
Anti-Predation Responses to Conspecific versus Heterospecific Alarm Calls by the Nestlings of Two Sympatric Birds


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Subjects
2.2. Production of Playback Sounds
2.3. Playback Experiments
2.4. Statistical Analysis
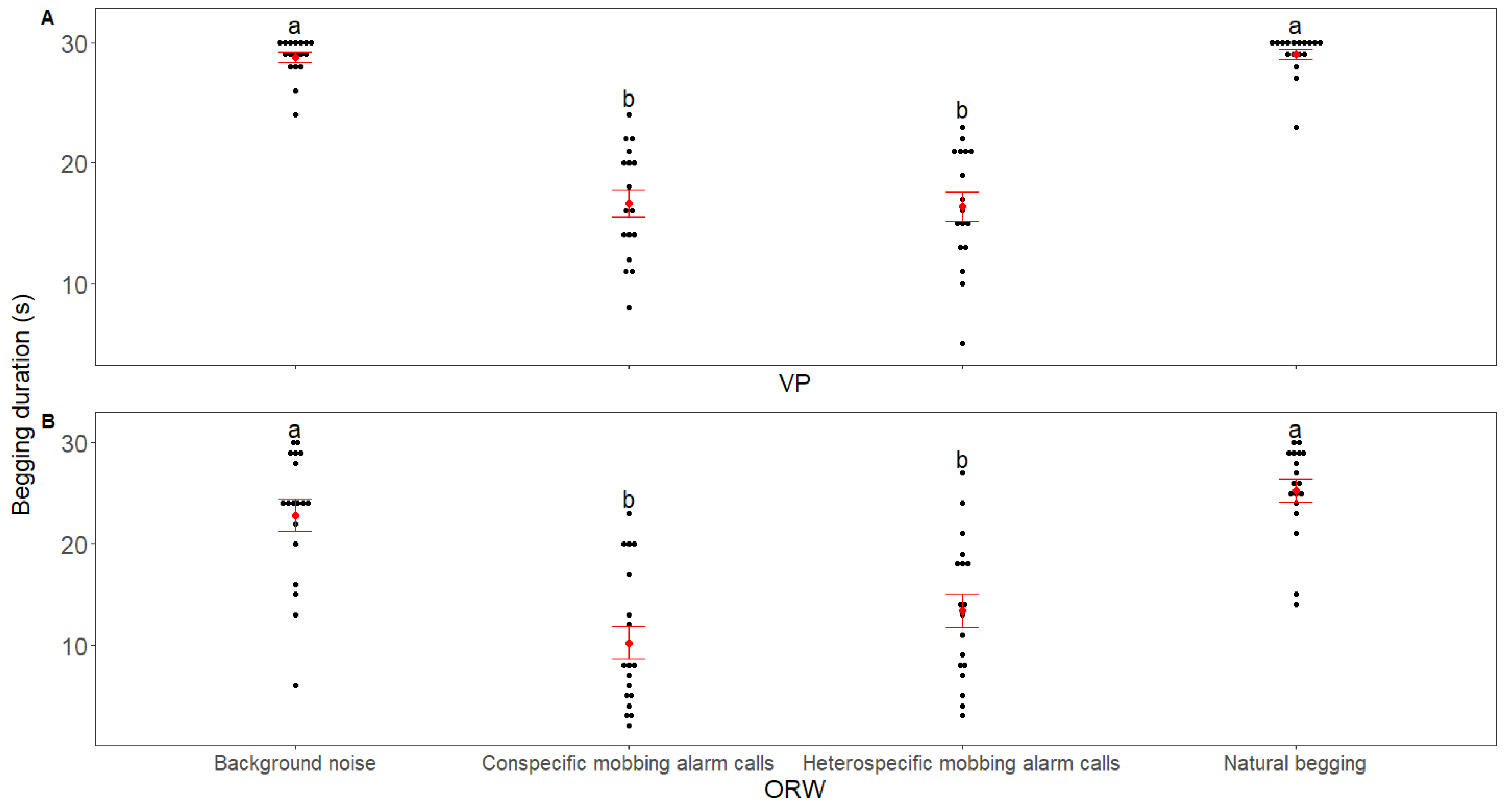
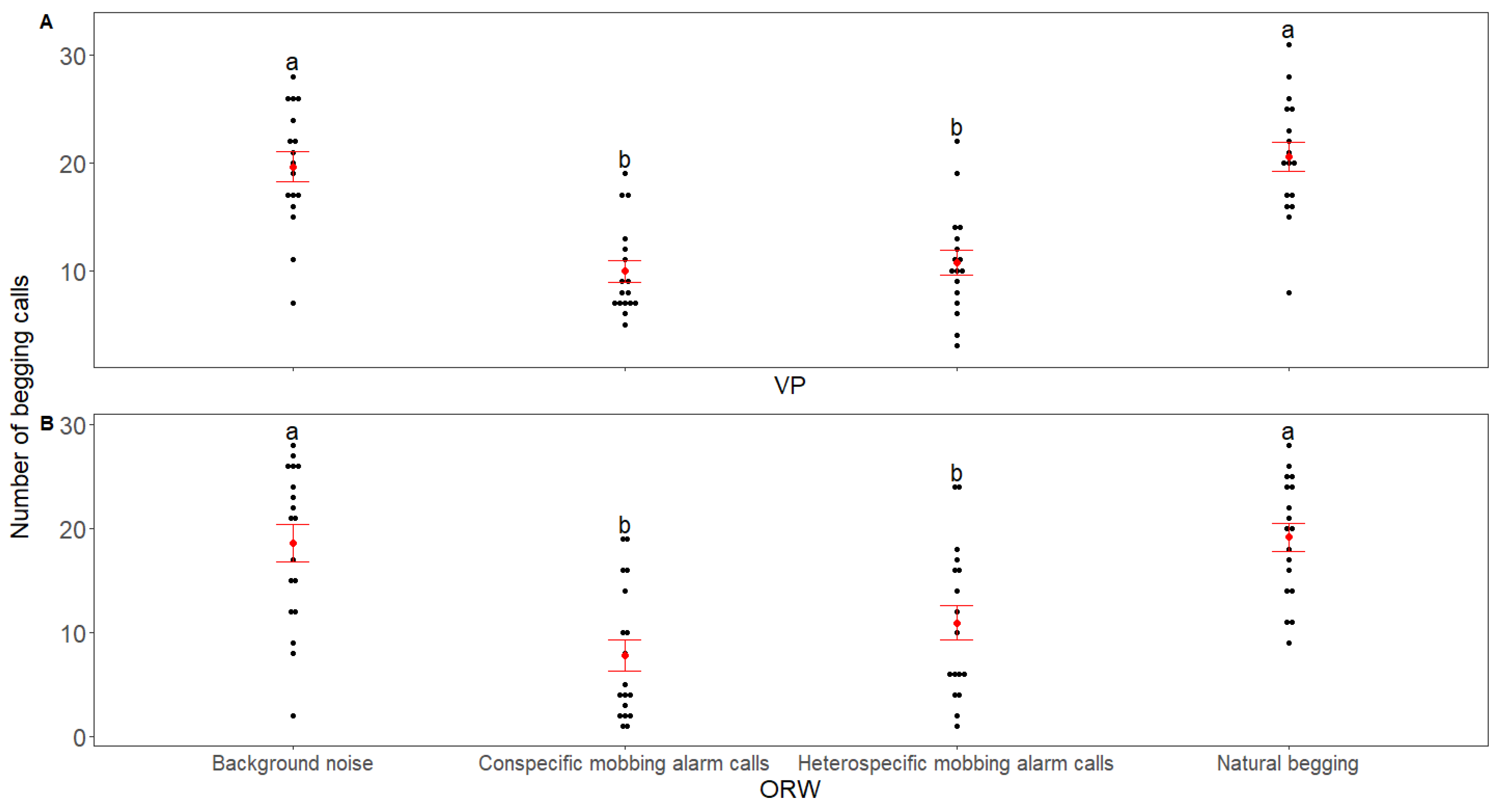
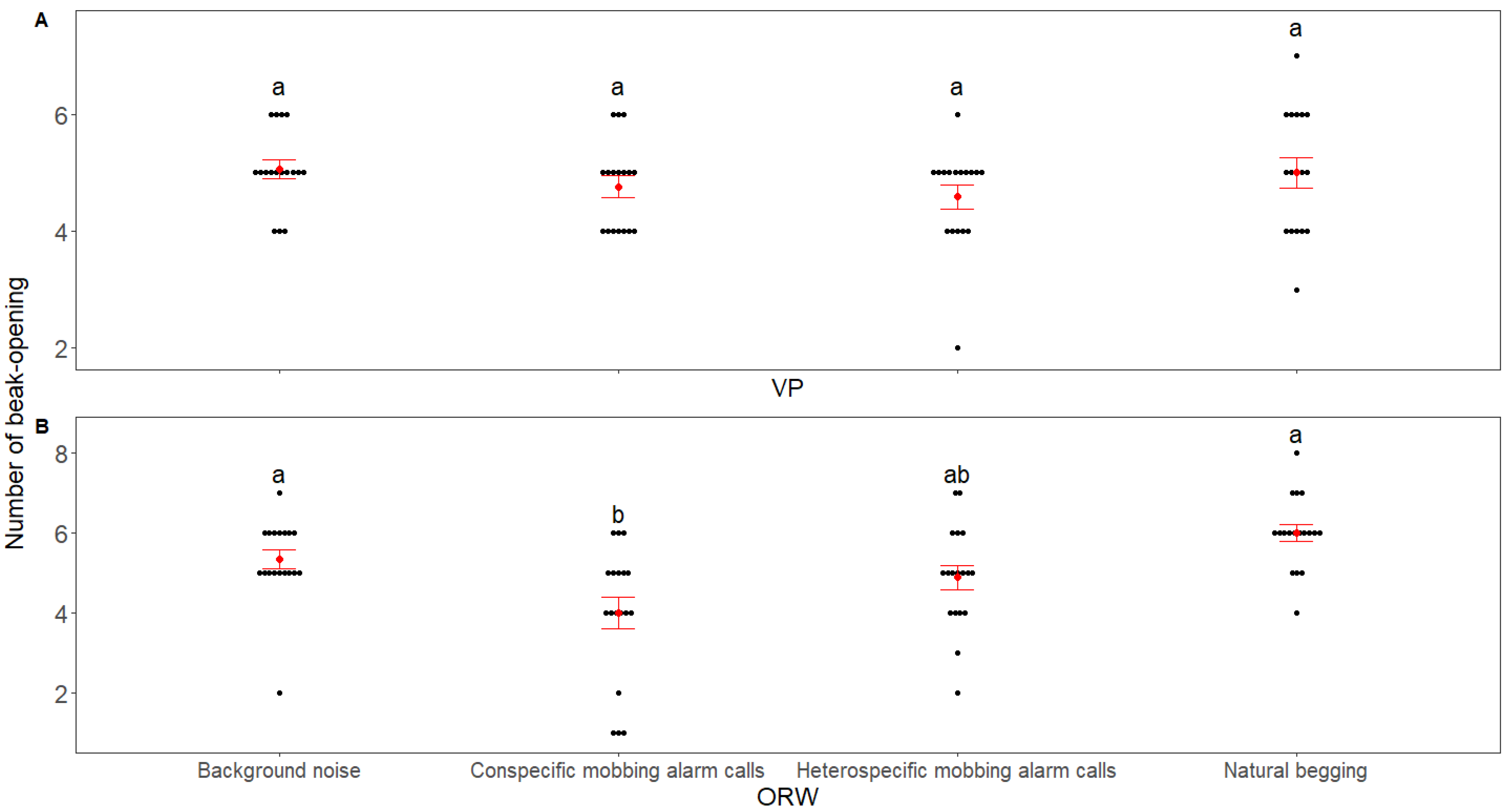
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caro, T.M. Antipredatory Defenses in Birds and Mammals; The University Chicago: Chicago, IL, USA, 2005. [Google Scholar]
- Zuberbühler, K. Predator-specific alarm calls in Campbell’s monkeys, Cercopithecus campbelli. Behav. Ecol. Sociobiol. 2001, 50, 414–422. [Google Scholar] [CrossRef]
- Gill, S.A.; Bierema, A.M.K. On the meaning of alarm calls: A review of functional reference in avian alarm calling. Ethology 2013, 119, 449–461. [Google Scholar] [CrossRef]
- Suzuki, T.N.; Wheatcroft, D.; Griesser, M. Experimental evidence for compositional syntax in bird calls. Nat. Commun. 2016, 7, 10986. [Google Scholar] [CrossRef] [Green Version]
- Manser, M.B. The acoustic structure of suricates’ alarm calls varies with predator type and the level of response urgency. Proceed. R. Soc. B 2001, 268, 2315–2324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fong, J.M.; Navea, F.; Labra, A. Does Liolaemus lemniscatus eavesdrop on the distress calls of the sympatric weeping lizard? J. Ethol. 2021, 39, 11–17. [Google Scholar] [CrossRef]
- Magrath, R.D.; Haff, T.M.; Fallow, P.M.; Radford, A.N. Eavesdropping on heterospecific alarm calls: From mechanisms to consequences. Biol. Rev. 2015, 90, 560–586. [Google Scholar] [CrossRef] [PubMed]
- Kalb, N.; Randler, C. Behavioral responses to conspecific mobbing calls are predator-specific in great tits (Parus major). Ecol. Evol. 2019, 9, 9207–9213. [Google Scholar] [CrossRef] [Green Version]
- McRae, T.R. A review of squirrel alarm-calling behavior: What we know and what we do not know about how predator attributes affect alarm calls. Anim. Behav. Cogn. 2020, 7, 168–191. [Google Scholar] [CrossRef]
- Templeton, C.N.; Greene, E.; Davies, K. Allometry of alarm calls: Black-capped chickadees encode information about predator size. Sicence 2005, 308, 1934–1937. [Google Scholar] [CrossRef] [Green Version]
- Dutour, M.; Walsh, S.L.; Ridley, A.R. Australian magpies adjust their alarm calls according to predator distance. Bioacoustics 2021, 30, 458–468. [Google Scholar] [CrossRef]
- Suzuki, T.N. Other species’ alarm calls evoke a predatorspecific search image in birds. Curr. Biol. 2020, 30, 2616–2620. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.A.; Sealy, S.G. Functional reference in an alarm signal given during nest defence: Seet calls of yellow warblers denote brood-parasitic brown-headed cowbirds. Behav. Ecol. Sociobiol. 2004, 56, 71–80. [Google Scholar] [CrossRef]
- Lawson, S.L.; Enos, J.K.; Mendes, N.C. Heterospecific eavesdropping on an anti-parasitic referential alarm call. Commun. Biol. 2020, 3, 143. [Google Scholar] [CrossRef] [PubMed]
- Carlson, N.V.; Greene, E.; Templeton, C.N. Nuthatches vary their alarm calls based upon the source of the eavesdropped signals. Nat. Commun. 2020, 11, 526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lilly, M.V.; Lucore, E.C.; Tarvin, K.A. Eavesdropping grey squirrels infer safety from bird chatter. PLoS ONE 2019, 14, e0221279. [Google Scholar] [CrossRef] [Green Version]
- Fallow, P.M.; Magrath, R.D. Eavesdropping on other species: Mutual interspecific understanding of urgency information in avian alarm calls. Anim. Behav. 2010, 79, 411–417. [Google Scholar] [CrossRef]
- Davídková, M.; Veselá, P.; Syrová, M.; Nácarová, J.; Bugnyar, T. Ravens respond to unfamiliar corvid alarm calls. J. Ornithol. 2020, 161, 967–975. [Google Scholar] [CrossRef]
- Keen, S.C.; Cole, E.F.; Sheehan, M.J.; Sheldon, B.C. Social learning of acoustic anti-predator cues occurs between wild bird species. Proceed. R. Soc. B 2020, 287, 20192513. [Google Scholar] [CrossRef] [Green Version]
- Farine, D.R.; Aplin, L.M.; Sheldon, B.C.; Hoppitt, W. Interspecific social networks promote information transmission in wild songbirds. R. Soc. 2015, 282, 20142804. [Google Scholar] [CrossRef] [Green Version]
- Danchin, E.; Giraldeau, L.; Valone, T.J.; Wagner, R.H. Public information: From nosy neighbors to cultural evolution. Science 2004, 305, 487–491. [Google Scholar] [CrossRef] [Green Version]
- Avey, M.T.; Hoeschele, M.; Moscicki, M.K.; Bloomfield, L.L.; Sturdy, C.B. Neural correlates of threat perception: Neural equivalence of conspecific and heterospecific mobbing calls is learned. PLoS ONE 2011, 6, e23844. [Google Scholar] [CrossRef] [PubMed]
- Goodale, E.; Beauchamp, G.; Magrath, R.D.; Nieh, J.C.; Ruxton, G.D. Interspecific information transfer influences animal community structure. Trends Ecol. Evol. 2010, 25, 354–361. [Google Scholar] [CrossRef]
- Barati, A.; McDonald, P.G. Nestlings reduce their predation risk by attending to predator-information encoded within conspecific alarm calls. Sci. Rep. 2017, 7, 11736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Platzen, D.; Magrath, R.D. Adaptive differences in response to two types of parental alarm call in altricial nestlings. Proceed. R. Soc. Biol. Sci. 2005, 272, 1101–1106. [Google Scholar] [CrossRef] [Green Version]
- Haff, T.M.; Magrath, R.D. Calling at a cost: Elevated nestling calling attracts predators to active nests. Biol. Lett. 2011, 7, 493–495. [Google Scholar] [CrossRef] [Green Version]
- Kullberg, C.; Lind, J. An experimental study of predator recognition in great tit fledglings. Ethology 2002, 108, 429–441. [Google Scholar] [CrossRef]
- Jiang, Y.X.; Han, J.R.; Zhang, Z.Q.; Chen, X.Y.; Yang, C.C. Parent-offspring and inter-offspring responses to conspecific versus heterospecific distress calls in 2 sympatric birds. Curr. Zool. 2021; in press. [Google Scholar] [CrossRef]
- Wang, Q.X.; Yang, C.; Xiao, H. The breeding ecology of oriental great reed warbler (Acrocephalus orientalis) in hongjiannao of Shaanxi province. Sichuan J. Zool. 2013, 32, 543–546. [Google Scholar]
- Guo, Z.M.; Chen, W.; Hu, J.C. Analysis on nest habitation factors and chick growth of Paradoxornis webbianus. Sichuan J. Zool. 2006, 25, 858–861. [Google Scholar]
- Nakagawa, S.; Schielzeth, H.; O’Hara, R.B. A general and simple method for obtaining R2 from generalized linear mixed-effects models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Haff, T.M.; Magrath, R.D. Learning to listen? Nestling response to heterospecific alarm calls. Anim. Behav. 2012, 84, 1401–1410. [Google Scholar] [CrossRef]
- Whittingham, M.J.; Butler, S.J.; Quinn, J.L. The effect of limited visibility on vigilance behaviour and speed of predator detection: Implications for the conservation of Granivorous passerines. Oikos 2004, 106, 377–385. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameter of Phrase | VP (n = 12) | ORW (n = 11) | w | p |
|---|---|---|---|---|
| Highest frequency (Hz) | 6000.17 ± 332.89 | 6601.91 ± 178.96 | 32 | 0.04 |
| Delta (Hz) | 3945.6 ± 306.27 | 4535.47 ± 190.47 | 24 | 0.008 |
| t | p | |||
| Duration (s) | 0.08 ± 0.004 | 0.15 ± 0.023 | −2.91 | 0.02 |
| Peak frequency (Hz) | 4772.52 ± 142.69 | 4588.59 ± 111.005 | 1.02 | 0.32 |
| Lowest frequency (Hz) | 2154.57 ± 109.06 | 1966.45 ± 110.92 | 1.21 | 0.24 |
| Response Variable | VP | ORW | |||||
|---|---|---|---|---|---|---|---|
| SE | Z | p | SE | Z | p | ||
| Begging duration, marginal pseudo-R2 = 0.636 (VP) and 0.52 (ORW) | |||||||
| Intercept | 0.05 | 73.03 | <0.001 | Intercept | 0.08 | 35.59 | <0.001 |
| Conspecific mobbing alarm calls | 0.07 | −7.67 | <0.001 | Conspecific mobbing alarm calls | 0.09 | −10.4 | <0.001 |
| Background noise | 0.06 | −0.16 | 0.83 | Background noise | 0.08 | −7.99 | 0.13 |
| Heterospecific mobbing alarm calls | 0.07 | −7.47 | <0.001 | Heterospecific mobbing alarm calls | 0.08 | −1.5 | <0.001 |
| Number of begging calls, marginal pseudo-R2 = 0.45 (VP) and 0.36 (ORW) | |||||||
| Intercept | 0.08 | 35.29 | <0.001 | Intercept | 0.12 | 24.69 | <0.001 |
| Conspecific mobbing alarm calls | 0.09 | −7.13 | <0.001 | Conspecific mobbing alarm calls | 0.1 | −9.03 | <0.001 |
| Background noise | 0.08 | −0.61 | 0.54 | Background noise | 0.08 | −6.3 | 0.67 |
| Heterospecific mobbing alarm calls | 0.09 | −7.79 | <0.001 | Heterospecific mobbing alarm calls | 0.09 | −0.42 | <0.001 |
| Beak opening frequency, marginal pseudo-R2 = 0.01 (VP) and 0.2 (ORW) | |||||||
| Intercept | 0.11 | 14.84 | <0.001 | Intercept | 0.01 | 18.62 | <0.001 |
| Conspecific mobbing alarm calls | 0.16 | −0.55 | 0.58 | Conspecific mobbing alarm calls | 0.15 | −2.67 | 0.01 |
| Background noise | 0.15 | −0.08 | 0.93 | Background noise | 0.14 | −1.43 | 0.15 |
| Heterospecific mobbing alarm calls | 0.16 | −0.31 | 0.76 | Heterospecific mobbing alarm calls | 0.14 | −0.84 | 0.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Y.; Han, J.; Yang, C. Anti-Predation Responses to Conspecific versus Heterospecific Alarm Calls by the Nestlings of Two Sympatric Birds. Animals 2022, 12, 2156. https://doi.org/10.3390/ani12162156
Jiang Y, Han J, Yang C. Anti-Predation Responses to Conspecific versus Heterospecific Alarm Calls by the Nestlings of Two Sympatric Birds. Animals. 2022; 12(16):2156. https://doi.org/10.3390/ani12162156
Chicago/Turabian StyleJiang, Yuxin, Jingru Han, and Canchao Yang. 2022. "Anti-Predation Responses to Conspecific versus Heterospecific Alarm Calls by the Nestlings of Two Sympatric Birds" Animals 12, no. 16: 2156. https://doi.org/10.3390/ani12162156