
Chemical Characterization of the Marking Fluid of Breeding and Non-Breeding Male Cheetahs



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Experimental Location and Animals
2.3. Marking Fluid Collection and Storage
2.4. Sample Analysis
2.5. Identification of VOCs
2.6. Statistical Analysis
3. Results and Discussion
3.1. Sample Collection
3.2. Presence of VOCs
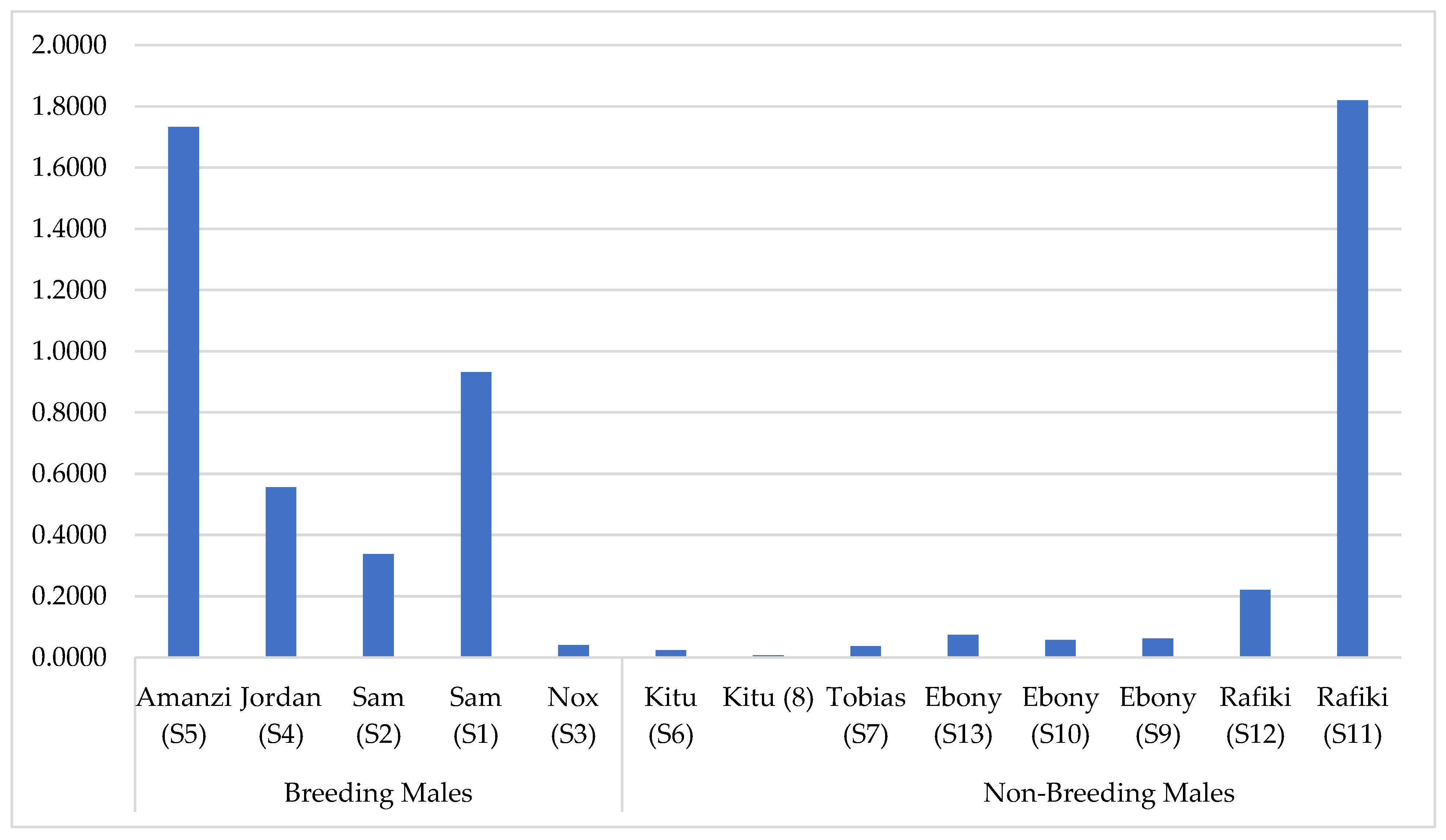
3.3. Factors Affecting Presence of VOCs
3.4. Relative Concentrations of the 12 Common VOCs
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wemmer, C.; Scow, K. Communication in the Felidae with Emphasis on Scent Marking and Contact Marking and Contact Patterns (Book Section). In How Animals Communicate; Sebeok, T.A., Ed.; Indiana University Press: Bloomington, IN, USA, 1977; pp. 749–766. [Google Scholar]
- Wielebnowski, N.; Brown, J.L. Behavioral Correlates of Physiological Estrus in Cheetahs. Zoo Biol. 1998, 17, 193–209. [Google Scholar] [CrossRef]
- Andersen, K.F.; Vulpius, T. Urinary Volatile Constituents of the Lion, Panthera Leo. Chem. Senses 1999, 24, 179–189. [Google Scholar] [CrossRef] [PubMed]
- McLean, L.; Hurst, J.L.; Gaskell, C.J.; Lewis, J.C.M.; Beynon, R.J. Characterization of Cauxin in the Urine of Domestic and Big Cats. J. Chem. Ecol. 2007, 33, 1997–2009. [Google Scholar] [CrossRef]
- Bland, K.P. Tom-Cat Odour and Other Pheromones in Feline Reproduction. Vet. Sci. Commun. 1979, 3, 125–136. [Google Scholar] [CrossRef]
- Brahmachary, R.L.; Poddar-Sarkar, M. Fifty Years of Tiger Pheromone Research. Curr. Sci. 2015, 108, 2178–2185. [Google Scholar]
- Soso, S.B. Chemical Constituency and Odor of Semiochemicals: Studying the Chemical Composition and Odor of Volatile Organic Compounds of Great Cat Marking Fluid in an Effort to Aid Tiger and Lion Conservation. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2016. [Google Scholar]
- Asa, C.S. Relative Contributions of Urine and Anal-Sac Secretions in Scent Marks of Large Felids. Am. Zool. 1993, 33, 167–172. [Google Scholar] [CrossRef]
- Regnier, F.E. Semiochemicals—Structure and Function. Biol. Reprod. 1971, 4, 309–326. [Google Scholar] [CrossRef]
- Poddar-Sarkar, M. Mammalian Semiochemicals: Chemical and Behavioural Aspects with Special Reference to Tiger. Ph.D. Thesis, University of Calcutta, Kolkata, West Bengal, India, 1995. [Google Scholar]
- Visser, R.R.C. Chemical Communication: Chemical Characterization of Volatile Constituents of Urine of the Southern African Cheetah, Acinonyx Jubatus Jubatus, Using Headspace Sampling and GC-MS. Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, 2002. [Google Scholar]
- Soso, S.B.; Koziel, J.A. Analysis of Odorants in Marking Fluid of Siberian Tiger (Panthera Tigris Altaica) Using Simultaneous Sensory and Chemical Analysis with Headspace Solid-Phase Microextraction and Multidimensional Gas Chromatography-Mass Spectrometry-Olfactometry. Molecules 2016, 21, 834. [Google Scholar] [CrossRef]
- Soso, S.B.; Koziel, J.A. Characterizing the Scent and Chemical Composition of Panthera Leo Marking Fluid Using Solid-Phase Microextraction and Multidimensional Gas Chromatography-Mass Spectrometry-Olfactometry. Sci. Rep. 2017, 7, 5137. [Google Scholar] [CrossRef]
- Verberne, G.; de Boer, J. Chemocommunication among Domestic Cats, Mediated by the Olfactory and Vomeronasal Senses: I. Chemocommunication. Z. Tierpsychol. 1976, 42, 86–109. [Google Scholar]
- Mellen, J.D. A Comparative Analysis of Scent-Marking, Social and Reproductive Behavior in 20 Species of Small Cats (Felis). Am. Zool. 1993, 33, 151–166. [Google Scholar] [CrossRef]
- Martínez-Macipe, M.; Lafont-Lecuelle, C.; Manteca, X.; Pageat, P.; Cozzi, A. Evaluation of an Innovative Approach for Sensory Enrichment in Zoos: Semiochemical Stimulation for Captive Lions (Panthera Leo). Anim. Welf. 2015, 24, 455–461. [Google Scholar] [CrossRef]
- Burger, B.V.; Visser, R.; Moses, A.; le Roux, M. Elemental Sulfur Identified in Urine of Cheetah, Acinonyx Jubatus. J. Chem. Ecol. 2006, 32, 1347–1352. [Google Scholar] [CrossRef] [PubMed]
- Mossotti, R.H. Female Reaction to Male Urine Scents as Potential Indicator of Mate Choice in Captive Cheetahs (Acinonyx Jubatus). Ph.D. Thesis, Southern Illinois University, Carbondale, IL, USA, 2010. [Google Scholar]
- Semren, T.Ž.; Karačonji, I.B.; Safner, T.; Brajenović, N.; Lovaković, B.T.; Pizent, A. Gas Chromatographic-Mass Spectrometric Analysis of Urinary Volatile Organic Metabolites: Optimization of the HS-SPME Procedure and Sample Storage Conditions. Talanta 2018, 176, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Pawliszyn, J. Solid Phase Microextraction—Theory and Practice, 1st ed.; Wiley: New York, NY, USA, 1997. [Google Scholar]
- Rostad, C.E.; Pereira, W.E. Kovats and Lee Retention Indices Determined by Gas Chromatography/Mass Spectrometry for Organic Compounds of Environmental Interest. J. High Resolut. Chromatogr. 1986, 9, 328–334. [Google Scholar] [CrossRef]
- Shellie, R.; Mondello, L.; Marriott, P.; Dugo, G. Characterisation of Lavender Essential Oils by Using Gas Chromatography-Mass Spectrometry with Correlation of Linear Retention Indices and Comparison with Comprehensive Two-Dimensional Gas Chromatography. J. Chromatogr. A 2002, 970, 225–234. [Google Scholar] [CrossRef]
- Eckel, W.P.; Kind, T. Use of Boiling Point-Lee Retention Index Correlation for Rapid Review of Gas Chromatography-Mass Spectrometry Data. Anal. Chim. Acta. 2003, 494, 235–243. [Google Scholar] [CrossRef]
- van den Dool, H.; Kratz, P.; Dec, A. Generalization of the Retention Index System Including Linear Temperature Programmed Gas-Liquid Partition Chromatography. J. Chromatogr. 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Linstrom, P.J.; Mallard, W.G. The NIST Chemistry WebBook. Linstrom, P.J., Mallard, W.G., Eds.; National Institute of Standards and Technology: Gaithersburg, MD, USA, 2001; Volume 46. [Google Scholar]
- Apfelbach, R.; Parsons, M.H.; Soini, H.A.; Novotny, M.V. Are Single Odorous Components of a Predator Sufficient to Elicit Defensive Behaviors in Prey Species? Front Neurosci. 2015, 9, 263. [Google Scholar] [CrossRef]
- Manrique, G.; Lorenzo, M. The Sexual Behaviour of Chagas’ Disease Vectors: Chemical Signals Mediating Communication between Male and Female Triatomine Bugs. Psyche (Stuttg) 2012. [CrossRef]
- Manrique, G.; Vitta, A.C.R.; Ferreira, R.A.; Zani, C.L.; Unelius, C.R.; Lazzari, C.R.; Diotaiuti, L.; Lorenzo, M.G. Chemical Communication in Chagas Disease Vectors. Source, Identity, and Potential Function of Volatiles Released by the Metasternal and Brindley’s Glands of Triatoma Infestans Adults. J. Chem. Ecol. 2006, 32, 2035–2052. [Google Scholar] [CrossRef] [PubMed]
- Bertschinger, H.J.; Meltzer, D.G.A.; van Dyk, A. Captive Breeding of Cheetahs in South Africa-30 Years of Data from the de Wildt Cheetah and Wildlife Centre. Reprod. Domest. Anim. 2008, 43, 66–73. [Google Scholar] [CrossRef]
- Gopas, J.; Abd El Qader, A.; Isaacson, C.; Eichler, D.; Zeiri, Y. Urine Volatile Organic Compounds Composition in Mice Bearing Breast and Melanoma Tumors: Effect of Low-Protein Diet. Curr. Biomark. Find. 2018, 8, 1–13. [Google Scholar] [CrossRef]
- Penn, D.J.; Oberzaucher, E.; Grammer, K.; Fischer, G.; Soini, H.A.; Wiesler, D.; Novotny, M.V.; Dixon, S.J.; Xu, Y.; Brereton, R.G. Individual and Gender Fingerprints in Human Body Odour. J. R. Soc. Interface 2007, 4, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Starkenmann, C.; Niclass, Y.; Cayeux, I.; Brauchli, R.; Gagnon, A.C. Odorant Volatile Sulfur Compounds in Cat Urine: Occurrence of (+/-)-3,7-Dimethyloct-3-Sulfanyl-6-En-1-Ol and Its Cysteine Conjugate Precursor. Flavour. Fragr. J. 2015, 30, 91–100. [Google Scholar] [CrossRef]
- Uetake, K.; Abumi, T.; Suzuki, T.; Hisamatsu, S.; Fukuda, M. Volatile Faecal Components Related to Sex and Age in Domestic Cats (Felis Catus). J. Appl. Anim. Res. 2018, 46, 766–770. [Google Scholar] [CrossRef]
- Kowacich, D.; Hatano, E.; Schal, C.; Ponnusamy, L.; Apperson, C.S.; Shymanovich, T.; Wasserberg, G. The Egg and Larval Pheromone Dodecanoic Acid Mediates Density-Dependent Oviposition of Phlebotomus Papatasi. Parasit. Vectors 2020, 13, 1–11. [Google Scholar] [CrossRef]
- Goitom, A.H. Identification of the Putative Urinary Intraspecific Recognition Pheromone of the Caracal. Ph.D. Thesis, University of Stellenbosch, Stellenbosch, South Africa, 2017. [Google Scholar]
- Burger, B.V.; Viviers, M.Z.; Bekker, J.P.I.; le Roux, M.; Fish, N.; Fourie, W.B.; Weibchen, G. Chemical Characterization of Territorial Marking Fluid of Male Bengal Tiger, Panthera Tigris. J. Chem. Ecol. 2008, 34, 659. [Google Scholar] [CrossRef]
- Carrel, J.E. Aphrodisiacs and Anaphrodisiacs: Better Loving through Chemistry. Stadler Genet. Symp. 1976, 8, 79–109. [Google Scholar]
- Apps, P.J.; Weldon, P.J.; Kramer, M. Chemical Signals in Terrestrial Vertebrates: Search for Design Features. Nat. Prod. Rep. 2015, 32, 1131–1153. [Google Scholar] [CrossRef]
- Rajanarayanan, S.; Archunan, G. Identification of Urinary Sex Pheromones in Female Buffaloes and Their Influence on Bull Reproductive Behaviour. Res. Vet. Sci. 2011, 91, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Pugliese, G.; Carrito, M.; Moura, C.; Vasconcelos, P.; Cera, N.; Li, M.; Nobre, P.; Georgiadis, J.R.; Schubert, J.K.; et al. Breath Chemical Markers of Sexual Arousal in Humans. Sci. Rep. 2022, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Manna, S.; Ray, S.; Das, P.; Rai, U.; Ghosh, B.; Sarkar, M.P. Do Urinary Volatiles Carry Communicative Messages in Himalayan Snow Leopards [Panthera Uncia, (Schreber, 1775)]? In Chemical Signals in Vertebrates 14; Springer: Cham, Germany, 2019; pp. 27–37. [Google Scholar]
- Singer, A.G.; Agosta, W.C.; O’Connell, R.J.; Pfaffmann, C.; Bowen, D.V.; Field, F.H. Dimethyl Disulfide: An Attractant Pheromone in Hamster Vaginal Secretion. Science (1979) 1976, 191, 948–950. [Google Scholar] [CrossRef] [PubMed]
- Dzięcioł, M.; Woszczylo, M.; Szumny, A.; Jezierski, T.; Kupczyński, R.; Godzińska, E.J.; Pieczewska, B.; Niżański, W. Identification of Putative Volatile Sex Pheromones in Female Domestic Dogs (Canis Familiaris). Anim. Reprod. Sci. 2018, 197, 87–92. [Google Scholar] [CrossRef] [PubMed]
- McLean, S.; Nichols, D.S.; Davies, N.W. Volatile Scent Chemicals in the Urine of the Red Fox, Vulpes Vulpes. PLoS ONE 2021, 16, e0248961. [Google Scholar]
- Janda, E.D.; Perry, K.L.; Hankinson, E.; Walker, D.; Vaglio, S. Sex Differences in Scent-Marking in Captive Red-Ruffed Lemurs. Am. J. Primatol. 2019, 81, e22951. [Google Scholar] [CrossRef]
- Tommasi, A.; Koziel, J.A.; Molotsi, A.H.; Esposito, G. Understanding the Role of Semiochemicals on the Reproductive Behaviour of Cheetahs (Acinonyx Jubatus)—A Review. Animals 2021, 11, 3140. [Google Scholar] [CrossRef]
- Banik, C.; Koziel, J.A.; Li, J.Z. Simultaneous Chemical and Sensory Analysis of Domestic Cat Urine and Feces with Headspace Solid-Phase Microextraction and Gc-Ms-Olfactometry. Separations 2021, 8, 15. [Google Scholar] [CrossRef]
- Lee, M.; Koziel, J.A.; Murphy, W.; Jenks, W.S.; Chen, B.; Li, P.; Banik, C. Evaluation of TiO2 Based Photocatalytic Treatment of Odor and Gaseous Emissions from Swine Manure with UV-A and UV-C. Animals 2021, 11, 1289. [Google Scholar] [CrossRef]
- Lee, M.; Li, P.; Koziel, J.A.; Ahn, H.; Wi, J.; Chen, B.; Meiirkhanuly, Z.; Banik, C.; Jenks, W.S. Pilot-Scale Testing of UV-A Light Treatment for Mitigation of NH3, H2S, GHGs, VOCs, Odor, and O3 Inside the Poultry Barn. Front. Chem. 2020, 8, 613. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Functional Group | Sample in Which the VOC Was Identified | p | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Breeding Males (8.9 ± 1.3y) | Non-Breeding Males (5.5 ± 0.8y) | |||||||||||||||
| Name of Identified VOC | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | |||
| Ketones | 1 | 3-hexanone | ✓ | ✓ | ✓ | ✓ | 1.00 | |||||||||
| 2 | 3-pentanone | ✓ | ✓ | ✓ | ✓ | 0.10 | ||||||||||
| 3 | 4-heptanone | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 0.13 | |||
| 4 | 3-heptanone | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 0.51 | ||||
| 5 | 2-heptanone | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 0.22 | |||||
| 6 | 3-octanone | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1.00 | ||
| 7 | 3-ethylcyclopentanone | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 0.40 | |
| 8 | 6-methyl-5-hepten-2-one | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 0.27 | |||||||
| 9 | 3-ethylcyclopent-2-en-1-one | ✓ | 0.38 | |||||||||||||
| 10 | 2-Undecanone | ✓ | ✓ | ✓ | 0.49 | |||||||||||
| 11 | Benzyl methyl ketone | ✓ | ✓ | ✓ | ✓ | 1.00 | ||||||||||
| 12 | 1-phenyl-2-propanone | ✓ | ✓ | 0.13 | ||||||||||||
| 13 | Acetophenone | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1.00 | |
| 14 | Tetrahydro-6-pentyl-2H-pyran-2-one | ✓ | ✓ | ✓ | 0.49 | |||||||||||
| 15 | Delta octalactone | ✓ | 0.38 | |||||||||||||
| Aldehydes | 16 | Hexanal | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1.00 |
| 17 | Furfural | ✓ | ✓ | ✓ | ✓ | 1.00 | ||||||||||
| 18 | 2-furancarboxaldehyde | ✓ | ✓ | 0.13 | ||||||||||||
| 19 | Decanal | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1.00 | ||||
| 20 | Benzaldehyde | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1.00 | |
| 21 | 2,6,6-trimethyl-1-cyclohexene-1-carboxaldehyde | ✓ | 0.38 | |||||||||||||
| 22 | Hexadecanal | ✓ | ✓ | 0.49 | ||||||||||||
| Esters | 23 | Ethyl pentanoate | ✓ | 1.00 | ||||||||||||
| 24 | Ethyl hexanoate | ✓ | 1.00 | |||||||||||||
| 25 | Methyl hexadecanoate | ✓ | ✓ | ✓ | 0.51 | |||||||||||
| Alcohols | 26 | 1-octen-3-ol | ✓ | 1.00 | ||||||||||||
| 27 | 2-ethylhexanol | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 0.29 | ||||||
| 28 | 2-furanmethanol | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1.00 | |
| 29 | α-methyl benzenemethanol | ✓ | ✓ | ✓ | 1.00 | |||||||||||
| 30 | Phenylethyl alcohol | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 0.29 | ||||||
| 31 | Phenol | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1.00 | |
| Acids | 32 | Hexanoic acid | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1.00 | |||||||
| 33 | Heptanoic Acid | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 0.59 | |||||||
| 34 | Octanoic acid | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1.00 | |
| 35 | Nonanoic Acid | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1.00 | |
| 36 | Decanoic acid | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 0.49 | |||
| 37 | Undecanoic acid | ✓ | ✓ | ✓ | ✓ | ✓ | 0.29 | |||||||||
| 38 | Dodecanoic acid | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1.00 | |
| 39 | Hexadecanoic acid | ✓ | ✓ | ✓ | 1.00 | |||||||||||
| Hydrocarbons | 40 | Styrene | ✓ | ✓ | 1.00 | |||||||||||
| 41 | Octadecane | ✓ | 0.38 | |||||||||||||
| 42 | 2-methylnaphthalene | ✓ | 0.38 | |||||||||||||
| Sulfur containing compounds | 43 | Dimethyl disulfide | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1.00 |
| 44 | 2-methylmercaptofuran | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1.00 | |
| 45 | Dimethyl trisulfide | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1.00 | |||
| Nitrogen-Containing Compounds | 46 | 1-methyl-1H-pyrrole | ✓ | 1.00 | ||||||||||||
| 47 | 2-ethenyl-6-methyl-pyrazine | ✓ | 1.00 | |||||||||||||
| 48 | Ethyl carbamate | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 0.59 | ||||||||
| 49 | 2-piperidinone | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 0.59 | ||||||||
| 50 | Indole | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | ✓ | 1.00 | |
| Terpenoids | 51 | Linalool | ✓ | 1.00 | ||||||||||||
| 52 | α-terpineol | ✓ | ✓ | ✓ | 0.49 | |||||||||||
| Furans | 53 | 2-pentyl-furan | ✓ | ✓ | ✓ | ✓ | ✓ | 0.56 | ||||||||
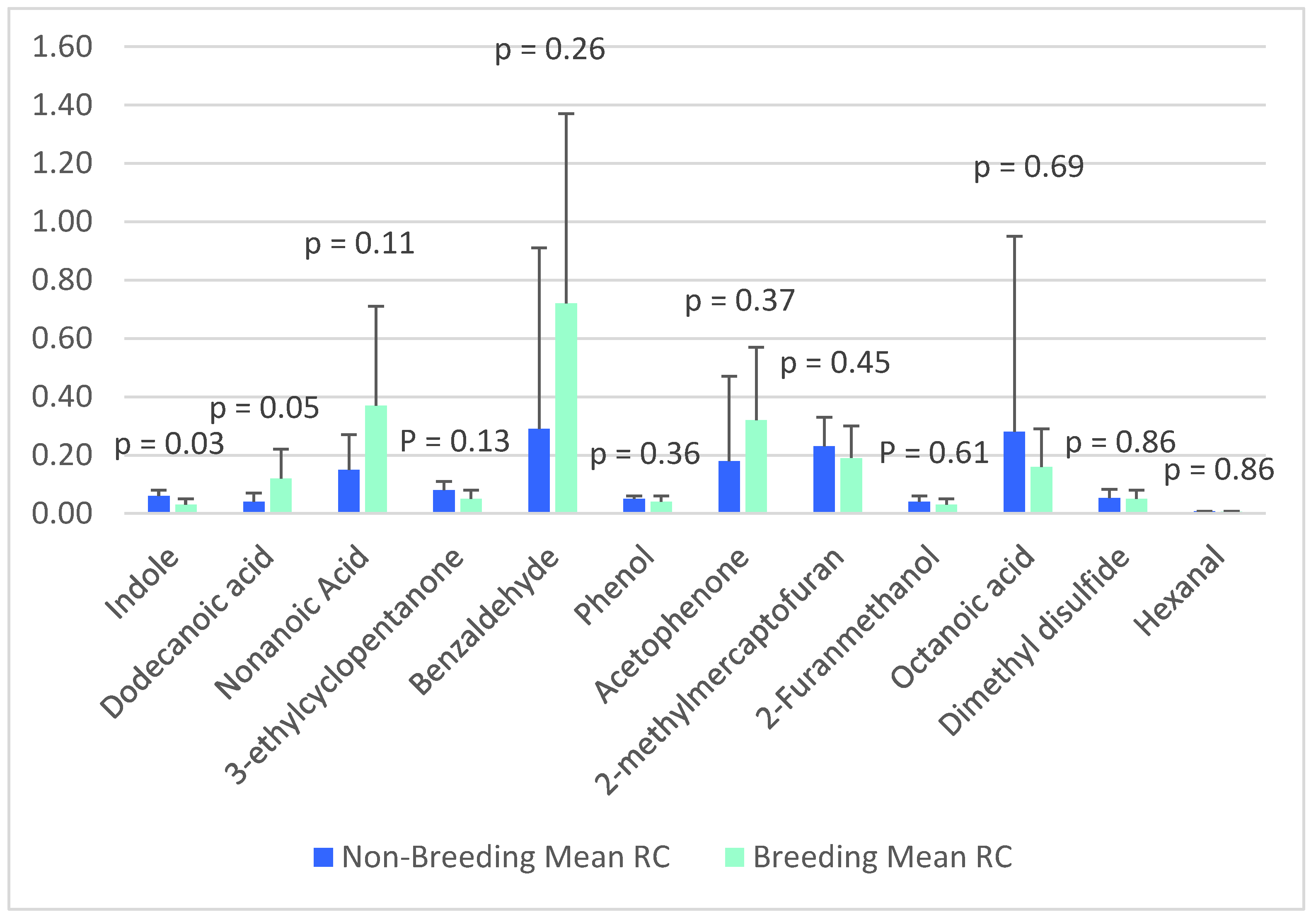
| VOC | RC (Unitless) | |
|---|---|---|
| Non-Breeding (5.5 ± 0.8y) | Breeding (8.9 ± 1.3y) | |
| Dimethyl disulfide | 0.05 ± 0.03 | 0.05 ± 0.03 |
| Hexanal | 0.01 ± 0.00 | 0.01 ± 0.00 |
| 2-methylmercaptofuran | 0.23 ± 0.10 | 0.19 ± 0.11 |
| 3-ethylcyclopentanone | 0.08 ± 0.03 | 0.05 ± 0.03 |
| Benzaldehyde | 0.29 ± 0.62 | 0.72 ± 0.65 |
| Acetophenone | 0.18 ± 0.29 | 0.32 ± 0.25 |
| 2-Furanmethanol | 0.04 ± 0.02 | 0.03 ± 0.02 |
| Phenol | 0.05 ± 0.01 | 0.04 ± 0.02 |
| Octanoic acid | 0.28 ± 0.67 | 0.16 ± 0.13 |
| Nonanoic acid | 0.15 ± 0.12 | 0.37 ± 0.34 |
| Indole | 0.06 ± 0.02 | 0.03 ± 0.02 |
| Dodecanoic acid | 0.04 ± 0.03 | 0.12 ± 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tommasi, A.; Tredoux, A.G.J.; Koziel, J.A.; Esposito, G. Chemical Characterization of the Marking Fluid of Breeding and Non-Breeding Male Cheetahs. Animals 2022, 12, 2284. https://doi.org/10.3390/ani12172284
Tommasi A, Tredoux AGJ, Koziel JA, Esposito G. Chemical Characterization of the Marking Fluid of Breeding and Non-Breeding Male Cheetahs. Animals. 2022; 12(17):2284. https://doi.org/10.3390/ani12172284
Chicago/Turabian StyleTommasi, Alexia, Andreas G. J. Tredoux, Jacek A. Koziel, and Giulia Esposito. 2022. "Chemical Characterization of the Marking Fluid of Breeding and Non-Breeding Male Cheetahs" Animals 12, no. 17: 2284. https://doi.org/10.3390/ani12172284