

Ruminal Bacterial Community Successions in Response to Monensin Supplementation in Goats

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design, Sample Preparation and 16S rRNA Sequencing
2.2. Statistical Analysis
3. Results
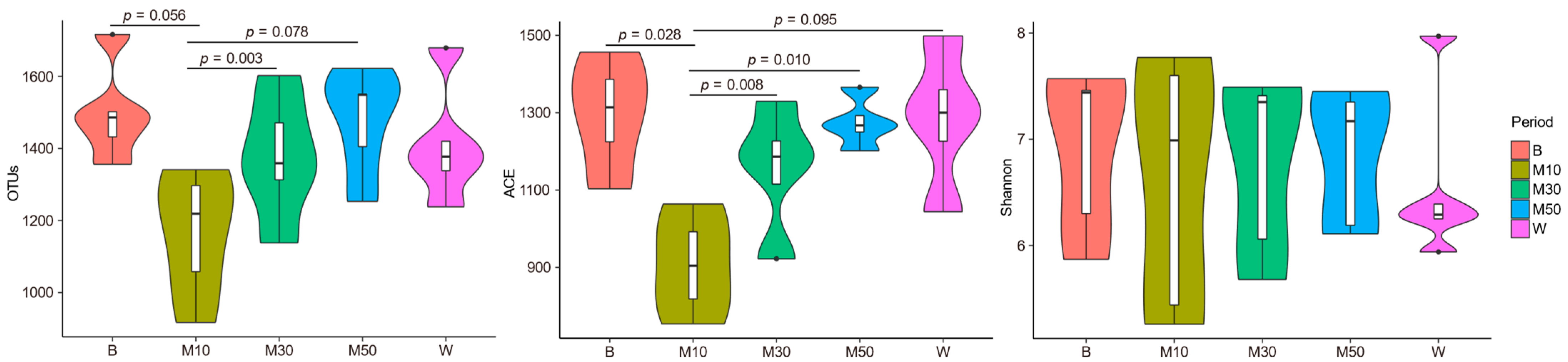
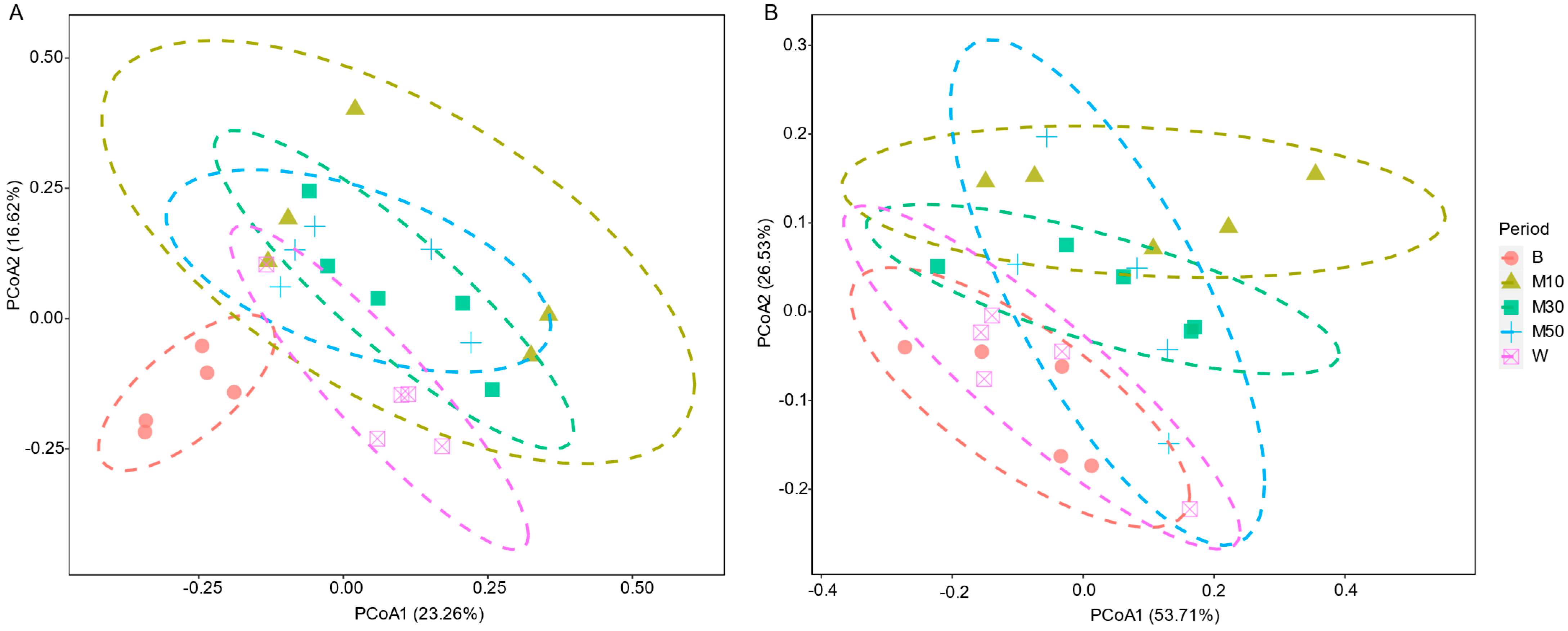
3.1. Bacterial Community Composition and Diversity
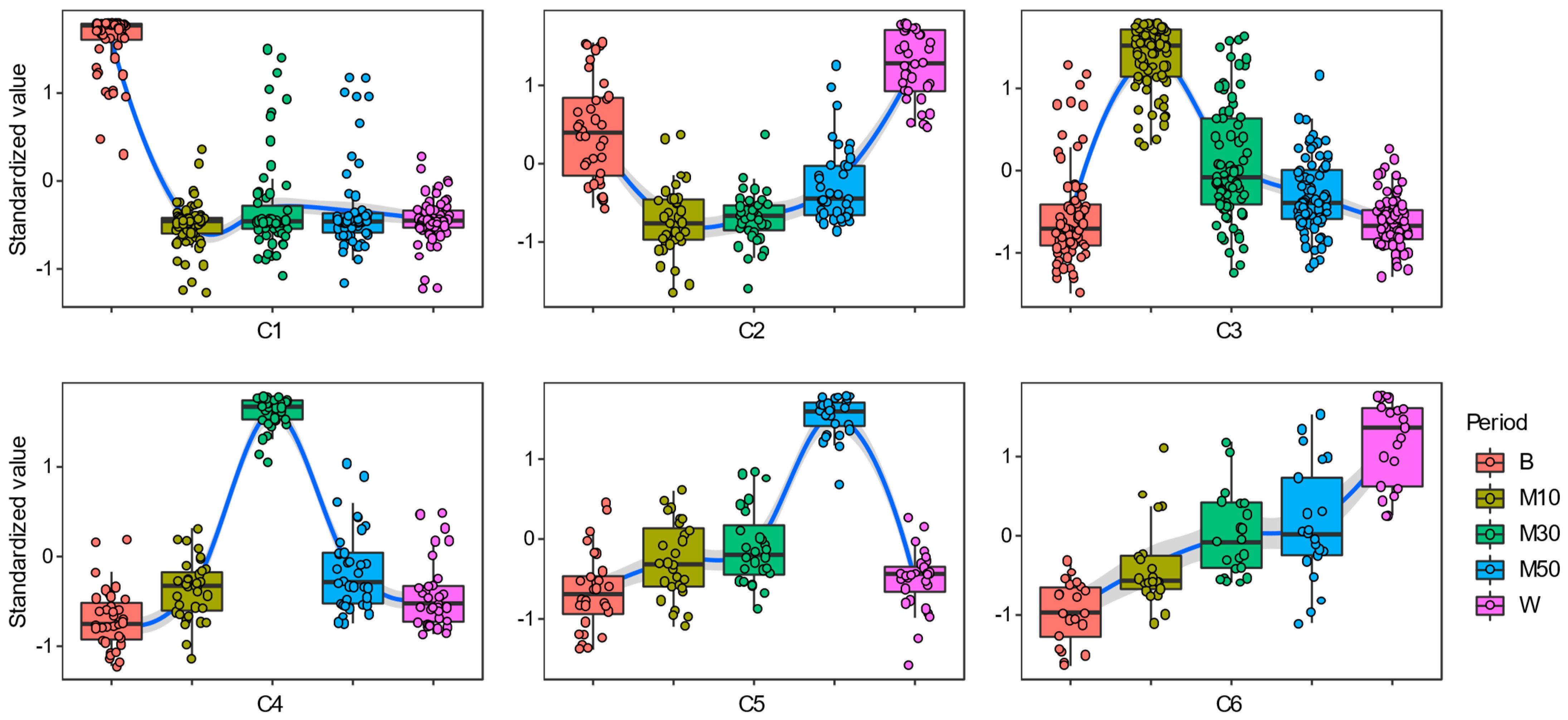
3.2. Succession Patterns of Bacterial Communities
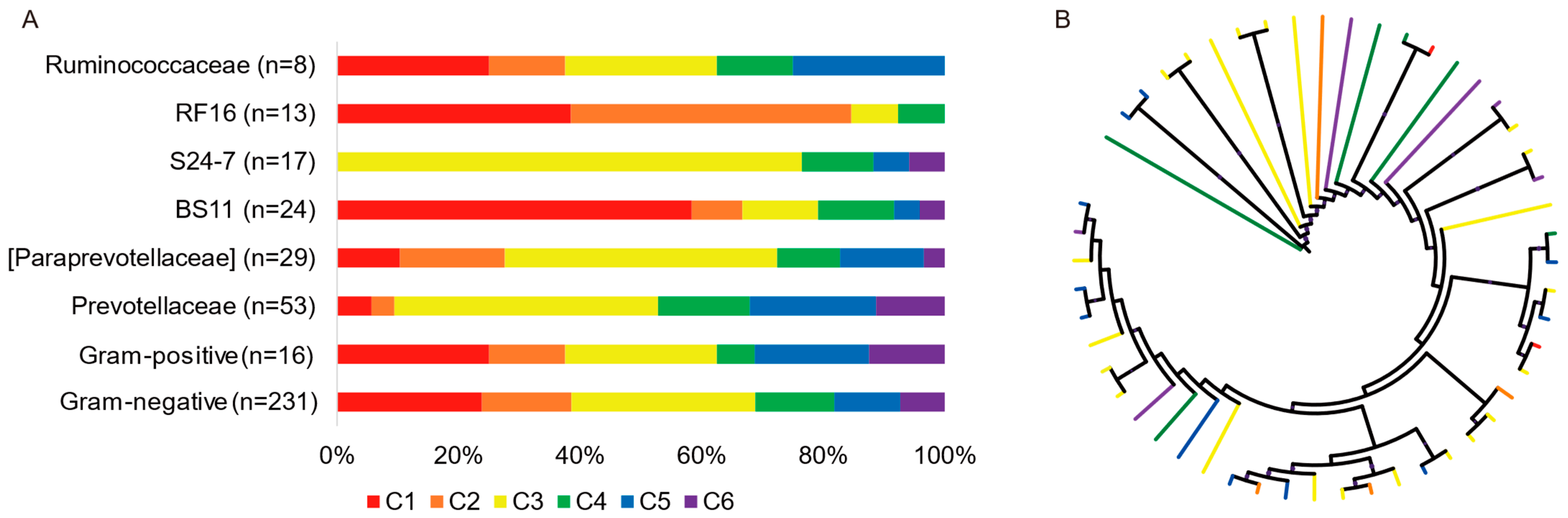
3.3. Succession Patterns of Amylolytic and Cellulolytic Genera
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Russell, J.B.; Rychlik, J.L. Factors that alter rumen microbial ecology. Science 2001, 292, 1119–1122. [Google Scholar] [CrossRef] [PubMed]
- Cottle, D.; Kahn, L. Beef Cattle Production and Trade; CSIRO Publishing: Clayton, Australia, 2014; p. 221. [Google Scholar]
- McGuffey, R.K.; Richardson, L.F.; Wilkinson, J.I.D. Ionophores for dairy cattle: Current status and future outlook. J. Dairy Sci. 2001, 84, E194–E203. [Google Scholar] [CrossRef]
- Duffield, T.F.; Merrill, J.K.; Bagg, R.N. Meta-analysis of the effects of monensin in beef cattle on feed efficiency, body weight gain, and dry matter intake. J. Anim. Sci. 2012, 90, 4583–4592. [Google Scholar] [CrossRef] [PubMed]
- Hristov, A.N.; Oh, J.; Lee, C.; Meinen, R. Mitigation of Greenhouse Gas Emissions in Livestock Production: A Review of Technical Options for Non-CO2 Emissions; FAO Animal Production and Health Paper; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; ISBN 978-92-5-107658-3. [Google Scholar]
- Hayes, D.J.; Jensen, H.H.; Fabiosa, J. Technology choice and the economic effects of a ban on the use of antimicrobial feed additives in swine rations. Food Control 2002, 13, 97–101. [Google Scholar] [CrossRef]
- Casewell, M.; Friis, C.; Marco, E.; McMullin, P.; Phillips, I. The European ban on growth-promoting antibiotics and emerging consequences for human and animal health. J. Antimicrob. Chemother. 2003, 52, 159–161. [Google Scholar] [CrossRef] [PubMed]
- Khorrami, B.; Vakili, A.R.; Mesgaran, M.D.; Klevenhusen, F. Thyme and cinnamon essential oils: Potential alternatives for monensin as a rumen modifier in beef production systems. Anim. Feed Sci. Technol. 2015, 200, 8–16. [Google Scholar] [CrossRef]
- Morsy, A.S.; Soltan, Y.A.; Sallam, S.M.A.; Kreuzer, M.; Alencar, S.M.; Abdalla, A.L. Comparison of the in vitro efficiency of supplementary bee propolis extracts of different origin in enhancing the ruminal degradability of organic matter and mitigating the formation of methane. Anim. Feed Sci. Technol. 2015, 199, 51–60. [Google Scholar] [CrossRef]
- Shen, J.; Liu, Z.; Yu, Z.; Zhu, W. Monensin and nisin affect rumen fermentation and microbiota differently in vitro. Front. Microbiol. 2017, 8, 1111. [Google Scholar] [CrossRef]
- Pan, X.; Cai, Y.; Li, Z.; Chen, X.; Heller, R.; Wang, N.; Wang, Y.; Zhao, C.; Wang, Y.; Xu, H.; et al. Modes of genetic adaptations underlying functional innovations in the rumen. Sci. China Life Sci. 2021, 64, 1–21. [Google Scholar] [CrossRef]
- Li, Z.; Lei, X.; Chen, X.; Yin, Q.; Shen, J.; Yao, J. Long-term and combined effects of N-[2-(Nitrooxy)Ethyl]-3-Pyridinecarboxamide and fumaric acid on methane production, rumen fermentation, and lactation performance in dairy goats. J. Anim. Sci. Biotechnol. 2021, 12, 125. [Google Scholar] [CrossRef]
- Russell, J.B.; Houlihan, A.J. Ionophore resistance of ruminal bacteria and its potential impact on human health. FEMS Microbiol. Rev. 2003, 27, 65–74. [Google Scholar] [CrossRef]
- Patra, A.; Park, T.; Kim, M.; Yu, Z. Rumen methanogens and mitigation of methane emission by anti-methanogenic compounds and substances. J. Anim. Sci. Biotechnol. 2017, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Myer, P.R. Bovine Genome-Microbiome Interactions: Metagenomic frontier for the selection of efficient productivity in cattle systems. mSystems 2019, 4, e00103-19. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Felix, T.L.; Loerch, S.C.; Yu, Z. Effect of haylage and monensin supplementation on ruminal bacterial communities of feedlot cattle. Curr. Microbiol. 2014, 69, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Newbold, C.; Wallace, R.; Walker, N. The effect of tetronasin and monensin on fermentation, microbial numbers and the development of ionophore-resistant bacteria in the rumen. J. Appl. Bacteriol. 1993, 75, 129–134. [Google Scholar] [CrossRef]
- Ravva, S.V.; Sarreal, C.Z.; Mandrell, R.E. Altered protozoan and bacterial communities and survival of Escherichia coli O157:H7 in monensin-treated wastewater from a dairy lagoon. PLoS ONE 2013, 8, e54782. [Google Scholar] [CrossRef]
- Kim, M.; Eastridge, M.L.; Yu, Z. Investigation of ruminal bacterial diversity in dairy cattle fed supplementary monensin alone and in combination with fat, using pyrosequencing analysis. Can. J. Microbiol. 2014, 60, 65–71. [Google Scholar] [CrossRef]
- Guan, H. Efficacy of ionophores in cattle diets for mitigation of enteric methane. J. Anim. Sci. 2006, 84, 1896–1906. [Google Scholar] [CrossRef]
- Li, Z.; Ren, H.; Liu, S.; Cai, C.; Han, J.; Li, F.; Yao, J. Dynamics of methanogenesis, ruminal fermentation, and alfalfa degradation during adaptation to monensin supplementation in goats. J. Dairy Sci. 2018, 101, 1048–1059. [Google Scholar] [CrossRef]
- Ding, J.; Dai, R.; Yang, L.; He, C.; Xu, K.; Liu, S.; Zhao, W.; Xiao, L.; Luo, L.; Zhang, Y.; et al. Inheritance and establishment of gut microbiota in chickens. Front. Microbiol. 2017, 8, 1967. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610. [Google Scholar] [CrossRef] [PubMed]
- Bray, J.R.; Curtis, J.T. An ordination of the upland forest communities of southern wisconsin. Ecol. Monogr. 1957, 27, 326–349. [Google Scholar] [CrossRef]
- Petzel, J.P.; Hartman, P.A. Monensin-based medium for determination of total gram-negative bacteria and Escherichia coli. Appl. Environ. Microbiol. 1985, 49, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, E.; DeLong, E.F.; Lory, S.; Stackebrandt, E.; Thompson, F. (Eds.) The Prokaryotes: Other Major Lineages of Bacteria and the Archaea, 4th ed.; Springer: Berlin/Heidelberg, Germany, 2014; ISBN 978-3-642-38953-5. [Google Scholar]
- Dworkin, M.; Falkow, S.; Rosenberg, E.; Schleifer, K.-H.; Stackebrandt, E. (Eds.) The Prokaryotes Handbook on the Biology Bacteria, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 2016; Volume 3, ISBN 978-0-387-25476-0. [Google Scholar]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Shade, A.; Caporaso, J.G.; Handelsman, J.; Knight, R.; Fierer, N. A meta-analysis of changes in bacterial and archaeal communities with time. ISME J. 2013, 7, 1493. [Google Scholar] [CrossRef]
- Jiao, S.; Zhang, Z.; Yang, F.; Lin, Y.; Chen, W.; Wei, G. Temporal dynamics of microbial communities in microcosms in response to pollutants. Mol. Ecol. 2017, 26, 923–936. [Google Scholar] [CrossRef]
- McGarvey, J.A.; Place, S.; Palumbo, J.; Hnasko, R.; Mitloehner, F. Dosage-dependent effects of monensin on the rumen microbiota of lactating dairy cattle. MicrobiologyOpen 2018, 8, e00783. [Google Scholar] [CrossRef]
- Jia, P.; Cui, K.; Ma, T.; Wan, F.; Wang, W.; Yang, D.; Wang, Y.; Guo, B.; Zhao, L.; Diao, Q. Influence of dietary supplementation with bacillus licheniformis and saccharomyces cerevisiae as alternatives to monensin on growth performance, antioxidant, immunity, ruminal fermentation and microbial diversity of fattening lambs. Sci. Rep. 2018, 8, 16712. [Google Scholar] [CrossRef] [PubMed]
- Reece, W.O. Dukes’ Physiology of Domestic Animals, 12th ed.; Comstock Pub. Associates: Ithaca, NY, USA, 2004; ISBN 978-0-8014-4238-4. [Google Scholar]
- Li, Z.; Deng, Q.; Liu, Y.; Yan, T.; Li, F.; Cao, Y.; Yao, J. Dynamics of methanogenesis, ruminal fermentation and fiber digestibility in ruminants following elimination of protozoa: A meta-analysis. J. Anim. Sci. Biotechnol. 2018, 9, 89. [Google Scholar] [CrossRef] [PubMed]
- Callaway, T.R.; Russell, J.B. Variations in the ability of ruminal gram-negative prevotella species to resist monensin. Curr. Microbiol. 2000, 40, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Millen, D.D.; Arrigoni, M.D.B.; Pacheco, R.D.L. (Eds.) Rumenology, 1st ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; ISBN 978-3-319-30531-8. [Google Scholar]
- Ormerod, K.L.; Wood, D.L.A.; Lachner, N.; Gellatly, S.L.; Daly, J.N.; Parsons, J.D.; Dal’Molin, C.G.O.; Palfreyman, R.W.; Nielsen, L.K.; Cooper, M.A.; et al. Genomic characterization of the uncultured bacteroidales family S24-7 inhabiting the guts of homeothermic animals. Microbiome 2016, 4, 36. [Google Scholar] [CrossRef]
- Solden, L.M.; Hoyt, D.W.; Collins, W.B.; Plank, J.E.; Daly, R.A.; Hildebrand, E.; Beavers, T.J.; Wolfe, R.; Nicora, C.D.; Purvine, S.O.; et al. New roles in hemicellulosic sugar fermentation for the uncultivated bacteroidetes family BS11. ISME J. 2017, 11, 691–703. [Google Scholar] [CrossRef]
- Stewart, R.D.; Auffret, M.D.; Warr, A.; Walker, A.W.; Roehe, R.; Watson, M. Compendium of 4,941 Rumen metagenome-assembled genomes for rumen microbiome biology and enzyme discovery. Nat. Biotechnol. 2019, 37, 953–961. [Google Scholar] [CrossRef]
- Sylvester, J.T.; Karnati, S.K.R.; Dehority, B.A.; Morrison, M.; Smith, G.L.; St-Pierre, N.R.; Firkins, J.L. Rumen ciliated protozoa decrease generation time and adjust 18S ribosomal dna copies to adapt to decreased transfer interval, starvation, and monensin. J. Dairy Sci. 2009, 92, 256–269. [Google Scholar] [CrossRef]
- Li, Z.; Wang, X.; Zhang, Y.; Yu, Z.; Zhang, T.; Dai, X.; Pan, X.; Jing, R.; Yan, Y.; Liu, Y.; et al. Genomic insights into the phylogeny and biomass-degrading enzymes of rumen ciliates. ISME J. 2022. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OTUs | Family | |||
|---|---|---|---|---|
| R | p | R | p | |
| Total | 0.345 | 0.001 | 0.154 | 0.032 |
| B vs. M10 | 0.664 | 0.012 | 0.524 | 0.007 |
| B vs. M30 | 0.848 | 0.006 | 0.304 | 0.079 |
| B vs. M50 | 0.816 | 0.009 | 0.392 | 0.025 |
| B vs. W | 0.716 | 0.011 | 0.000 | 0.412 |
| M10 vs. W | 0.244 | 0.055 | 0.368 | 0.024 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.; Liu, Y.; Jiang, Y.; Yao, J.; Li, Z. Ruminal Bacterial Community Successions in Response to Monensin Supplementation in Goats. Animals 2022, 12, 2291. https://doi.org/10.3390/ani12172291
Guo X, Liu Y, Jiang Y, Yao J, Li Z. Ruminal Bacterial Community Successions in Response to Monensin Supplementation in Goats. Animals. 2022; 12(17):2291. https://doi.org/10.3390/ani12172291
Chicago/Turabian StyleGuo, Xi, Yuqin Liu, Yu Jiang, Junhu Yao, and Zongjun Li. 2022. "Ruminal Bacterial Community Successions in Response to Monensin Supplementation in Goats" Animals 12, no. 17: 2291. https://doi.org/10.3390/ani12172291