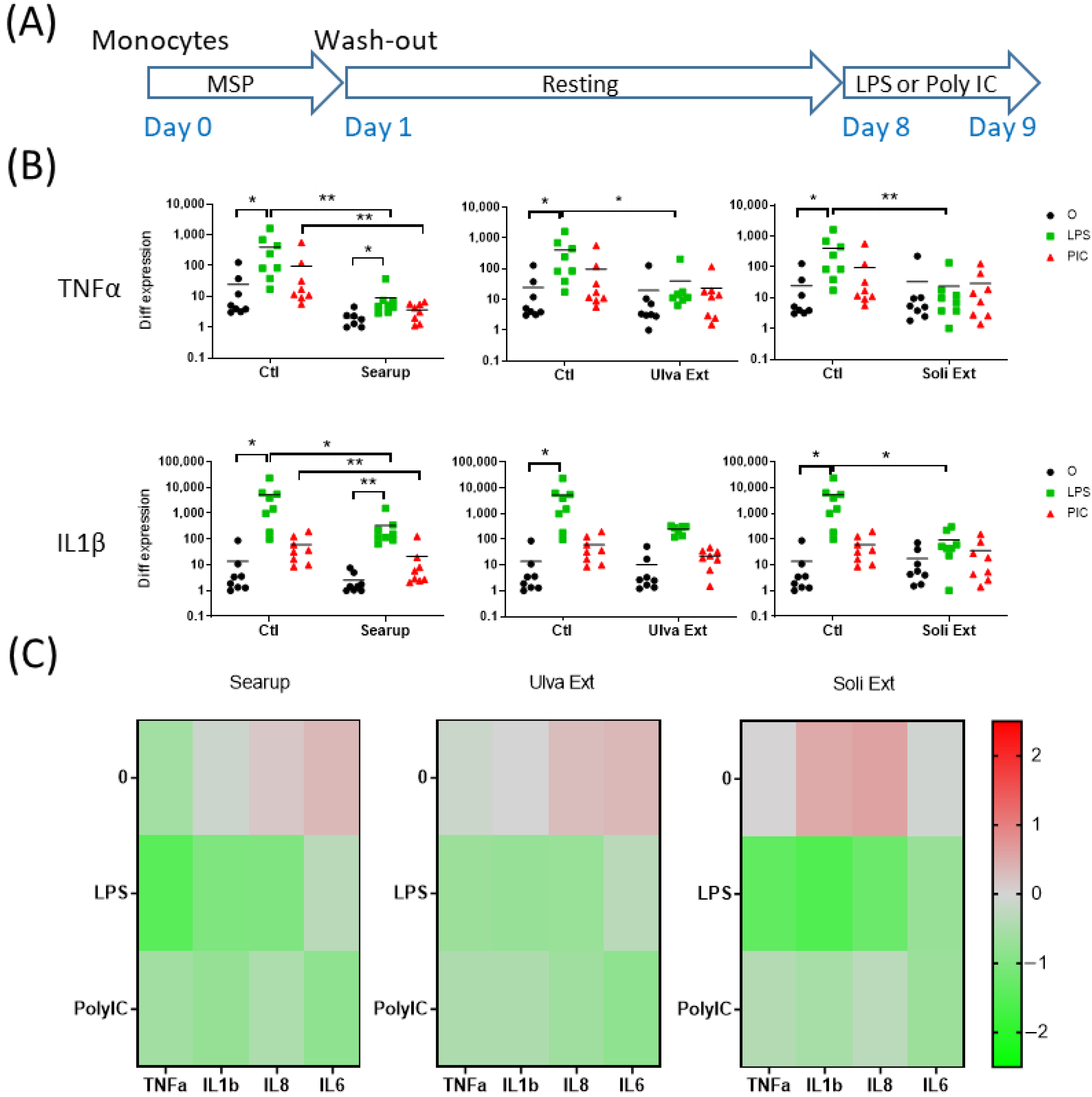
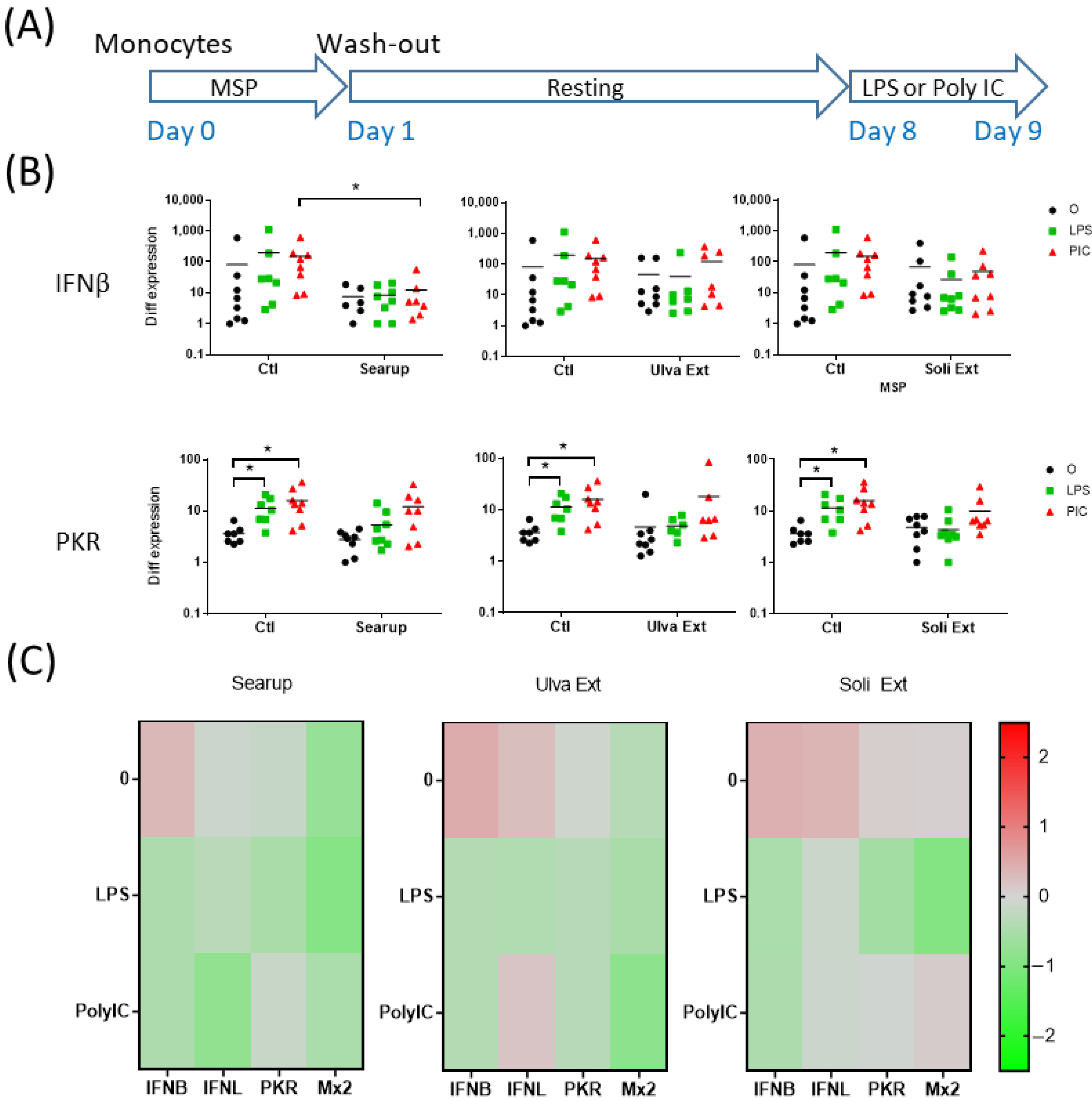
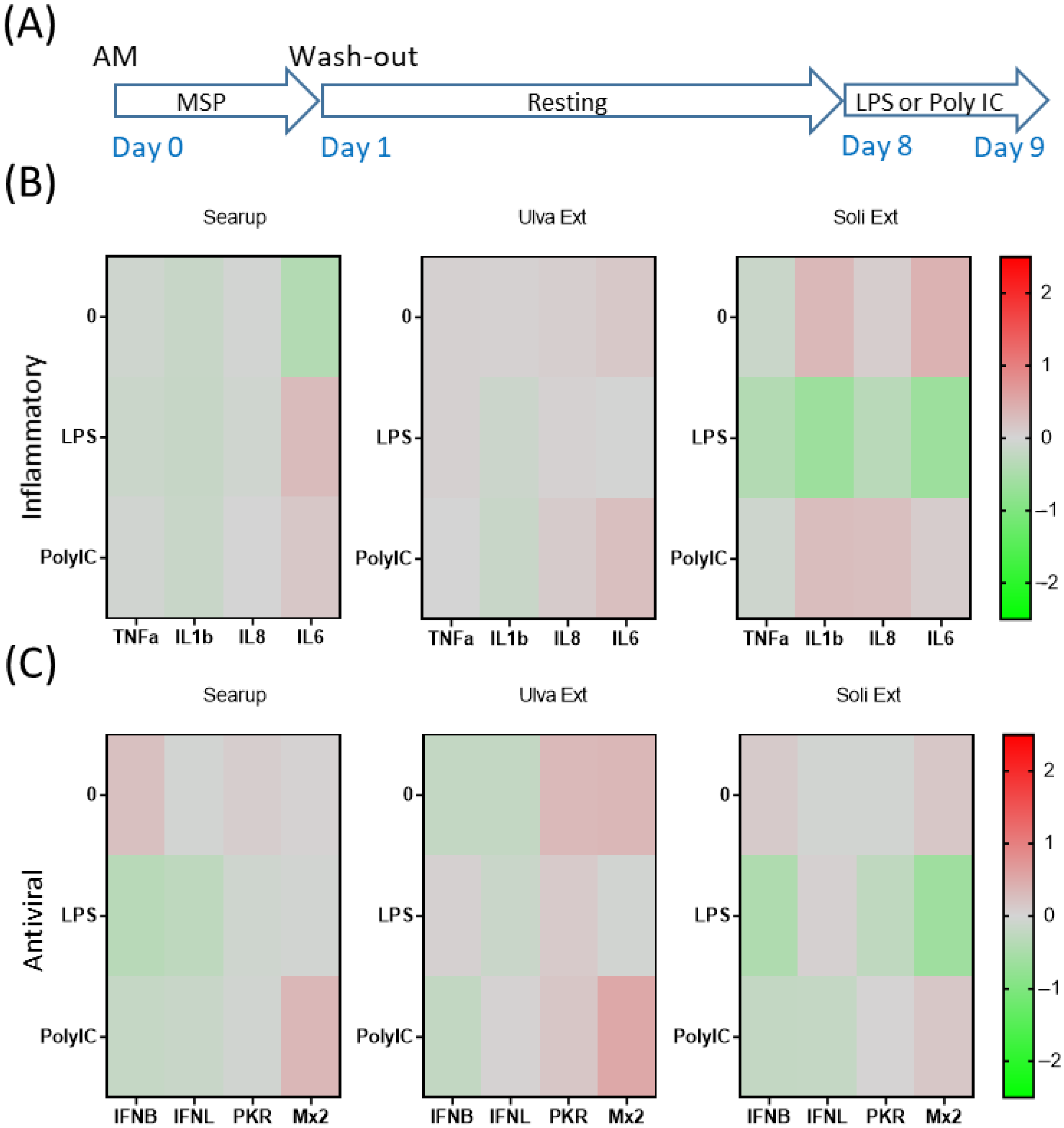
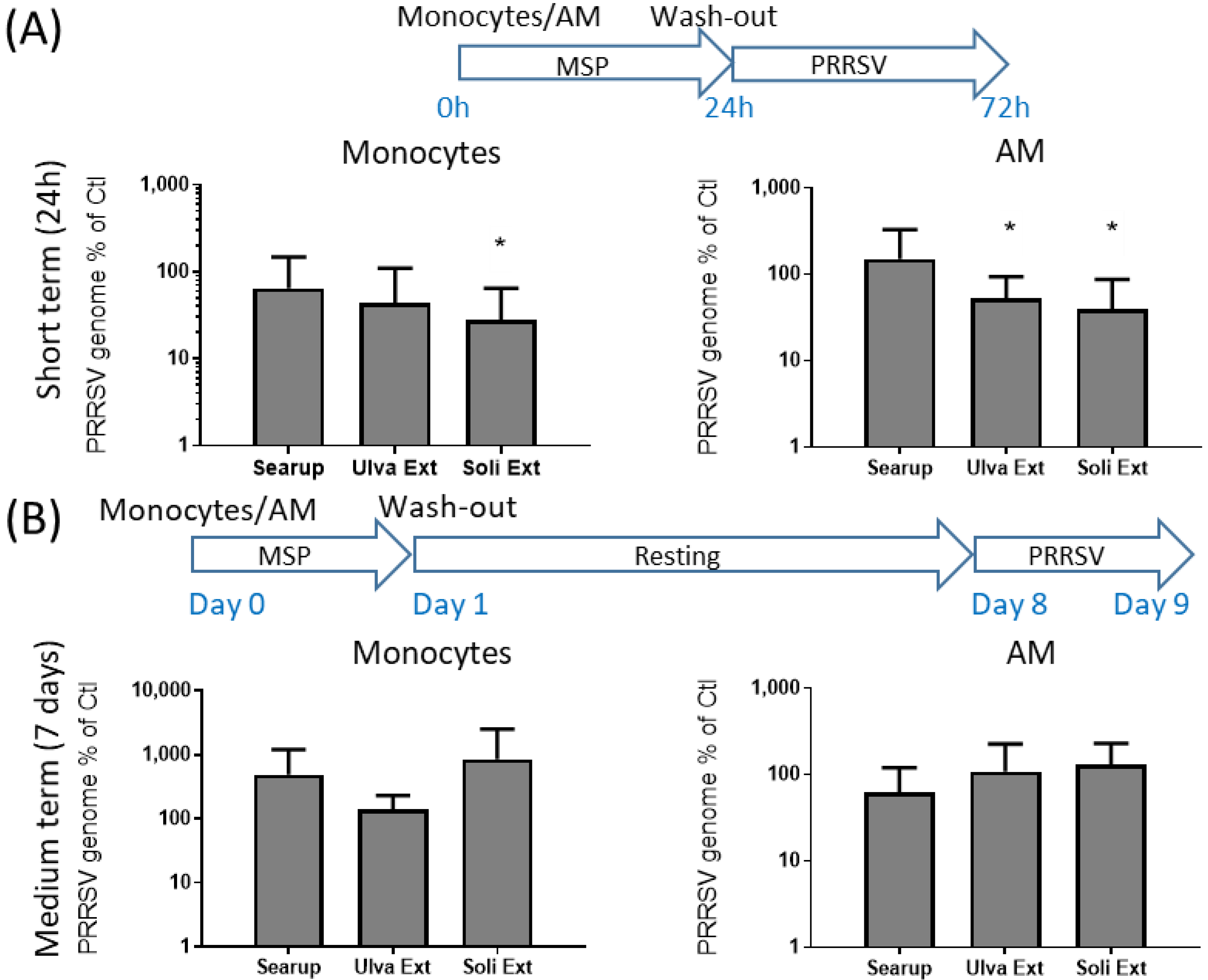
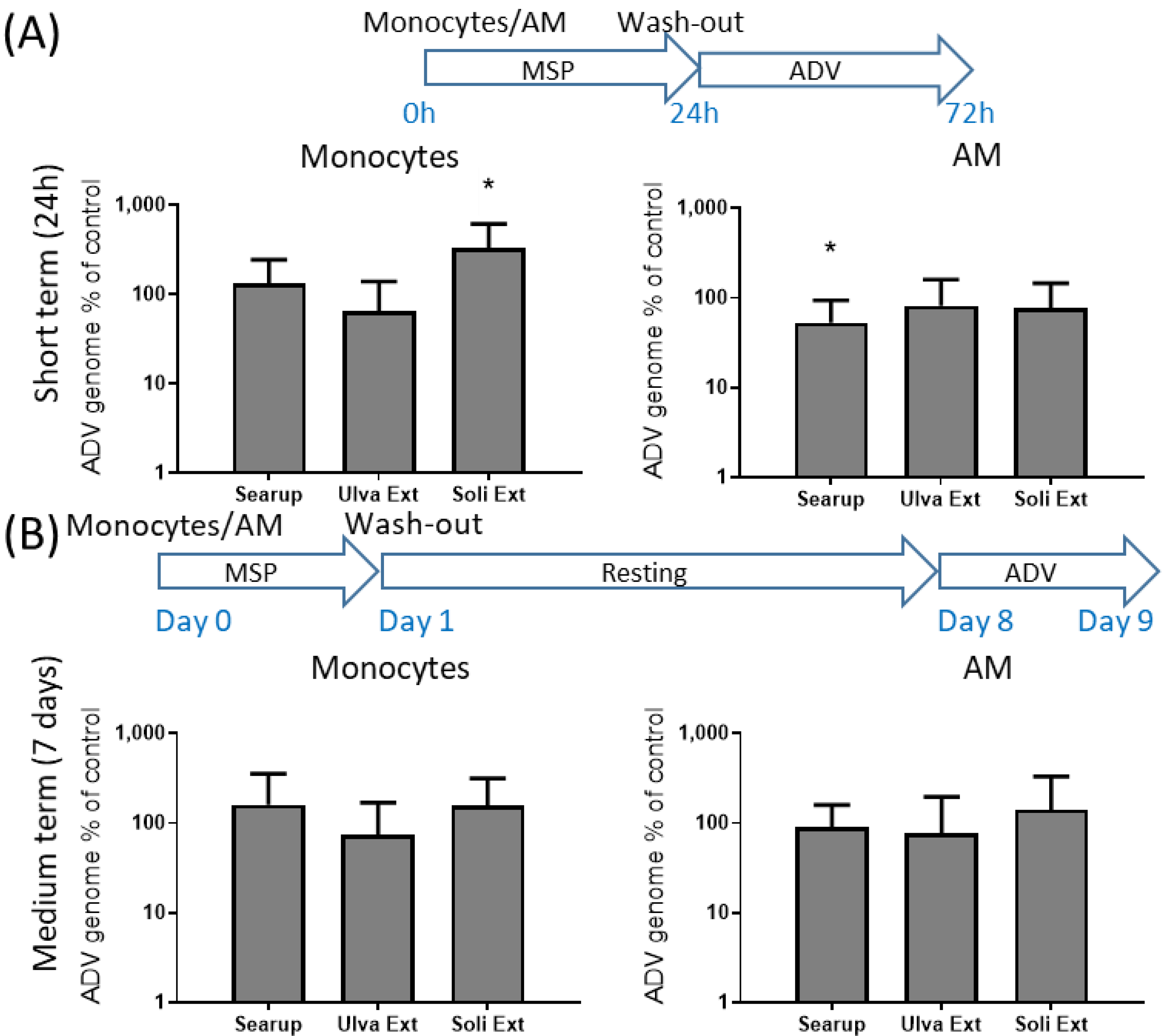
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saade, G.; Deblanc, C.; Bougon, J.; Marois-Créhan, C.; Fablet, C.; Auray, G.; Belloc, C.; Leblanc-Maridor, M.; Gagnon, C.A.; Zhu, J.; et al. Coinfections and their molecular consequences in the porcine respiratory tract. Veter Res. 2020, 51, 80. [Google Scholar] [CrossRef] [PubMed]
- Ye, G.; Liu, H.; Zhou, Q.; Liu, X.; Huang, L.; Weng, C. A tug of war: Pseudorabies virus and host antiviral innate immunity. Viruses 2022, 14, 547. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, R.; Qiao, S.; Chen, X.-X.; Deng, R.; Zhang, G. Porcine sialoadhesin suppresses type I interferon production to support porcine reproductive and respiratory syndrome virus infection. Veter Res. 2020, 51, 18. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, X.; Bai, J.; Sun, Y.; Nauwynck, H.; Wang, X.; Yang, Y.; Jiang, P. A new long noncoding RNA, mahat, inhibits replication of porcine reproductive and respiratory syndrome virus by recruiting DDX6 to bind to ZNF34 and promote an innate immune response. J. Virol. 2022, e01154-22. [Google Scholar] [CrossRef]
- Li, S.; Zhang, X.; Yao, Y.; Zhu, Y.; Zheng, X.; Liu, F.; Feng, W. Inducible miR-150 inhibits porcine reproductive and respiratory syndrome virus replication by targeting viral genome and suppressor of cytokine signaling 1. Viruses 2022, 14, 1485. [Google Scholar] [CrossRef]
- Ghosh, C.; Sarkar, P.; Issa, R.; Haldar, J. Alternatives to conventional antibiotics in the era of antimicrobial resistance. Trends Microbiol. 2019, 27, 323–338. [Google Scholar] [CrossRef]
- Wijesekara, I.; Pangestuti, R.; Kim, S.-K. Biological activities and potential health benefits of sulfated polysaccharides derived from marine algae. Carbohydr. Polym. 2011, 84, 14–21. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Guerrero, A.J.; Rodríguez, A.D.; Taglialatela-Scafati, O.; Nakamura, F.; Fusetani, N. Marine pharmacology in 2014–2015: Marine compounds with antibacterial, antidiabetic, antifungal, anti-inflammatory, antiprotozoal, antituberculosis, antiviral, and anthelmintic activities; affecting the immune and nervous systems, and other miscellaneous mechanisms of action. Mar. Drugs 2020, 18, 5. [Google Scholar] [CrossRef]
- Raposo, M.F.D.J.; de Morais, R.M.S.C.; de Morais, A.M.M.B. Health applications of bioactive compounds from marine microalgae. Life Sci. 2013, 93, 479–486. [Google Scholar] [CrossRef]
- Chojnacka, K. Biologically active compounds in seaweed extracts—The prospects for the application. Open Conf. Proc. J. 2012, 3, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Bussy, F.; Matthieu, L.G.; Salmon, H.; Delaval, J.; Berri, M.; Pi, N.C. Immunomodulating effect of a seaweed extract from lva armoricana in pig: Specific IgG and total IgA in colostrum, milk, and blood. Veter Anim. Sci. 2019, 7, 100051. [Google Scholar] [CrossRef]
- Apostolova, E.; Lukova, P.; Baldzhieva, A.; Katsarov, P.; Nikolova, M.; Iliev, I.; Peychev, L.; Trica, B.; Oancea, F.; Delattre, C.; et al. Immunomodulatory and anti-inflammatory effects of fucoidan: A review. Polymers 2020, 12, 2338. [Google Scholar] [CrossRef]
- Corino, C.; Modina, S.C.; Di Giancamillo, A.; Chiapparini, S.; Rossi, R. Seaweeds in pig nutrition. Animals 2019, 9, 1126. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, W.A.J.P.; Jeon, Y.-J. Biological activities and potential cosmeceutical applications of bioactive components from brown seaweeds: A review. Phytochem. Rev. 2011, 10, 431–443. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Ferreira, L.G.; Noseda, M.D.; Gonçalves, A.G.; Ducatti, D.R.B.; Fujii, M.T.; Duarte, M.E.R. Chemical structure of the complex pyruvylated and sulfated agaran from the red seaweed palisada flagellifera (Ceramiales, Rhodophyta). Carbohydr. Res. 2012, 347, 83–94. [Google Scholar] [CrossRef]
- Berri, M.; Olivier, M.; Holbert, S.; Dupont, J.; Demais, H.; Le Goff, M.; Collen, P.N. Ulvan from Ulva armoricana (Chlorophyta) activates the PI3K/Akt signalling pathway via TLR4 to induce intestinal cytokine production. Algal Res. 2017, 28, 39–47. [Google Scholar] [CrossRef]
- Berri, M.; Slugocki, C.; Olivier, M.; Helloin, E.; Jacques, I.; Salmon, H.; Demais, H.; Le Goff, M.; Collen, P.N. Marine-sulfated polysaccharides extract of Ulva armoricana green algae exhibits an antimicrobial activity and stimulates cytokine expression by intestinal epithelial cells. J. Appl. Phycol. 2016, 28, 2999–3008. [Google Scholar] [CrossRef]
- Lahaye, M.; Robic, A. Structure and function properties of Ulvan, a polysaccharide from green seaweeds. Biomacromolecules 2007, 8, 1765–1774. [Google Scholar] [CrossRef]
- Guriec, N.; Bussy, F.; Gouin, C.; Mathiaud, O.; Le Goff, M.; Delarue, J.; Nyvall Collén, P. Activation of heterophils and monocytes in chicken with a formulation containing a seaweed extract from Ulva armoricana. Open Access J. Veter Sci. Res. 2021, 6, 1–7. [Google Scholar] [CrossRef]
- Guriec, N.; Bussy, F.; Gouin, C.; Mathiaud, O.; Quero, B.; Le Goff, M.; Collén, P.N. Ulvan activates chicken heterophils and monocytes through toll-like receptor 2 and toll-like receptor 4. Front. Immunol. 2018, 9, 1–19. [Google Scholar] [CrossRef]
- Netea, M.G.; Domínguez-Andrés, J.; Barreiro, L.B.; Chavakis, T.; Divangahi, M.; Fuchs, E.; Joosten, L.A.B.; Van Der Meer, J.W.M.; Mhlanga, M.M.; Mulder, W.J.M.; et al. Defining trained immunity and its role in health and disease. Nat. Rev. Immunol. 2020, 20, 375–388. [Google Scholar] [CrossRef]
- Divangahi, M.; Aaby, P.; Khader, S.A.; Barreiro, L.B.; Bekkering, S.; Chavakis, T.; van Crevel, R.; Curtis, N.; DiNardo, A.R.; Dominguez-Andres, J.; et al. Trained immunity, tolerance, priming and differentiation: Distinct immunological processes. Nat. Immunol. 2021, 22, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Byrne, K.A.; Tuggle, C.K.; Loving, C.L. Differential induction of innate memory in porcine monocytes by β-glucan or bacillus calmette-guerin. Innate Immun. 2020, 27, 448–460. [Google Scholar] [CrossRef]
- Ferrari, M.; Scalvini, A.; Losio, M.; Corradi, A.; Soncini, M.; Bignotti, E.; Milanesi, E.; Ajmone-Marsan, P.; Barlati, S.; Bellotti, D.; et al. Establishment and characterization of two new pig cell lines for use in virological diagnostic laboratories. J. Virol. Methods 2002, 107, 205–212. [Google Scholar] [CrossRef]
- Hughey, J.R.; Gabrielson, P.W.; Maggs, C.A.; Mineur, F. Genomic analysis of the lectotype specimens of European Ulva rigida and Ulva lacinulata (Ulvaceae, Chlorophyta) reveals the ongoing misapplication of names. Eur. J. Phycol. 2021, 57, 143–153. [Google Scholar] [CrossRef]
- Fort, A.; McHale, M.; Cascella, K.; Potin, P.; Perrineau, M.-M.; Kerrison, P.D.; da Costa, E.; Calado, R.; Domingues, M.D.R.; Azevedo, I.C.; et al. Exhaustive reanalysis of barcode sequences from public repositories highlights ongoing misidentifications and impacts taxa diversity and distribution. Mol. Ecol. Resour. 2021, 22, 86–101. [Google Scholar] [CrossRef]
- Ifrim, D.C.; Quintin, J.; Joosten, L.A.B.; Jacobs, C.; Jansen, T.; Jacobs, L.; Gow, N.A.R.; Williams, D.L.; van der Meer, J.W.M.; Netea, M.G. Trained immunity or tolerance: Opposing functional programs induced in human monocytes after engagement of various pattern recognition receptors. Clin. Vaccine Immunol. 2014, 21, 534–545. [Google Scholar] [CrossRef]
- Bordet, E.; Blanc, F.; Tiret, M.; Crisci, E.; Bouguyon, E.; Renson, P.; Maisonnasse, P.; Bourge, M.; Leplat, J.-J.; Giuffra, E.; et al. Porcine reproductive and respiratory syndrome virus type 1.3 lena triggers conventional dendritic cells 1 activation and t helper 1 immune response without infecting dendritic cells. Front. Immunol. 2018, 9, 2299. [Google Scholar] [CrossRef]
- Delgado-Ortega, M.; Melo, S.; Punyadarsaniya, D.; Ramé, C.; Olivier, M.; Soubieux, D.; Marc, D.; Simon, G.; Herrler, G.; Berri, M.; et al. Innate immune response to a H3N2 subtype swine influenza virus in newborn porcine trachea cells, alveolar macrophages, and precision-cut lung slices. Veter Res. 2014, 45, 42. [Google Scholar] [CrossRef] [Green Version]
- Dobrescu, I.; Levast, B.; Lai, K.; Delgado-Ortega, M.; Walker, S.; Banman, S.; Townsend, H.; Simon, G.; Zhou, Y.; Gerdts, V.; et al. In vitro and ex vivo analyses of co-infections with swine influenza and porcine reproductive and respiratory syndrome viruses. Veter Microbiol. 2013, 169, 18–32. [Google Scholar] [CrossRef]
- Maisonnasse, P.; Bouguyon, E.; Piton, G.; Ezquerra, A.; Urien, C.; Deloizy, C.; Bourge, M.; Leplat, J.-J.; Simon, G.; Chevalier, C.; et al. The respiratory DC/macrophage network at steady-state and upon influenza infection in the swine biomedical model. Mucosal Immunol. 2016, 9, 835–849. [Google Scholar] [CrossRef]
- Renson, P.; Deblanc, C.; Bougon, J.; Le Dimna, M.; Gorin, S.; Mahé, S.; Barbier, N.; Paboeuf, F.; Simon, G.; Bourry, O. Concomitant swine influenza A virus infection alters PRRSV1 MLV viremia in piglets but does not interfere with vaccine protection in experimental conditions. Vaccines 2021, 9, 356. [Google Scholar] [CrossRef]
- Saade, G.; Ménard, D.; Hervet, C.; Renson, P.; Hue, E.; Zhu, J.; Dubreil, L.; Paillot, R.; Pronost, S.; Bourry, O.; et al. Porcine reproductive and respiratory syndrome virus interferes with swine influenza A virus infection of epithelial cells. Vaccines 2020, 8, 508. [Google Scholar] [CrossRef]
- Rose, N.; Renson, P.; Andraud, M.F.; Paboeuf, F.; Le Potier, M.; Bourry, O. Porcine reproductive and respiratory syndrome virus (PRRSv) modified-live vaccine reduces virus transmission in experimental conditions. Vaccine 2015, 33, 2493–2499. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Lunney, J.K.; Fang, Y.; Ladinig, A.; Chen, N.; Li, Y.; Rowland, B.; Renukaradhya, G.J. Porcine reproductive and respiratory syndrome virus (PRRSV): Pathogenesis and interaction with the immune system. Annu. Rev. Anim. Biosci. 2016, 4, 129–154. [Google Scholar] [CrossRef]
- Kappes, M.A.; Faaberg, K.S. PRRSV structure, replication and recombination: Origin of phenotype and genotype diversity. Virology 2015, 479-480, 475–486. [Google Scholar] [CrossRef]
- Wei, Q.; Fu, G.; Wang, K.; Yang, Q.; Zhao, J.; Wang, Y.; Ji, K.; Song, S. Advances in research on antiviral activities of sulfated polysaccharides from seaweeds. Pharmaceuticals 2022, 15, 581. [Google Scholar] [CrossRef]
- Briseño, J.A.A.; Cruz-Suarez, L.E.; Sassi, J.-F.; Ricque-Marie, D.; Zapata-Benavides, P.; Mendoza-Gamboa, E.; Rodríguez-Padilla, C.; Trejo-Avila, L.M. Sulphated polysaccharides from Ulva clathrata and cladosiphon okamuranus seaweeds both inhibit viral attachment/entry and cell-cell fusion, in NDV infection. Mar. Drugs 2015, 13, 697–712. [Google Scholar] [CrossRef]
- Mendes, G.D.S.; Soares, A.R.; Martins, F.O.; De Albuquerque, M.C.M.; Costa, S.S.; Yoneshigue-Valentin, Y.; Gestinari, L.M.D.S.; Santos, N.; Romanos, M.T.V. Antiviral activity of the green marine alga Ulva fasciata on the replication of human metapneumovirus. Rev. Inst. Med. Trop. Sao Paulo 2010, 52, 3–10. [Google Scholar] [CrossRef]
- Bussy, F.; Rémy, S.; Le Goff, M.; Collén, P.N.; Trapp-Fragnet, L. The sulphated polysaccharides extract ulvans from Ulva armoricana limits Marek’s disease virus dissemination in vitro and promotes viral reactivation in lymphoid cells. BMC Veter Res. 2022, 18, 155. [Google Scholar] [CrossRef]
- Yamada, T.; Ogamo, A.; Saito, T.; Watanabe, J.; Uchiyama, H.; Nakagawa, Y. Preparation and anti-HIV activity of low-molecular-weight carrageenans and their sulfated derivatives. Carbohydr. Polym. 1997, 32, 51–55. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracts | DM% | MM% | OM% | pH | C (%DM) | N (%DM) | S (%DM) |
|---|---|---|---|---|---|---|---|
| Ulva | 13.4% | 50.1% | 49.9% | 2.6 | 18.5% | 1.6% | 9.8% |
| Soleria | 16.1% | 62.7% | 37.3% | 2.6 | 14.4% | 2.9% | 5.3% |
| Genes | Sequences (5′-3′) | Size (bp) | Effic. | Tm (°C) | References |
|---|---|---|---|---|---|
| Cytokines | |||||
| TNFα | F: TGGTGGTGCCGACAGATG R: CAGCCTTGGCCCCTGAA | 64 | 0.96 | 64 | [29] |
| IL1β | F: TGCCAACGTGCAGTCTATGG R: TGGGCCAGCCAGCACTAG | 70 | 0.89 | 60 | NM_214055 |
| IL8/CXCL8 | F: CCGTGTCAACATGACTTCCAA R: GAGCTGCAGAAAGCAGGAAAA | 65 | 0.97 | 60 | NM_213867 |
| IL6 | F: CTGCTTCTGGTGATGGCTACTG R: GGCATCACCTTTGGCATCTT | 69 | 0.94 | 60 | [29] |
| IFNβ | F: TGTGGAACTTGATGGGCAGA R: GAATGGTCATGTCTCCCCTGGT | 92 | 0.98 | 60 | MH538100 |
| IFNλ1 | F: GAGGCTGAGCTAGACTTGAC R: CCTGAAGTTCGACGTGGATG | 115 | 0.95 | 60 | [30] |
| PKR | F: CACATCGGCTTCAGAGTCAG R: GGGCGAGGTAAATGTAGGTG | 166 | 0.96 | 61 | [31] |
| Mx2 | F: CCGACTTCAGTTCAGGATGG R: ACAGGAGACGGTCCGTTTAC | 156 | 1.03 | 62 | [31] |
| References | |||||
| RPS24 | F: AAGGAACGCAAGAACAGAATGAA R: TTTGCCAGCACCAACGTTG | 62 | 0.95 | 60 | [32] |
| RPL19 | F: AACTCCCGTCAGCAGATCC R: AGTACCCTTCCGCTTACCG | 147 | 0.90 | 60 | [30] |
| HPRT1 | F: CTGCACCTCCGCCTCTC R: TCACTAATCACGACGCTGGG | 119 | 0.90 | 60 | XM_021079504 |
| Viruses | |||||
| PRRSV | F: TATGCGAGCTGAATGGGACC R: AGGATATGAGTGGCAACCGG P: HEX-TGGGCAGTTGAGACTTTCGTGCT-TAMRA | / | 60 | [33] | |
| ADV | F: GCGGGTACGTGTACTACGAG R: GAGGCCCTGGAAGAAGTTGG P: 6FAM-ACTACAGCTACGTGCGCATGGTGGAG-TAMRA | 287 | 63 | [34] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hervet, C.; Bussy, F.; Le Goff, C.; Ménard, D.; Collén, P.N.; Goff, M.L.; Meurens, F.; Bertho, N. Marine-Sulfated Polysaccharides Extracts Exhibit Contrasted Time-Dependent Immunomodulatory and Antiviral Properties on Porcine Monocytes and Alveolar Macrophages. Animals 2022, 12, 2576. https://doi.org/10.3390/ani12192576
Hervet C, Bussy F, Le Goff C, Ménard D, Collén PN, Goff ML, Meurens F, Bertho N. Marine-Sulfated Polysaccharides Extracts Exhibit Contrasted Time-Dependent Immunomodulatory and Antiviral Properties on Porcine Monocytes and Alveolar Macrophages. Animals. 2022; 12(19):2576. https://doi.org/10.3390/ani12192576
Chicago/Turabian StyleHervet, Caroline, Frédérick Bussy, Claude Le Goff, Déborah Ménard, Pi Nyvall Collén, Matthieu Le Goff, François Meurens, and Nicolas Bertho. 2022. "Marine-Sulfated Polysaccharides Extracts Exhibit Contrasted Time-Dependent Immunomodulatory and Antiviral Properties on Porcine Monocytes and Alveolar Macrophages" Animals 12, no. 19: 2576. https://doi.org/10.3390/ani12192576