
Does the Domestication Syndrome Apply to the Domestic Pig? Not Completely

Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Prediction 1
1.2. Prediction 2
2. Materials and Methods
2.1. Ethical Statement
2.2. The Study Group
2.3. Data Collection and Video Analyses
2.4. Operational Definitions and Structural Indices
Asymmetry Index (AI)
2.5. Statistical Elaboration
3. Results
3.1. Social Play Levels and Structure in Piglets and Wild Boar Hybrids (Prediction 1)
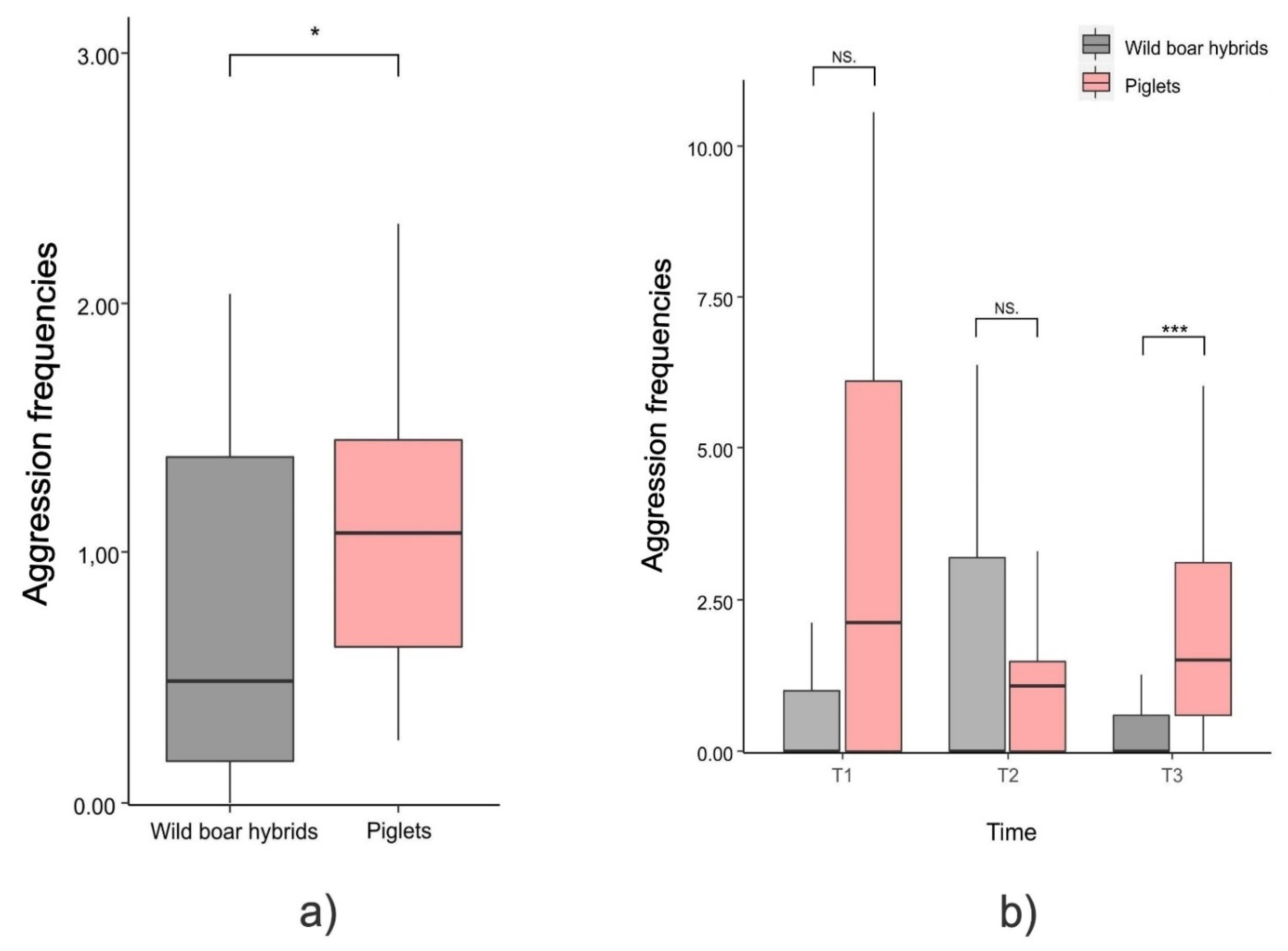
3.2. Aggression Level and Structure in Piglets vs. Wild Boar Hybrids (Prediction 2)
4. Discussion
4.1. Social Play Levels and Structure in Piglets and Wild Boar Hybrids
4.2. Aggression Level and Structure in Piglets vs. Wild Boar Hybrids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mignon-Grasteau, S.; Boissy, A.; Bouix, J.; Faure, J.-M.; Fisher, A.D.; Hinch, G.N.; Jensen, P.; Le Neindre, P.; Mormède, P.; Prunet, P.; et al. Genetics of Adaptation and Domestication in Livestock. Livest. Prod. Sci. 2005, 93, 3–14. [Google Scholar] [CrossRef]
- Price, E.O. Behavioral Aspects of Animal Domestication. Q. Rev. Biol. 1984, 59, 1–32. [Google Scholar] [CrossRef]
- Trut, L.; Oskina, I.; Kharlamova, A. Animal Evolution during Domestication: The Domesticated Fox as a Model. Bioessays 2009, 31, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Clutton-Brock, J. The Process of Domestication. Mamm. Rev. 1992, 22, 79–85. [Google Scholar] [CrossRef]
- Zeder, M.A. The Domestication of Animals. J. Anthropol. Res. 2012, 68, 161–190. [Google Scholar] [CrossRef]
- Zeder, M.A. Core Questions in Domestication Research. Proc. Natl. Acad. Sci. USA 2015, 112, 3191–3198. [Google Scholar] [CrossRef]
- Del Savio, L.; Mameli, M. Human Domestication and the Roles of Human Agency in Human Evolution. Hist. Philos. Life Sci. 2020, 42, 21. [Google Scholar] [CrossRef]
- Murphy, M.R. The Hamster; Siegel, H.I., Ed.; Springer: Boston, MA, USA, 1985. [Google Scholar]
- Tennessen, T.; Hudson, R.J. Traits Relevant to the Domestication of Herbivores. Appl. Anim. Ethol. 1981, 7, 87–102. [Google Scholar] [CrossRef]
- Theofanopoulou, C.; Gastaldon, S.; O’Rourke, T.; Samuels, B.D.; Messner, A.; Martins, P.T.; Delogu, F.; Alamri, S.; Boeckx, C. Self-Domestication in Homo Sapiens: Insights from Comparative Genomics. PLoS ONE 2017, 12, e0185306. [Google Scholar] [CrossRef]
- Trut, L.N. Early Canid Domestication: The Farm-Fox Experiment: Foxes Bred for Tamability in a 40-Year Experiment Exhibit Remarkable Transformations That Suggest an Interplay between Behavioral Genetics and Development. Am. Sci. 1999, 87, 160–169. [Google Scholar] [CrossRef]
- Goodwin, D.; Bradshaw, J.W.S.; Wickens, S.M. Paedomorphosis Affects Agonistic Visual Signals of Domestic Dogs. Anim. Behav. 1997, 53, 297–304. [Google Scholar] [CrossRef]
- Mcnamara, K. Shapes of Time: The Evolution of Growth and Development; Johns Hopkins University Press: Baltimore, MD, USA, 1997. [Google Scholar]
- Coppinger, R.; Glendinning, J.; Torop, E.; Matthay, C.; Sutherland, M.; Smith, C. Degree of Behavioral Neoteny Differentiates Canid Polymorphs. Ethology 2010, 75, 89–108. [Google Scholar] [CrossRef]
- Morey, D.F. The Early Evolution of the Domestic Dog. Am. Sci. 1994, 82, 336–347. [Google Scholar]
- Waller, B.M.; Peirce, K.; Caeiro, C.C.; Scheider, L.; Burrows, A.M.; McCune, S.; Kaminski, J. Paedomorphic Facial Expressions Give Dogs a Selective Advantage. PLoS ONE 2013, 8, e82686. [Google Scholar] [CrossRef]
- Lord, K.A.; Larson, G.; Coppinger, R.P.; Karlsson, E.K. The History of Farm Foxes Undermines the Animal Domestication Syndrome. Trends Ecol. Evol. 2020, 35, 125–136. [Google Scholar] [CrossRef]
- Zipser, B.; Schleking, A.; Kaiser, S.; Sachser, N. Effects of Domestication on Biobehavioural Profiles: A Comparison of Domestic Guinea Pigs and Wild Cavies from Early to Late Adolescence. Front. Zool. 2014, 11, 30. [Google Scholar] [CrossRef] [PubMed]
- Hansen Wheat, C.; Fitzpatrick, J.L.; Rogell, B.; Temrin, H. Behavioural Correlations of the Domestication Syndrome Are Decoupled in Modern Dog Breeds. Nat. Commun. 2019, 10, 2422. [Google Scholar] [CrossRef]
- Hare, B.; Wobber, V.; Wrangham, R. The Self-Domestication Hypothesis: Evolution of Bonobo Psychology Is Due to Selection against Aggression. Anim. Behav. 2012, 83, 573–585. [Google Scholar] [CrossRef]
- Himmler, B.T.; Stryjek, R.; Modlinska, K.; Derksen, S.M.; Pisula, W.; Pellis, S.M. How Domestication Modulates Play Behavior: A Comparative Analysis between Wild Rats and a Laboratory Strain of Rattus Norvegicus. J. Comp. Psychol. 2013, 127, 453–464. [Google Scholar] [CrossRef]
- Künzl, C.; Sachser, N. The Behavioral Endocrinology of Domestication: A Comparison between the Domestic Guinea Pig (Cavia Apereaf. Porcellus) and Its Wild Ancestor, the Cavy (Cavia Aperea). Horm. Behav. 1999, 35, 28–37. [Google Scholar] [CrossRef]
- Trut, L.N. The Evolutionary Concept of Destabilizing Selection: Status Quo In Commemoration of DK Belyaev. J. Anim. Breed. Genet. 1998, 115, 415–431. [Google Scholar] [CrossRef]
- Serpell, J. The Walking Larder; Clutton-Brock, J., Ed.; Routledge: London, UK, 1989. [Google Scholar]
- Zhang, S.-J.; Wang, G.-D.; Ma, P.; Zhang, L.-L.; Yin, T.-T.; Liu, Y.-H.; Otecko, N.O.; Wang, M.; Ma, Y.-P.; Wang, L.; et al. Genomic Regions under Selection in the Feralization of the Dingoes. Nat. Commun. 2020, 11, 671. [Google Scholar] [CrossRef] [PubMed]
- Zohary, D.; Tchernov, E.; Horwitz, L.K. The Role of Unconscious Selection in the Domestication of Sheep and Goats. J. Zool. 1998, 245, 129–135. [Google Scholar] [CrossRef]
- Price, M.; Hongo, H. The Archaeology of Pig Domestication in Eurasia. J. Archaeol. Res. 2020, 28, 557–615. [Google Scholar] [CrossRef]
- Frantz, L.; Meijaard, E.; Gongora, J.; Haile, J.; Groenen, M.A.M.; Larson, G. The Evolution of Suidae. Annu. Rev. Anim. Biosci. 2016, 4, 61–85. [Google Scholar] [CrossRef] [PubMed]
- Larson, G.; Albarella, U.; Dobney, K.; Rowley-Conwy, P.; Schibler, J.; Tresset, A.; Vigne, J.-D.; Edwards, C.J.; Schlumbaum, A.; Dinu, A.; et al. Ancient DNA, Pig Domestication, and the Spread of the Neolithic into Europe. Proc. Natl. Acad. Sci. USA 2007, 104, 15276–15281. [Google Scholar] [CrossRef]
- Price, E.O. Behavioral Development in Animals Undergoing Domestication. Appl. Anim. Behav. Sci. 1999, 65, 245–271. [Google Scholar] [CrossRef]
- D’eath, R.B.; Turner, S.P. The Welfare of Pigs; Springer: Dordrecht, The Netherlands, 2009. [Google Scholar]
- Mendl, M.T. The Social Behaviour of Non-Lactating Cows and Its Implications for Managing Sow Aggression. Pig Vet. J. 1994, 34, 9–20. [Google Scholar]
- Stolba, A.; Wood-Gush, D.G. The Behaviour of Pigs in a Semi-Natural Environment. Anim. Sci. 1989, 48, 419–425. [Google Scholar] [CrossRef]
- Dellmeier, G.R.; Friend, T.H. Behavior and Extensive Management of Domestic Sows (Sus Scrofa) and Litters. Appl. Anim. Behav. Sci. 1991, 29, 327–341. [Google Scholar] [CrossRef]
- Jensen, P. The Weaning Process of Free-Ranging Domestic Pigs: Within-and between-Litter Variations. Ethology 1995, 100, 14–25. [Google Scholar] [CrossRef]
- Petersen, V.; Recén, B.; Vestergaard, K. Behaviour of Sows and Piglets during Farrowing under Free-Range Conditions. Appl. Anim. Behav. Sci. 1990, 26, 169–179. [Google Scholar] [CrossRef]
- Kurz, J.C.; Marchinton, R.L. Radiotelemetry Studies of Feral Hogs in South Carolina. J. Wildl. Manage. 1972, 36, 1240. [Google Scholar] [CrossRef]
- Gustafsson, M.; Jensen, P.; de Jonge, F.H.; Illmann, G.; Spinka, M. Maternal Behaviour of Domestic Sows and Crosses between Domestic Sows and Wild Boar. Appl. Anim. Behav. Sci. 1999, 65, 29–42. [Google Scholar] [CrossRef]
- Gustafsson, M.; Jensen, P.; de Jonge, F.H.; Schuurman, T. Domestication Effects on Foraging Strategies in Pigs (Sus Scrofa). Appl. Anim. Behav. Sci. 1999, 62, 305–317. [Google Scholar] [CrossRef]
- Špinka, M.; Illmann, G.; de Jonge, F.; Andersson, M.; Schuurman, T.; Jensen, P. Dimensions of Maternal Behaviour Characteristics in Domestic and Wild×domestic Crossbred Sows. Appl. Anim. Behav. Sci. 2000, 70, 99–114. [Google Scholar] [CrossRef]
- Cordoni, G.; Gioia, M.; Demuru, E.; Norscia, I. The Dark Side of Play: Play Fighting as a Substitute for Real Fighting in Domestic Pigs, Sus Scrofa. Anim. Behav. 2021, 175, 21–31. [Google Scholar] [CrossRef]
- Lega, C.; Raia, P.; Rook, L.; Fulgione, D. Size Matters: A Comparative Analysis of Pig Domestication. Holocene 2016, 26, 327–332. [Google Scholar] [CrossRef]
- Leno-Colorado, J.; Hudson, N.J.; Reverter, A.; Pérez-Enciso, M. A Pathway-Centered Analysis of Pig Domestication and Breeding in Eurasia. G3 (Bethesda) 2017, 7, 2171–2184. [Google Scholar] [CrossRef]
- Horback, K. Nosing around: Play in Pigs. Anim. Behav. Cogn. 2014, 2, 186. [Google Scholar] [CrossRef]
- Petersen, H.V.; Vestergaard, K.; Jensen, P. Integration of Piglets into Social Groups of Free-Ranging Domestic Pigs. Appl. Anim. Behav. Sci. 1989, 23, 223–236. [Google Scholar] [CrossRef]
- Newberry, R.C.; Wood-Gush, D.G.M. Development of Some Behaviour Patterns in Piglets under Semi-Natural Conditions. Anim. Sci. 1988, 46, 103–109. [Google Scholar] [CrossRef]
- Newberry, R.C.; Wood-Gush, D.G.; Hall, J.W. Playful Behaviour of Piglets. Behav. Processes 1988, 17, 205–216. [Google Scholar] [CrossRef]
- Clouard, C.; Resmond, R.; Prunier, A.; Tallet, C.; Merlot, E. Exploration of Early Social Behaviors and Social Styles in Relation to Individual Characteristics in Suckling Piglets. Sci. Rep. 2022, 12, 2318. [Google Scholar] [CrossRef] [PubMed]
- Burghardt, G.M. The Genesis of Animal Play: Testing the Limits; MIT Press: Cambridge, MA, USA, 2005. [Google Scholar]
- Burghardt, G.M. The Oxford Handbook of the Development of Play; Pellegrini, A.D., Nathan, P., Eds.; Oxford University Press: New York, NY, USA, 2011. [Google Scholar]
- Aldis, O. Play Fighting; Elsevier Science: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Pellis, S.M.; Pellis, V.C. Play Fighting of Rats in Comparative Perspective: A Schema for Neurobehavioral Analyses. Neurosci. Biobehav. Rev. 1998, 23, 87–101. [Google Scholar] [CrossRef]
- Bekoff, M. Social Play Behaviour. Cooperation, Fairness, Trust, and the Evolution of Morality. J. Conscious. Stud. 2001, 8, 81–90. [Google Scholar]
- Newberry, R.C.; Wood-Gush, D.G. The Suckling Behaviour of Domestic Pigs in a Semi-Natural Environment. Behaviour 1985, 95, 11–25. [Google Scholar] [CrossRef]
- Gepts, P.; Papa, R. Encyclopedia of Life Sciences; Nature Publishing Group: London, UK, 2002. [Google Scholar]
- Wilkins, A.S.; Wrangham, R.W.; Fitch, W.T. The “Domestication Syndrome” in Mammals: A Unified Explanation Based on Neural Crest Cell Behavior and Genetics. Genetics 2014, 197, 795–808. [Google Scholar] [CrossRef]
- Wright, D. Article Commentary: The Genetic Architecture of Domestication in Animals. Bioinform. Biol. Insights 2015, 9, 11–20. [Google Scholar] [CrossRef]
- De Boer, S.F.; Buwalda, B.; Koolhaas, J. Stress: Concepts, Cognition, Emotion, and Behavior; Academic Press: Cambridge, MA, USA, 2016; Volume 36. [Google Scholar]
- Norscia, I.; Palagi, E. Cambridge Studies in Biological and Evolutionary Anthropology: The Missing Lemur Link: An Ancestral Step in the Evolution of Human Behaviour Series Number 74: An Ancestral Step in the Evolution of Human Behaviour; Cambridge University Press: Cambridge, UK, 2016. [Google Scholar] [CrossRef]
- Koolhaas, J.M.; Coppens, C.M.; De Boer, S.F.; Buwalda, B.; Meerlo, P.; Timmermans, P.J. The Resident-Intruder Paradigm: A Standardized Test for Aggression, Violence and Social Stress. J. Vis. Exp. 2013, 77, e4367. [Google Scholar] [CrossRef]
- Altmann, J. Observational Study of Behavior: Sampling Methods. Behaviour 1974, 49, 227–266. [Google Scholar] [CrossRef] [PubMed]
- Rauw, W.M. A Note on the Consistency of a Behavioral Play Marker in Piglets. J. Anim. Sci. Biotechnol. 2013, 4, 33. [Google Scholar] [CrossRef] [Green Version]
- Špinka, M. Advances in Pig Welfare; Elsevier Science: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Rushen, J.; Pajor, E. Offence and Defence in Fights between Young Pigs (Sus Scrofa). Aggress. Behav. 1987, 13, 329–346. [Google Scholar] [CrossRef]
- O’Malley, C.I.; Steibel, J.P.; Bates, R.O.; Ernst, C.W.; Siegford, J.M. The Social Life of Pigs: Changes in Affiliative and Agonistic Behaviors Following Mixing. Animals 2022, 12, 206. [Google Scholar] [CrossRef] [PubMed]
- Cordoni, G.; Nicotra, V.; Palagi, E. Unveiling the “Secret” of Play in Dogs (Canis Lupus Familiaris): Asymmetry and Signals. J. Comp. Psychol. 2016, 130, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Cordoni, G.; Norscia, I.; Bobbio, M.; Palagi, E. Differences in Play Can Illuminate Differences in Affiliation: A Comparative Study on Chimpanzees and Gorillas. PLoS ONE 2018, 13, e0193096. [Google Scholar] [CrossRef] [PubMed]
- Türkmen, G.; Kazanci, N. Applications of Various Biodiversity Indices to Benthic Macroinvertebrate Assemblages in Streams of a National Park in Turkey. Rev. Hydrobiol. 2010, 3, 111–125. [Google Scholar]
- Morris, E.K. Choosing and Using Diversity Indices: Insights for Ecological Applications from the German Biodiversity Exploratories. Ecol. Evol. 2014, 4, 3514–3524. [Google Scholar] [CrossRef]
- Supriatna, J. Biodiversity Indexes: Value and evaluation purposes. In E3S Web of Conferences; EDP Sciences: Les Ulis Cedex, France, 2018; Volume 48, p. 01001. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Baltimore, MD, USA, 1949. [Google Scholar]
- Mandaville, S.M. Benthic Macroinvertebrates in Freshwaters: Taxa Tolerance Values, Metrics, and Protocols; Soil & Water Conservation Society of Metro Halifax: Halifax, NS, Canada, 2002; Volume 128. [Google Scholar]
- Keylock, C.J. Simpson Diversity and the Shannon-Wiener Index as Special Cases of a Generalized Entropy. Oikos 2005, 109, 203–207. [Google Scholar] [CrossRef]
- Wilsey, B.J.; Potvin, C. Biodiversity and Ecosystem Functioning: Importance of Species Evenness in an Old Field. Ecology 2000, 81, 887–892. [Google Scholar] [CrossRef]
- Eliazar, I.I.; Sokolov, I.M. Measuring Statistical Evenness: A Panoramic Overview. Phys. A Stat. Mech. Its Appl. 2012, 391, 1323–1353. [Google Scholar] [CrossRef]
- Daly, A.J.; Baetens, J.M.; De Baets, B. Ecological Diversity: Measuring the Unmeasurable. Mathematics 2018, 6, 119. [Google Scholar] [CrossRef]
- Pielou, E.C. Species-Diversity and Pattern-Diversity in the Study of Ecological Succession. J. Theor. Biol. 1966, 10, 370–383. [Google Scholar] [CrossRef]
- Siegel, S.; Castellan, N.J. Nonparametric Statistics for the Behavior Sciences; McGraw-Hill: New York, NY, USA, 1988. [Google Scholar]
- Paquette, D. Fighting and Playfighting in Captive Adolescent Chimpanzees. Aggress. Behav. 1994, 20, 49–65. [Google Scholar] [CrossRef]
- Mckinney, M.L. Piaget, Evolution, and Development; Psychology Press: New York, NY, USA, 1998. [Google Scholar]
- Parker, S.T.; Mckinney, M.L. The Evolution of Cognitive Development in Monkeys, Apes, and Humans; John Hopkins University Press: Baltimore, MD, USA, 1999. [Google Scholar]
- Palagi, E.; Cordoni, G. The Right Time to Happen: Play Developmental Divergence in the Two Pan Species. PLoS ONE 2012, 7, e52767. [Google Scholar] [CrossRef] [PubMed]
- Pellis, S.; Pellis, V. The Playful Brain: Venturing to the Limits of Neuroscience; Oneworld: London, UK, 2009. [Google Scholar]
- Abrantes, R. Evolution of Canine Social Behavior; Dogwise Publishing: Wenatchee, WA, USA, 2004. [Google Scholar]
- Bekoff, M. The Development of Social Interaction, Play, and Metacommunication in Mammals: An Ethological Perspective. Q. Rev. Biol. 1972, 47, 412–434. [Google Scholar] [CrossRef]
- Cordoni, G. Social Play in Captive Wolves (Canis Lupus): Not Only an Immature Affair. Behaviour 2009, 146, 1363–1385. [Google Scholar] [CrossRef]
- Cordoni, G.; Palagi, E. Aggression and Hierarchical Steepness Inhibit Social Play in Adult Wolves. Behaviour 2016, 153, 749–766. [Google Scholar] [CrossRef]
- Smuts, B. The Social Dog; Academic Press: Cambridge, MA, USA, 2014; pp. 105–130. [Google Scholar]
- Hare, B. Survival of the Friendliest: Homo Sapiens Evolved via Selection for Prosociality. Annu. Rev. Psychol. 2017, 68, 155–186. [Google Scholar] [CrossRef]
- Hare, B.; Wrangham, R.; Muller, M.; Wrangham, R.W. Chimpanzees and Human Evolution; Pilbeam, R., Ed.; Harvard University Press: Cambridge, UK, 2017. [Google Scholar]
- Wrangham, R.W. Two Types of Aggression in Human Evolution. Proc. Natl. Acad. Sci. USA 2018, 115, 245–253. [Google Scholar] [CrossRef]
- Pellis, S.M.; Pellis, V.C. What Is Play Fighting and What Is It Good For? Learn. Behav. 2017, 45, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Marshall-Pescini, S.; Schaebs, F.S.; Gaugg, A.; Meinert, A.; Deschner, T.; Range, F. The Role of Oxytocin in the Dog-Owner Relationship. Anim. Open Access J. MDPI 2019, 9, 792. [Google Scholar] [CrossRef] [PubMed]
- Gácsi, M.; Gyõri, B.; Miklósi, Á.; Virányi, Z.; Kubinyi, E.; Topál, J. Species-Specific Differences and Similarities in the Behavior of Hand-Raised Dog and Wolf Pups in Social Situations with Humans. Dev. Psychobiol 2005, 47, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Frank, H.; Frank, M.G. On the Effects of Domestication on Canine Social Development and Behavior. Appl. Anim. Ethol 1982, 8, 507–525. [Google Scholar] [CrossRef]
- Scott, J.P.; Fuller, J.L. Genetics and the Social Behaviou.r of the Dog; University of Chicago Press: Chicago, IL, USA, 1965. [Google Scholar]
- Lazzaroni, M.; Range, F.; Backes, J.; Portele, K.; Scheck, K.; Marshall-Pescini, S. The Effect of Domestication and Experience on the Social Interaction of Dogs and Wolves with a Human Companion. Front. Psychol. 2020, 11, 785. [Google Scholar] [CrossRef] [PubMed]
- Range, F.; Marshall-Pescini, S.; Kratz, C.; Virányi, Z. Wolves Lead and Dogs Follow, but They Both Cooperate with Humans. Sci. Rep. 2019, 9, 3796. [Google Scholar] [CrossRef] [PubMed]
- Cafazzo, S.; Marshall-Pescini, S.; Lazzaroni, M.; Virányi, Z.; Range, F. The Effect of Domestication on Post-Conflict Management: Wolves Reconcile While Dogs Avoid Each Other. R. Soc. Open Sci. 2018, 5, 171553. [Google Scholar] [CrossRef]
- Cordoni, G.; Palagi, E. Reconciliation in Wolves (Canis Lupus): New Evidence for a Comparative Perspective. Ethology 2008, 114, 298–308. [Google Scholar] [CrossRef]
- Gabor, T.M.; Hellgren, E.C.; Van Den Bussche, R.A.; Silvy, N.J. Demography, Sociospatial Behaviour and Genetics of Feral Pigs (Sus Scrofa) in a Semi-Arid Environment. J. Zool. 1999, 247, 311–322. [Google Scholar] [CrossRef]
- Kaminski, G.; Brandt, S.; Baubet, E.; Baudoin, C. Life-History Patterns in Female Wild Boars (Sus Scrofa): Mother-Daughter Postweaning Associations. Can. J. Zool. 2005, 83, 474–480. [Google Scholar] [CrossRef]
- Appleby, M.C. The Probability of Linearity in Hierarchies. Anim. Behav. 1983, 31, 600–608. [Google Scholar] [CrossRef]
- De Vries, H. An Improved Test of Linearity in Dominance Hierarchies Containing Unknown or Tied Relationships. Anim. Behav. 1995, 50, 1375–1389. [Google Scholar] [CrossRef]
- Drews, C. The Concept and Definition of Dominance in Animal Behaviour. Behaviour 1993, 125, 283–313. [Google Scholar] [CrossRef]
- Kendall, M.G. Rank Correlation Methods; Griffin: Oxford, UK, 1948. [Google Scholar]
- Landau, H.G. On Dominance Relations and the Structure of Animal Societies: I. Effect of Inherent Characteristics. Bull. Math. Biophys. 1951, 13, 1–19. [Google Scholar] [CrossRef]
- Andersen, L.B.; Hasselstrøm, H.; Grønfeldt, V.; Hansen, S.E.; Karsten, F. The Relationship between Physical Fitness and Clustered Risk, and Tracking of Clustered Risk from Adolescence to Young Adulthood: Eight Years Follow-up in the Danish Youth and Sport Study. Int. J. Behav. Nutr. Phys. Act. 2004, 1, 6. [Google Scholar] [CrossRef] [PubMed]
- Clutton-Brock, T.H. The Functions of Antlers. Behaviour 1982, 79, 108–124. [Google Scholar] [CrossRef]
- Krebs, J.R.; Davies, N.B. An Introduction to Behavioural Ecology, 4th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Norring, M.; Valros, A.; Bergman, P.; Marchant-Forde, J.N.; Heinonen, M. Body Condition, Live Weight and Success in Agonistic Encounters in Mixed Parity Groups of Sows during Gestation. Animal 2017, 13, 392–398. [Google Scholar] [CrossRef] [PubMed]
- Preuschoft, S.; Van Schaik, C.P. Dominance and Communication; Nature Conflict Resolution: Berkeley, CA, USA, 2000; pp. 77–105. [Google Scholar]
- Sapolsky, R.M. The Influence of Social Hierarchy on Primate Health. Science 2005, 308, 648–652. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Litter | Individuals | Date of Birth |
|---|---|---|---|
| Piglets | 1 | 11 (5 female; 6 male) | 16 Sept 2018 |
| 2 | 6 (2 female; 4 male) | 3 Oct 2018 | |
| 3 | 8 (4 female; 4 male) | 5 Nov 2018 | |
| Wild boar hybrids | 1 | 9 (5 female; 4 male) | 14 Jun 2018 |
| 2 | 8 (5 female; 3 male) | 05 Jun 2018 | |
| 3 | 10 (3 female; 7 male) | 10 Jun 2018 |
| Play Patterns | |||
|---|---|---|---|
| Category | Behavioural Pattern | Type | Description |
| Offensive | Attempt play bite | C | A piglet attempts to bite the partner, but there is no contact with it |
| Head play knocking | C | A piglet hits another individual with the head | |
| Play bite | C | A piglets bites a partner by delicately closing mouth over the other’s flesh | |
| Play lifting | C | A piglet attempts to displace a partner by lifting or levering it with snout or head | |
| Play mount/climb | C | A piglet places both front hoofs on the back of another piglet or sow | |
| Play push | C | A piglet drives its head, neck, or shoulders with minimal or moderate force into another piglet’s body. Occasionally, this pattern results in the displacement of the target animal. It is significantly more intensive than nudging | |
| Play run | LA | A piglet runs and hops in forward motions within the pen environment. Run can be performed both in solitary and social manner | |
| Neutral | Flopping | LA | A piglet drops to the pen floor from a normal upright position to a sitting or lying position. There is no contact with an object or another individual that could cause the change in position |
| Head tossing | LA | A piglet gently head shakes from one side to another | |
| Hopping | LA | A piglet has either its two front feet or all four feet off the pen floor at one time through an energetic upwards jumping movement. The piglet continues facing the same original direction for the whole of the behaviour | |
| Leg spreading | LA | A piglet spreads its fore and hind limbs and it moves quickly from side to side | |
| Nudge | C | A piglet uses its snout to gently touch another piglet’s body (excluding nose–nose contact). It is more intensive than touching, but also more gentle than pushing | |
| Object play | LA | A piglet manipulates an item or securely holds it in its mouth, energetically shaking it or carrying it around the pen | |
| Pivot | LA | A piglet twirls its body on the horizontal plane by a minimum of 90°. Pivot is usually associated with jumping on the spot | |
| Scamper | LA | A piglet performs two or more forward directed hops in quick succession of each other usually associated with excitability | |
| Defensive | Play kneeling | LA | A piglet goes down on its knees while playing |
| Play lying down | LA | A piglet places itself in a horizontal position during play | |
| Play sitting | LA | A piglet sits during play | |
| Aggressive Patterns | |||
|---|---|---|---|
| Category | Behavioural Pattern | Type | Description |
| Offensive | Aggressive bite | C | A piglet opens its mouth and closes its teeth tight on a small piece of the opponent’s flesh (except tail) |
| Aggressive head knocking | C | A piglet lunges or jerks its head with physical contact and mouth closed | |
| Aggressive kick | C | A piglet kicks with one or both hind limbs the opponent, striking it | |
| Aggressive lifting | C | A piglet attempts to displace the opponent by lifting or levering it with snout or head | |
| Aggressive mount/climb | C | A piglet forces the opponent to move away by rising upon the rear of the partner | |
| Aggressive push | C | A piglet presses its head, neck, shoulder or body against the opponent in an aggressive context | |
| Attempt aggressive bite | C | A piglet opens its mouth, directs or turns its head towards the body of the opponent and closes its mouth without contact | |
| Neutral | Head tilting | LA | A piglet moves the head to the side when the opponent passes or gets closer |
| Threat | LA | A piglet arches the back to the opponent or makes a forward movement of the head and stares at the opponent with no physical contact | |
| Rest during fight | LA | A piglet rests and does not exhibit aggressive behavior while being hit-during a reciprocal real fight session for at least 3 s. (reciprocal fighting must occur before and after this event for it to be classified as a rest during fight). | |
| Defensive | Asymmetric parallel | C | The piglets involved in a real fight face the same direction, standing side by side and one of them is slightly ahead of the other. A piglet—the one placed slightly in front of the other—moves forward, pushing the opponent away with his shoulder and moving his head away from the opponent to avoid having its ears bitten. |
| Avoidance | LA | A piglet moves away with a depressed tail when the opponent approaches | |
| Flee | LA | A piglet runs away from the opponent. The opponent can react with a chase. | |
| Withdrawal | LA | A piglet tries to leave a reciprocal real fight session, the opponent continues to bite the recipient with a rate greater than one bite for 3 s, and the recipient reacts with any harmful aggression for more than 3 s. After that, the piglets involved do not have interactions for at least 3 s. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Collarini, E.; Gioia, M.; Cordoni, G.; Norscia, I. Does the Domestication Syndrome Apply to the Domestic Pig? Not Completely. Animals 2022, 12, 2458. https://doi.org/10.3390/ani12182458
Collarini E, Gioia M, Cordoni G, Norscia I. Does the Domestication Syndrome Apply to the Domestic Pig? Not Completely. Animals. 2022; 12(18):2458. https://doi.org/10.3390/ani12182458
Chicago/Turabian StyleCollarini, Edoardo, Marika Gioia, Giada Cordoni, and Ivan Norscia. 2022. "Does the Domestication Syndrome Apply to the Domestic Pig? Not Completely" Animals 12, no. 18: 2458. https://doi.org/10.3390/ani12182458