
Influence of Water Salinity on the Growth and Survivability of Asp Larvae Leuciscus aspius (Linnaeus, 1758) under Controlled Conditions

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
- TL—mean total length of a specimen (longitudo totalis),
- n1—at the beginning of the time period,
- n2—at the end of the time period,
- t—duration of the rearing period (days—d).
- W1—mean initial weight of a reared individual (mg),
- W2—mean final weight of a reared individual (mg),
- n1—number of individuals (indiv.) at the onset of rearing,
- n2—number of individuals (indiv.) at the end of rearing,
- t—duration of the rearing-up period (days).
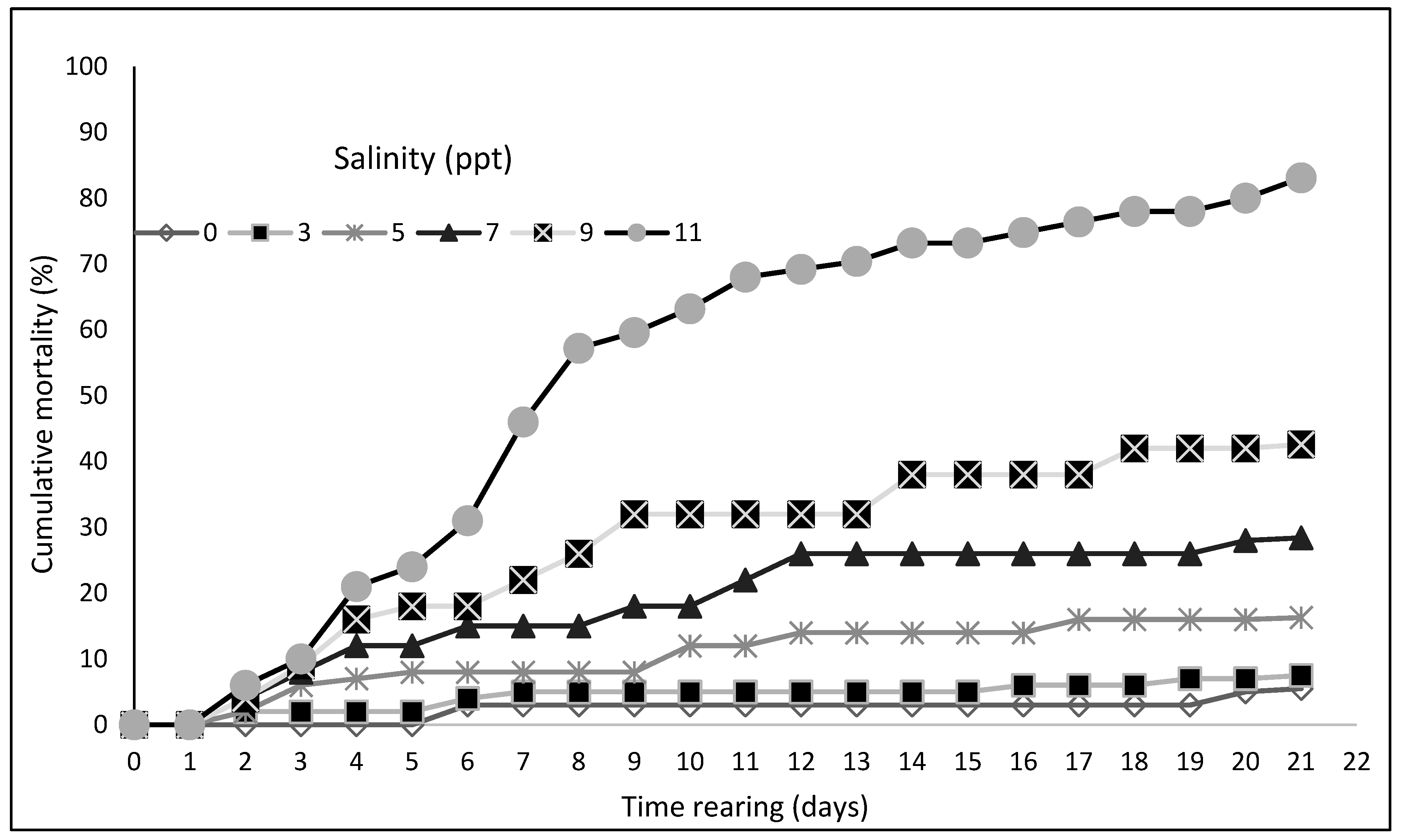
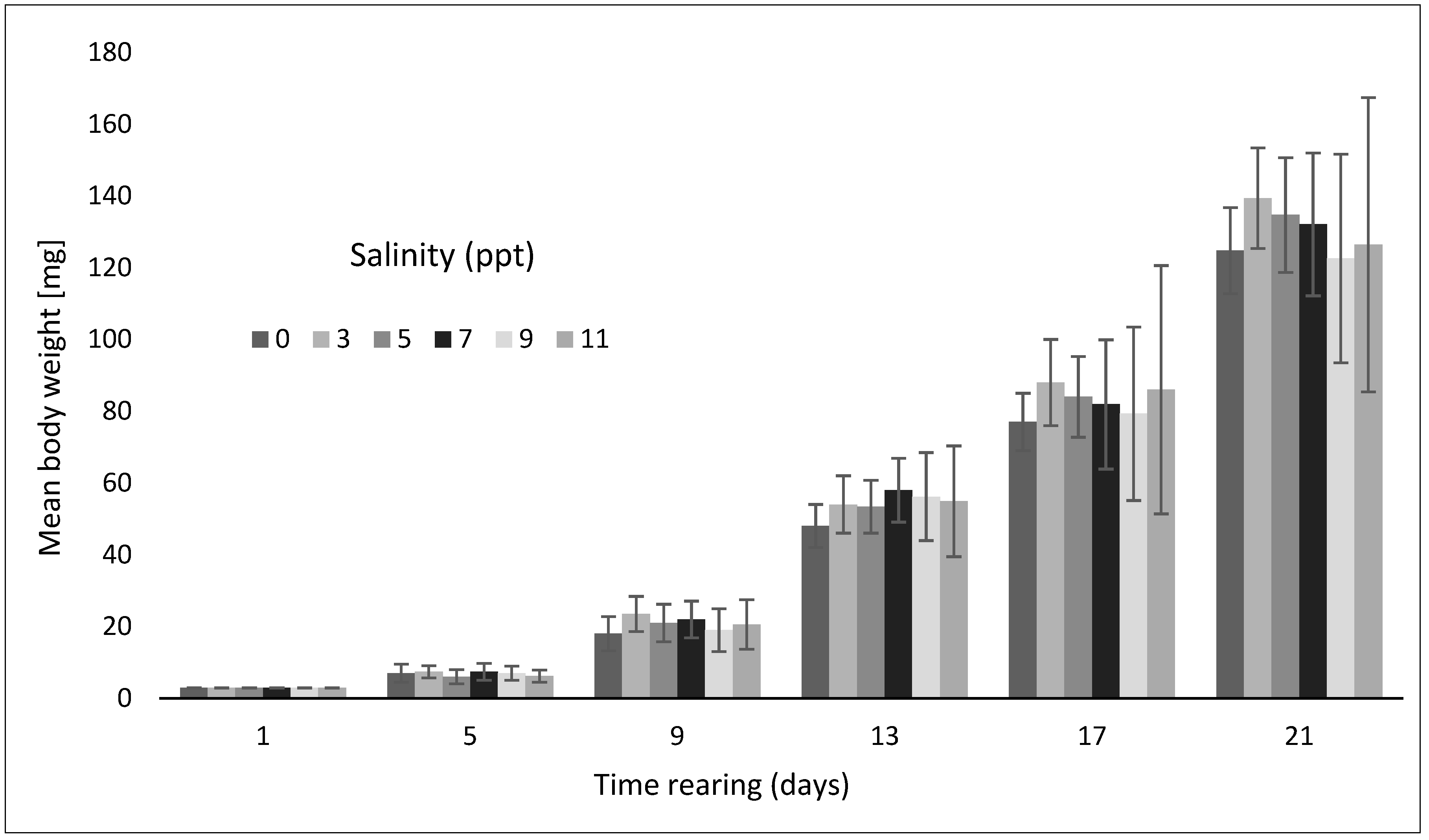
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brown, M.E. Experimental studies on growth. In The Physiology of Fishes; Brown, M.E., Ed.; Academic Press: New York, NY, USA, 1957; pp. 361–400. [Google Scholar]
- Bœuf, G.; Payan, P. How should salinity influence Fish growth? Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 130, 411–423. [Google Scholar] [CrossRef]
- James, K.R.; Cant, B.; Ryan, T. Responses of freshwater biota to rising salinity levels and implications for saline water management: A review. Aust. J. Bot. 2003, 51, 703–713. [Google Scholar] [CrossRef]
- Fridman, S.; Bron, J.; Rana, K. Influence of salinity on embryogenesis, survival, growth and oxygen consumption in embryos and yolk-sac larvae of the Nile tilapia. Aquaculture 2012, 334–337, 182–190. [Google Scholar] [CrossRef]
- Mardaneh Khatooni, M.; Mojazi Amiri, B.; Mirvaghefi, A.; Jafari, V.; Hoseinifar, S.H. The effects of salinity on the fertilization rate and rearing of the Persian sturgeon (Acipenser persicus) larvae. Aquac. Int. 2012, 20, 1097–1105. [Google Scholar] [CrossRef]
- Kültz, D. Physiological mechanisms used by fish to cope with salinity stress. J. Exp. Biol. 2015, 218, 1907–1914. [Google Scholar] [CrossRef] [PubMed]
- Kujawa, R.; Lach, M.; Pol, P.; Ptaszkowski, M.; Mamcarz, A.; Nowosad, J.; Furgała-Selezniow, G.; Kucharczyk, D. Influence of water salinity on the survival of embryos and growth of the sichel larvae Pelecus cultratus (L.) under controlled conditions. Aquac. Res. 2017, 48, 1302–1314. [Google Scholar] [CrossRef]
- Jackson, A.G. Salinity tolerance and osmotic behavior of European carp (Cyprinus carpio L.) from the river Murray. Aust. Trans. R. Soc. Aus. 1981, 103, 185–189. [Google Scholar]
- Wang, J.Q.; Lui, H.; Po, H.; Fan, L. Influence of salinity on food consumption, growth and energy conversion efficiency of common carp (Cyprinus carpio) fingerlings. Aquaculture 1997, 148, 115–124. [Google Scholar] [CrossRef]
- Al-Faiz, A.F.; Jabir, A.; Yesser, A. Variance salinity concentrations effects on survival, growth and feeding of common carp (Cyprinus carpio) fingerlings. Iraqi J. Aquac. 2009, 6, 59–70. [Google Scholar] [CrossRef]
- Kang’ombe, J.; Brown, J.A. Effect of salinity on growth, feed utilization, and survival of Tilapia rendalli under laboratory conditions. J. Appl. Aquac. 2010, 20, 256–271. [Google Scholar] [CrossRef]
- Al-Khshali, M.S.; Al Hilali, H.A. Influence of transfer to high salinity on chloride cells, oxygen and energy consumption in Common carp Cyprinus carpio. J. Anim. Sci. Prod. 2019, 2, 1–12. [Google Scholar]
- Leuciscus aspius (Linnaeus, 1758). Available online: https://www.fishbase.se/summary/Leuciscus-aspius.html (accessed on 5 August 2022).
- Targońska, K.; Kupren, K.; Żarski, D.; Król, R.; Kucharczyk, D. Influence of thermal conditions on successful ide (Leuciscus idus L.) artificial reproduction during spawning season. Ital. J. Anim. Sci. 2011, 10, e50. [Google Scholar] [CrossRef] [Green Version]
- Kujawa, R.; Mamcarz, A.; Kucharczyk, D. Effect of temperature on embryonic development of asp [Aspius aspius L.]. Pol. Arch. Hydrobiol. 1997, 44, 1–2. [Google Scholar]
- Babiak, I.; Glogowski, J.; Kujawa, R.; Kucharczyk, D.; Mamcarz, A. Cryopreservation of sperm from asp Aspius aspius. Progress. Fish-Cult. 1998, 60, 146–148. [Google Scholar] [CrossRef]
- Kupren, K.; Turkowski, K.; Kucharczyk, D.; Krejszeff, S.; Żarski, D.; Hakuć-Błażowska, A.; Targońska, K.; Kwiatkowski, M.; Jamróz, M.; Czarkowski, T. Economic aspects of rearing larval asp, Aspius aspius (L.), and ide, Leuciscus idus (L.), in closed recirculating systems. Fish. Aquat. Life 2008, 16, 413–420. [Google Scholar] [CrossRef]
- Kwiatkowski, M.; Żarski, D.; Kucharczyk, D.; Kupren, K.; Jamróz, M.; Targońska, K.; Krejszeff, S.; Hakuć-Błażowska, A.; Kujawa, R.; Mamcarz, A. Influence of feeding natural and formulated diets on chosen rheophilic cyprinid larvae. Fish. Aquat. Life 2008, 16, 383–396. [Google Scholar] [CrossRef]
- Hakuć-Błażowska, A.; Kupren, K.; Turkowski, K.; Targońska, K.; Jamróz, M.; Krejszeff, S.; Kwiatkowski, M.; Żarski, D.; Kucharczyk, D. Comparison of economic effectiveness of applying different hormonal preparations for reophile cyprinid fish reproduction stimulation based on the example of asp Aspius aspius (L.) and ide Leuciscus idus (L.). Pol. J. Nat. Sci. 2009, 24, 224–234. [Google Scholar] [CrossRef]
- Kucharczyk, D.; Nowosad, J.; Łuczyński, M.J.; Targońska, K. New technique for fertilizing eggs of burbot, asp and ide under hatchery conditions. Anim. Reprod. Sci. 2016, 172, 143–147. [Google Scholar] [CrossRef]
- Kopiejewska, W. Asp Aspius aspius (L.). In Freshwater Fish of Poland; Brylińska, M., Ed.; Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1986; pp. 220–223. (In Polish) [Google Scholar]
- Pennanen, J.T.; Kaukoranta, M. Asp, a Promising Predator for Stocking Eutrophic Waters; International Symposium on Mass Removal of Unwanted Fish in large Inland Waters: Lahti, Finland, 1991; pp. 1–5. [Google Scholar]
- Donabaum, K.; Schagerl, M.; Dokulil, M.T. Integrated management to restore macrophyte domination. Hydrobiologia 1999, 395/396, 87–97. [Google Scholar] [CrossRef]
- Lyach, R.; Čech, M. A new trend in Central European recreational fishing: More fishing visits but lower yield and catch. Fish. Res. 2018, 201, 131–137. [Google Scholar] [CrossRef]
- Kujawa, R.; Kucharczyk, D.; Mamcarz, A.; Jamróz, M.; Kwiatkowski, M.; Targońska, K.; Żarski, D. Impact of supplementing natural feed with dry diets on the growth and survival of larval asp, Aspius aspius (L.), and nase, Chondrostoma nasus (L.). Fish. Aquat. Life 2010, 18, 13–23. [Google Scholar] [CrossRef]
- Kujawa, R.; Pol, P.; Mamcarz, A.; Kucharczyk, D.; Furgała-Selezniow, G. Larval development sichel (Pelecus cultratus) under controlled conditions. In Hatchery Aquatic Organisms—Achievements, Challenges and Prospects; Zakęś, Z., Demska-Zakęś, K., Kowalska, A., Eds.; IRS: Olsztyn, Poland, 2012; pp. 149–154. (In Polish) [Google Scholar]
- Kujawa, R.; Furgała-Selezniow, G.; Mamcarz, A.; Lach, M.; Kucharczyk, D. Influence of temperature on the growth and survivability of sichel larvae Pelecus cultratus reared under controlled conditions. Ichthyol. Res. 2015, 62, 163–170. [Google Scholar] [CrossRef]
- Kupren, K.; Żarski, D.; Krejszeff, S.; Kucharczyk, D.; Targońska, K. Effect of stocking density on growth, survival and development of asp Aspius aspius (L.), ide Leuciscus idus (L.) and chub Leuciscus cephalus (L.) larvae during initial rearing under laboratory conditions. Ital. J. Anim. Sci. 2011, 10, e34. [Google Scholar] [CrossRef]
- Żarski, D.; Kucharczyk, D.; Kwiatkowski, M.; Targońska, K.; Kupren, K.; Krejszeff, S.; Jamróz, M.; Hakuć-Błażowska, A.; Kujawa, R.; Mamcarz, A. The effect of stocking density on the growth and survival of larval asp, Aspius aspius (L.), and European chub, Leuciscus cephalus (L.), during rearing under controlled conditions. Fish. Aquat. Life 2008, 16, 371–381. [Google Scholar] [CrossRef]
- Engstrom-Ost, J.; Lehtiniemi, M.; Jonasdottir, S.H.; Viitasalo, M. Growth of pike larvae (Esox lucius) under different conditions of food quality and salinity. Ecol. Freshw. Fish 2005, 14, 385–393. [Google Scholar] [CrossRef]
- Santos, J.C.E.; Luz, R.K. Effect of salinity and prey concentrations on Pseudoplatystoma corruscans, Prochilodus costatus and Lophiosilurus alexandri larviculture. Aquaculture 2009, 287, 324–328. [Google Scholar] [CrossRef]
- Jomori, R.K.; Luz, R.K.; Portella, M.C. Effect of Salinity on Larval Rearing of Pacu, Piaractus mesopotamicus, a Freshwater Species. J. World Aquac. Soc. 2012, 43, 423–432. [Google Scholar] [CrossRef]
- Jun, Q.; Pao, X.; Haizhen, W.; Ruiwei, L.; Hui, W. Combined effect of temperature, salinity and density on the growth and feed utilization of Nile tilapia juveniles (Oreochromis niloticus). Aquac. Res. 2012, 43, 1344–1356. [Google Scholar] [CrossRef]
- Dulski, T.; Kujawa, R.; Godzieba, M.; Ciesielski, S. Effect of salinity on the gut microbiome of pike fry (Esox lucius). Appl. Sci. 2020, 10, 2506. [Google Scholar] [CrossRef]
- Sorgeloos, P.; Bossuyt, E.; Lavina, E.; Baeza-Mesa, M.; Persoone, G. Decapsulation of Artemia cysts: A simple technique for the improvement of the use of brine shrimp in aquaculture. Aquaculture 1977, 12, 311–315. [Google Scholar] [CrossRef]
- Nowosad, J.; Żarski, D.; Biłas, M.; Dryl, K.; Krejszeff, S.; Kucharczyk, D. Dynamics of ammonia excretion in juvenile common tench, Tinca tinca (L.), during intensive rearing under controlled conditions. Aquac. Int. 2013, 21, 629–637. [Google Scholar] [CrossRef]
- Sikora, M.; Nowosad, J.; Kucharczyk, D. Nitrogen compound oxidation rate in recirculation systems using three biological filter medias in rearing common carp (Cyprinus carpio L.) juveniles. Aquaculture 2022, 547, 737532. [Google Scholar] [CrossRef]
- Imanpoor, M.R.; Najafi, E.; Kabir, M. Effects of different salinity and temperatures on the growth, survival, haematocrit and blood biochemistry of Goldfish (Carassius auratus). Aquac. Res. 2012, 43, 332–338. [Google Scholar] [CrossRef]
- Peňáz, M.; Prokeš, M.; Kouřil, J.; Hamáčková, J. Influence of water temperature on the early development and growth of the tench, Tinca tinca. Folia Zool. 1989, 38, 275–287. [Google Scholar]
- Myszkowski, L. Pitfalls of using growth rate coefficients. Pol. Arch. Hydrobiol. 1997, 44, 3. [Google Scholar]
- Duncan, D.B. Multiple range and multiple F-test. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Febry, R.; Lutz, P. Energy partitioning in fish: The activity related cost of osmoregulation in a euryhaline cichlid. J. Exp. Biol. 1987, 128, 63–85. [Google Scholar] [CrossRef]
- Varsamos, S.; Nebel, C.; Charmantier, G. Ontogeny of osmoregulation in postembryonic fish: A review. Comp. Biochem. Physiology. Part A Mol. Integr. Physiol. 2005, 141, 401–429. [Google Scholar] [CrossRef]
- Alderdice, D.F. Osmotic and ionic regulation in teleost eggs and larvae. In Fish Physiology; Hoar, W.S., Randall, D.J., Eds.; Academic Press: New York, NY, USA, 1988; Volume XIA, pp. 163–251. [Google Scholar]
- Watanabe, W.O.; Kuo, C.M.; Huang, M.C. Salinity tolerance of Nile tilapia fry (Oreochromis niloticus) spawned and hatched at various salinities. Aquaculture 1985, 48, 432–442. [Google Scholar] [CrossRef]
- Suresh, A.V.; Lin, C.K. Tilapia culture in saline water: A review. Aquaculture 1992, 106, 201–226. [Google Scholar] [CrossRef]
- El-Sayed, A.M. Tilapia culture in salt water: Environmental requirements, nutritional implications and economic potentials. In Proceedings of the 8th Symposium on Advances in Nutritional Aquaculture, Nuevo Leon, Mexico, 15–17 November 2006; pp. 15–17. [Google Scholar]
- Bein, R.; Ribi, G. Effects of larval density and salinity on the development of perch larvae (Perca fluviatilis L.). Aquat. Sci. 1994, 56, 97–105. [Google Scholar] [CrossRef]
- Jäger, T.; Nellen, W.; Schöfer, W.; Shodjai, F. Influence of salinity and temperature on early life stages of Coregonus albula, C. lavaretus, R. rutilus and L. lota. Rapp. Proces-Verbaux Réun. Cons. Perm. Int. Pour L’Explor. Mer 1981, 178, 345–348. [Google Scholar]
- Klinkhardt, M.B.; Winkler, H.M. Einfluß der Salinität auf die Befruchtungs- und Entwicklungsfähigkeit der Eier von vier Süßwasserfischarten Plötz (Rutilus rutilus), Barsch (Perca fluviatilis), Kaulbarsch (Gymnocephalus cernua) und Zander (Stizostedion lucioperca). Wissensch. Zeitschr. Univ. Rostock 1989, 38, 23–30. [Google Scholar]
- Grøtan, K.; Østbye, K.; Taugbøl, A.; Vøllestad, L.A. No shortterm effect of salinity on oxygen consumption in threespine stickleback (Gasterosteus aculeatus) from fresh, brackish, and salt water. Can. J. Zool. 2012, 90, 1386–1393. [Google Scholar] [CrossRef]
- Morgan, J.D.; Iwama, G.K. Effects of salinity on growth, metabolism, and ion regulation in juvenile rainbow trout (Oncorhynchus mykiss) and fall Chinook salmon (Oncorhynchus tshawytscha). Can. J. Fish. Aquacult. Sci. 1991, 48, 2083–2094. [Google Scholar] [CrossRef]
- Swanson, C. Interactive effects of salinity on metabolic rate, activity, growth and osmoregulation in the euryhaline milkfish (Chanos chanos). J. Exp. Biol. 1998, 201, 3355–3366. [Google Scholar] [CrossRef] [PubMed]
- Altinok, I.; Grizzle, J.M. Effects of brackish water on growth, feed conversion and energy absorption efficiency by juvenile euryhaline and freshwater stenohaline fishes. J. Fish Biol. 2001, 59, 1142–1152. [Google Scholar] [CrossRef]
- Holliday, F.G.T. The effects of salinity on the eggs and larvae of teleosts. In Fish Physiology; Hoar, W.S., Randall, D.J., Eds.; Academic Press: New York, NY, USA; London, UK, 1969; Volume IV, pp. 293–310. [Google Scholar]
- Bengtson, D.A.; Léger, P.; Sorgeloos, P. Use of Artemia as a food source for aquaculture. In Artemia Biology; CRC Press: Boca Raton, FL, USA, 2008; pp. 255–286. [Google Scholar]
- Sserwadda, M.; Kagambe, E.; Van Stappen, G. The Brine Shrimp Artemia Survives in Diluted Water of Lake Bunyampaka, an Inland Saline Lake in Uganda. Water 2018, 10, 189. [Google Scholar] [CrossRef]
- Soundarapandian, P.; Saravanakumar, G. Effect of Different Salinities on the Survival and Growth of Artemina spp. Curr. Res. J. Biol. Sci. 2009, 1, 20–22. [Google Scholar]
- Dhont, J.; Sorgeloos, P. Applications of Artemia. In Artemia: Basic and Applied Biology; Springer: Dordrecht, The Netherlands, 2002; Volume 1, pp. 251–277. [Google Scholar]
- Cañedo-Argüelles, M.; Kefford, B.J.; Piscart, C.; Prat, N.; Schäfer, R.B.; Schulz, C.J. Salinisation of rivers: An urgent ecological issue. Environ. Pollut. 2013, 173, 157–167. [Google Scholar] [CrossRef]
- Molenda, T. Impact of a Saline Mine Water Discharge on the Development of a Meromictic Pond, the Rontok Wielki Reservoir, Poland. Mine Water Environ. 2018, 37, 807–814. [Google Scholar] [CrossRef]
- Skovrind, M.; Olsen, M.T.; Vieira, F.G.; Pacheco, G.; Carl, H.; Gilbert, M.T.P.; Møller, P.R.R. Genomic population structure of freshwater-resident and anadromous ide (Leuciscus idus) in north-western Europe. Ecol. Evol. 2016, 6, 1064–1074. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Salinity (ppt) | ||||||
|---|---|---|---|---|---|---|
| 0 | 3 | 5 | 7 | 9 | 11 | |
| Survival rate (%) | 94.5 ± 3.71 a | 92.5 ± 4.24 a | 83.7 ± 5.12 b | 71.6 ± 3.54 c | 57.4 ± 4.39 d | 16.9 ± 3.98 e |
| ITL (mm·d−1) | 0.8 ± (0.02) a | 0.9 ± 0,02 b | 0.81 ± 0.02 c | 0.8 ± 0.02 a | 0.8 ± 0.03 a | 0.8 ± 0.04 a |
| PM (mg) | 121.9 ± 12.24 a | 136.5 ± 14.06 a | 131.8 ± 16.37 a | 129.2 ± 19.91 a | 119.7 ± 29.08 a | 123.5 ± 41.73 a |
| RGR for weight (%/d) | 19.62 ± 2.11 a | 20.25 ± 1.85 a | 20.06 ± 1.76 a | 19.94 ± 2.07 a | 19.52 ± 2.67 a | 19.69 ± 2.94 a |
| RGR for length (%/d) | 5.56 ± 0.43 a | 5.79 ± 0.52 b | 5.66 ± 0.27 c | 5.50 ± 0.38 a | 5.46 ± 0.41 a | 5.68 ± 0.43 a |
| RBR for biomass (RBR) (%/d) | 19.30 ± 2.43 a | 19.81 ± 1.03 a | 19.04 ± 1.14 a | 18.05 ± 0.42 ab | 16.40 ± 1.03 c | 9.99 ± 3.11 d |
| Biomass (g) | 117.94 a | 128.95 a | 112.74 a | 94.58 ab | 70.37 c | 21.36 d |
| Biomass (g·dm−3) | 4.7 ± 0.35 a | 5.2 ± 0.48 a | 4.5 ± 0.45 a | 3.8 ± 0.39 ab | 2.8 ± 0.28 c | 0.9 ± 0.26 d |
| Salinity (ppt) | RFL (%) | RFW (%) | RFB (%) |
|---|---|---|---|
| 3 | 105.35 | 111.70 | 109.33 |
| 5 | 102.06 | 107.93 | 95.60 |
| 7 | 99.59 | 105.85 | 80.20 |
| 9 | 97.94 | 98.24 | 59.67 |
| 11 | 102.47 | 101.28 | 18.11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kujawa, R.; Piech, P. Influence of Water Salinity on the Growth and Survivability of Asp Larvae Leuciscus aspius (Linnaeus, 1758) under Controlled Conditions. Animals 2022, 12, 2299. https://doi.org/10.3390/ani12172299
Kujawa R, Piech P. Influence of Water Salinity on the Growth and Survivability of Asp Larvae Leuciscus aspius (Linnaeus, 1758) under Controlled Conditions. Animals. 2022; 12(17):2299. https://doi.org/10.3390/ani12172299
Chicago/Turabian StyleKujawa, Roman, and Przemysław Piech. 2022. "Influence of Water Salinity on the Growth and Survivability of Asp Larvae Leuciscus aspius (Linnaeus, 1758) under Controlled Conditions" Animals 12, no. 17: 2299. https://doi.org/10.3390/ani12172299