
Transcriptomic Analysis of Circulating Leukocytes Obtained during the Recovery from Clinical Mastitis Caused by Escherichia coli in Holstein Dairy Cows

 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Microbiological Analysis
2.3. In Vitro Blood Tests
2.4. Measurement of IL-1B and Nitric Oxide
2.5. RNA Extraction
2.6. RNA-Sequencing, Mapping and Quantification
2.7. Analysis of Differentially Expression between Groups
2.8. Enrichment, Pathway and Cluster Analysis
2.9. Variant Calling of Reads from RNA-Seq
2.10. Statistical Analysis
3. Results
3.1. Group Characteristics
3.2. In Vitro PBMC Responses
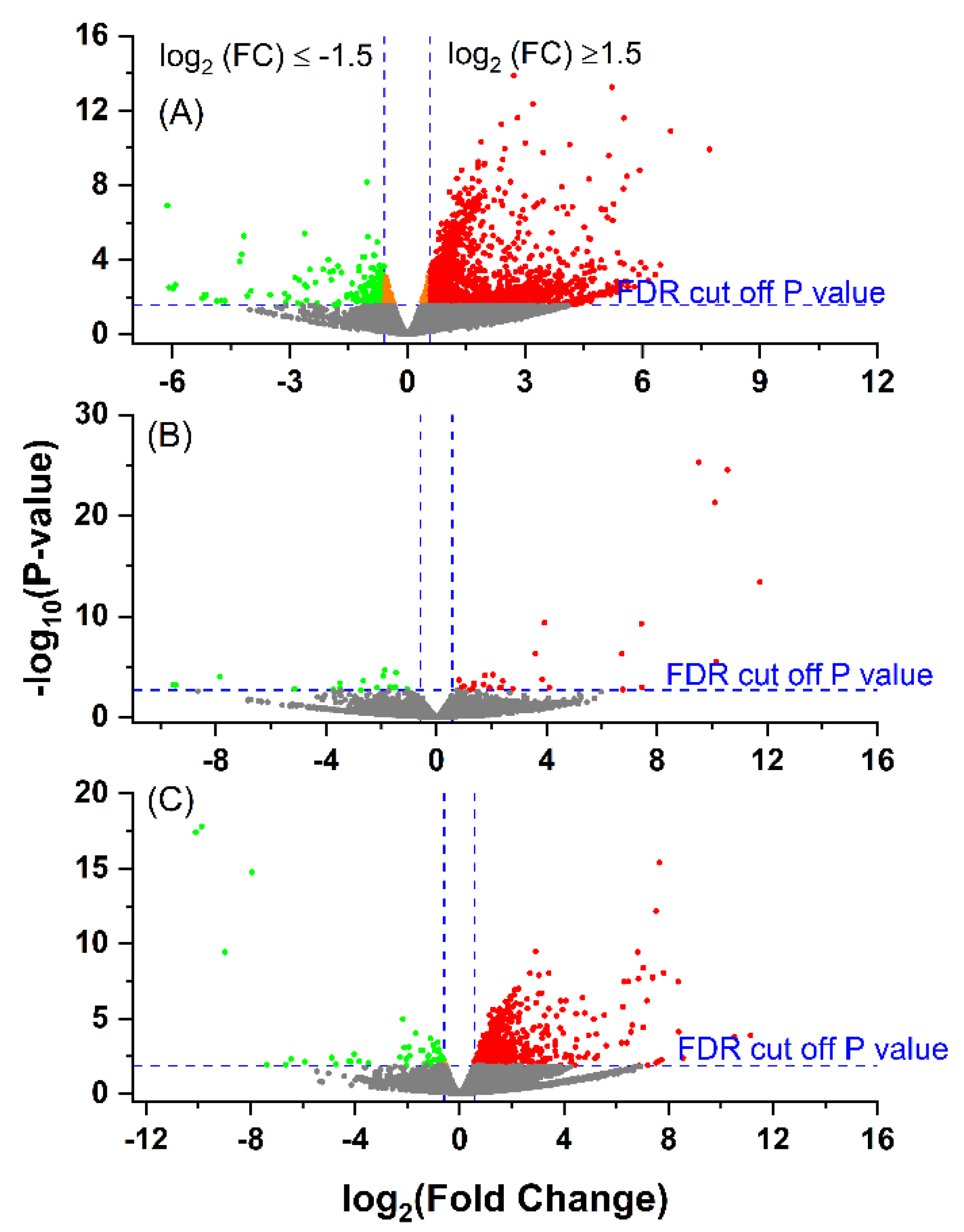
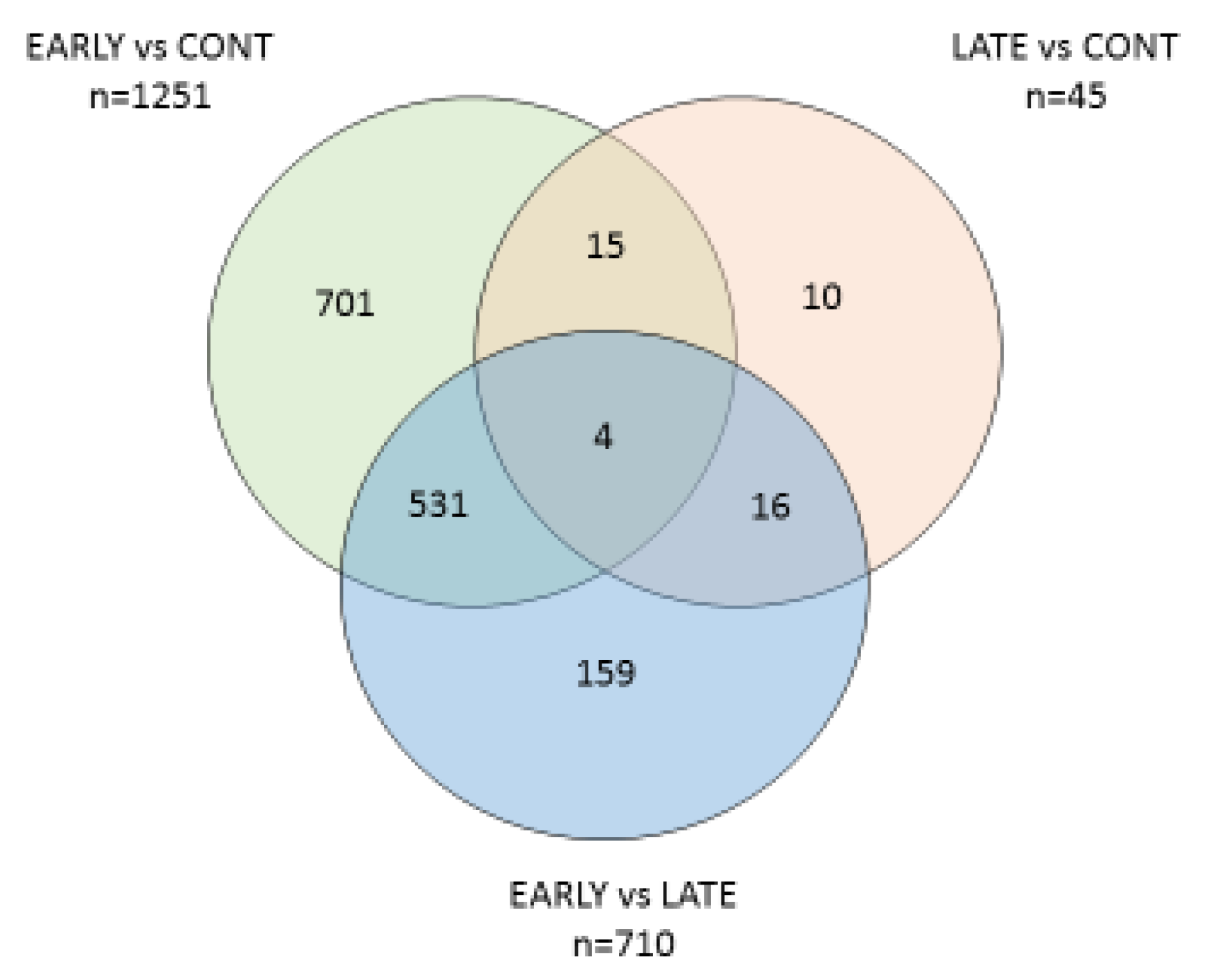
3.3. Differential Gene Expression between the Groups
3.4. Comparison between E. coli(+) (EARLY) and E. coli(−) (CONT) Cows
3.5. Comparison between E. coli(+) (LATE) and E. coli(-) (CONT) Cows
3.6. Comparison between E. coli(+) (EARLY) and E. coli(+) (LATE) Cows
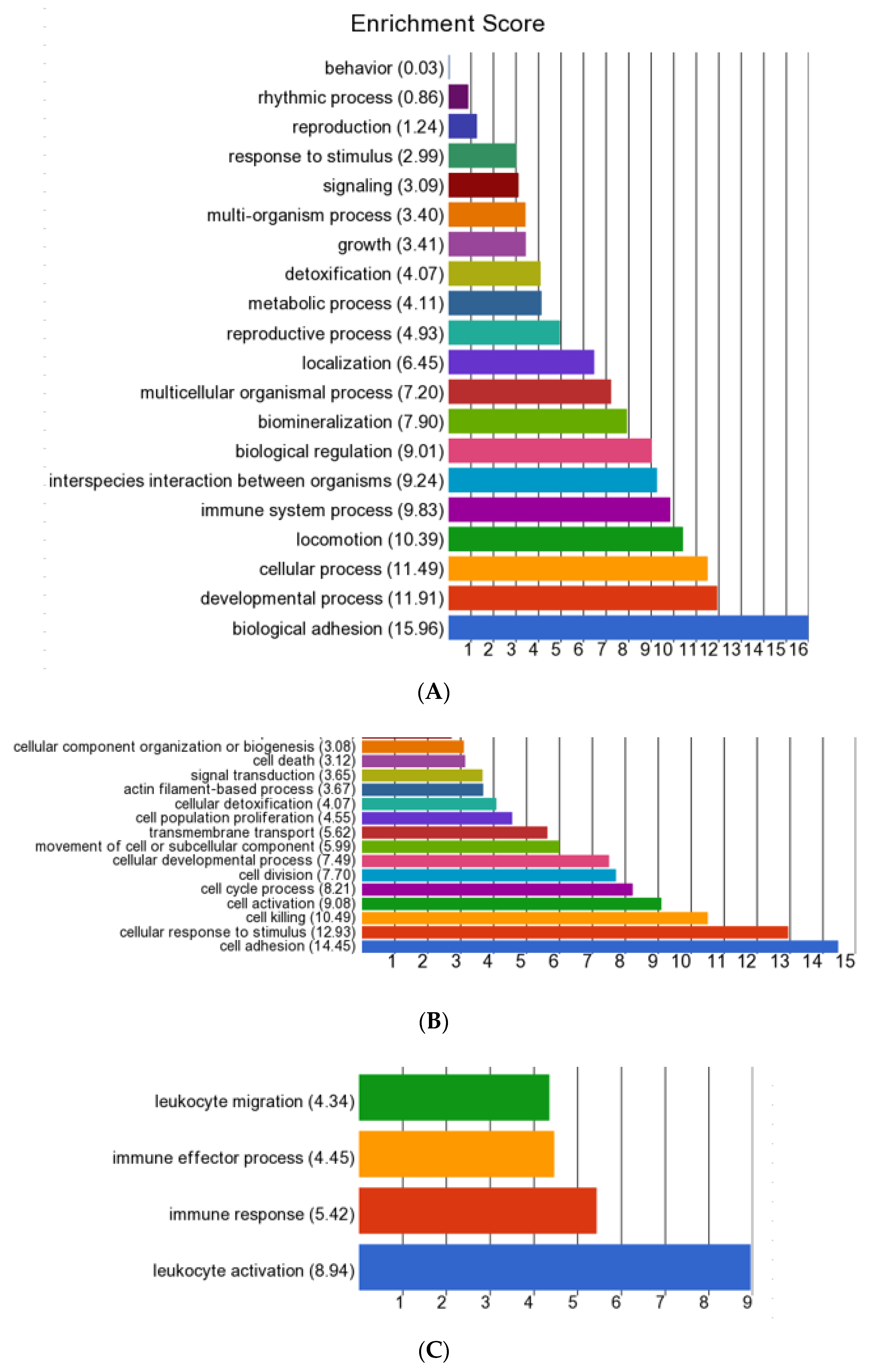
3.7. GO Enrichment and Cluster Analysis for Gene Functions
3.8. Variant Calling
4. Discussion
4.1. Evidence for an Ongoing Inflammatory Response during the Resolution Stage of an E. coli Infection
4.2. Evidence for Genetic Differences between E. coli Infected and Healthy Cows
4.3. Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aghamohammadi, M.; Haine, D.; Kelton, D.F.; Barkema, H.W.; Hogeveen, H.; Keefe, G.P.; Dufour, S. Herd-Level Mastitis-Associated Costs on Canadian Dairy Farms. Front. Vet. Sci. 2018, 5, 100. [Google Scholar] [CrossRef]
- Petzl, W.; Zerbe, H.; Gunther, J.; Seyfert, H.M.; Hussen, J.; Schuberth, H.J. Pathogen-specific responses in the bovine udder. Models and immunoprophylactic concepts. Res. Vet. Sci. 2018, 116, 55–61. [Google Scholar] [CrossRef]
- Harmon, R.J. Physiology of mastitis and factors affecting somatic cell counts. J. Dairy Sci. 1994, 77, 2103–2112. [Google Scholar] [CrossRef]
- Miles, A.M.; Huson, H.J. Graduate Student Literature Review: Understanding the genetic mechanisms underlying mastitis. J. Dairy Sci. 2021, 104, 1183–1191. [Google Scholar] [CrossRef]
- Bradley, A. Bovine mastitis: An evolving disease. Vet. J. 2002, 164, 116–128. [Google Scholar] [CrossRef]
- Burvenich, C.; Van Merris, V.; Mehrzad, J.; Diez-Fraile, A.; Duchateau, L. Severity of E. coli mastitis is mainly determined by cow factors. Vet. Res. 2003, 34, 521–564. [Google Scholar] [CrossRef] [Green Version]
- Dogan, B.; Klaessig, S.; Rishniw, M.; Almeida, R.A.; Oliver, S.P.; Simpson, K.; Schukken, Y.H. Adherent and invasive Escherichia coli are associated with persistent bovine mastitis. Vet. Microbiol. 2006, 116, 270–282. [Google Scholar] [CrossRef]
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell. Mol. Life Sci. 2021, 78, 1233–1261. [Google Scholar] [CrossRef]
- De Schepper, S.; De Ketelaere, A.; Bannerman, D.D.; Paape, M.J.; Peelman, L.; Burvenich, C. The toll-like receptor-4 (TLR-4) pathway and its possible role in the pathogenesis of Escherichia coli mastitis in dairy cattle. Vet. Res. 2008, 39, 5. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Schukken, Y.H.; Gunther, J.; Fitzpatrick, J.; Fontaine, M.C.; Goetze, L.; Holst, O.; Leigh, J.; Petzl, W.; Schuberth, H.J.; Sipka, A.; et al. Host-response patterns of intramammary infections in dairy cows. Vet. Immunol. Immunopathol. 2011, 144, 270–289. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, M.; Li, R.W.; Capuco, A.V. Mastitis associated transcriptomic disruptions in cattle. Vet. Immunol. Immunopathol. 2010, 138, 267–279. [Google Scholar] [CrossRef]
- Bruckmaier, R.M.; Wellnitz, O. TRIENNIAL LACTATION SYMPOSIUM/BOLFA: Pathogen-specific immune response and changes in the blood-milk barrier of the bovine mammary gland. J. Anim. Sci. 2017, 95, 5720–5728. [Google Scholar] [CrossRef]
- Blum, S.E.; Heller, E.D.; Jacoby, S.; Krifucks, O.; Leitner, G. Comparison of the immune responses associated with experimental bovine mastitis caused by different strains of Escherichia coli. J. Dairy Res. 2017, 84, 190–197. [Google Scholar] [CrossRef]
- Buitenhuis, B.; Rontved, C.M.; Edwards, S.M.; Ingvartsen, K.L.; Sorensen, P. In depth analysis of genes and pathways of the mammary gland involved in the pathogenesis of bovine Escherichia coli-mastitis. BMC Genom. 2011, 12, 130. [Google Scholar] [CrossRef] [Green Version]
- Loor, J.J.; Moyes, K.M.; Bionaz, M. Functional adaptations of the transcriptome to mastitis-causing pathogens: The mammary gland and beyond. J. Mammary Gland. Biol. Neoplasia 2011, 16, 305–322. [Google Scholar] [CrossRef]
- Lawless, N.; Reinhardt, T.A.; Bryan, K.; Baker, M.; Pesch, B.; Zimmerman, D.; Zuelke, K.; Sonstegard, T.; O’Farrelly, C.; Lippolis, J.D.; et al. MicroRNA regulation of bovine monocyte inflammatory and metabolic networks in an in vivo infection model. G3 Genes Genomes Genet. 2014, 4, 957–971. [Google Scholar] [CrossRef] [Green Version]
- Rainard, P.; Cunha, P.; Gilbert, F.B. Innate and Adaptive Immunity Synergize to Trigger Inflammation in the Mammary Gland. PLoS ONE 2016, 11, e0154172. [Google Scholar] [CrossRef]
- Gibson, A.J.; Woodman, S.; Pennelegion, C.; Patterson, R.; Stuart, E.; Hosker, N.; Siviter, P.; Douglas, C.; Whitehouse, J.; Wilkinson, W.; et al. Differential macrophage function in Brown Swiss and Holstein Friesian cattle. Vet. Immunol. Immunopathol. 2016, 181, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Genome Project Data Processing S. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- DePristo, M.A.; Banks, E.; Poplin, R.; Garimella, K.V.; Maguire, J.R.; Hartl, C.; Philippakis, A.A.; del Angel, G.; Rivas, M.A.; Hanna, M.; et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat. Genet. 2011, 43, 491–498. [Google Scholar] [CrossRef] [PubMed]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. Gigascience 2015, 4, 7. [Google Scholar] [CrossRef]
- Vaarst, M.; Enevoldsen, C. Patterns of clinical mastitis manifestations in Danish organic dairy herds. J. Dairy Res. 1997, 64, 23–37. [Google Scholar] [CrossRef]
- Warner, C.M.; Meeker, D.L.; Rothschild, M.F. Genetic control of immune responsiveness: A review of its use as a tool for selection for disease resistance. J. Anim. Sci. 1987, 64, 394–406. [Google Scholar] [CrossRef] [Green Version]
- White, L.J.; Schukken, Y.H.; Dogan, B.; Green, L.; Dopfer, D.; Chappell, M.J.; Medley, G.F. Modelling the dynamics of intramammary E. coli infections in dairy cows: Understanding mechanisms that distinguish transient from persistent infections. Vet. Res. 2010, 41, 13. [Google Scholar] [CrossRef] [Green Version]
- Blum, S.E.; Heller, D.E.; Jacoby, S.; Krifuks, O.; Merin, U.; Silanikove, N.; Lavon, Y.; Edery, N.; Leitner, G. Physiological response of mammary glands to Escherichia coli infection:A conflict between glucose need for milk production and immune response. Sci. Rep. 2020, 10, 9602. [Google Scholar] [CrossRef]
- Saxena, M.; Yeretssian, G. NOD-Like Receptors: Master Regulators of Inflammation and Cancer. Front. Immunol. 2014, 5, 327. [Google Scholar] [CrossRef] [Green Version]
- Roe, K.; Gibot, S.; Verma, S. Triggering receptor expressed on myeloid cells-1 (TREM-1): A new player in antiviral immunity? Front. Microbiol. 2014, 5, 627. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, F.B.; Cunha, P.; Jensen, K.; Glass, E.J.; Foucras, G.; Robert-Granie, C.; Rupp, R.; Rainard, P. Differential response of bovine mammary epithelial cells to Staphylococcus aureus or Escherichia coli agonists of the innate immune system. Vet. Res. 2013, 44, 40. [Google Scholar] [CrossRef] [Green Version]
- Younis, S.; Javed, Q.; Blumenberg, M. Meta-Analysis of Transcriptional Responses to Mastitis-Causing Escherichia coli. PLoS ONE 2016, 11, e0148562. [Google Scholar] [CrossRef] [Green Version]
- Afacan, N.J.; Yeung, A.T.; Pena, O.M.; Hancock, R.E. Therapeutic potential of host defense peptides in antibiotic-resistant infections. Curr. Pharm. Des. 2012, 18, 807–819. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; Buggiotti, L.; Salavati, M.; Marchitelli, C.; Palma-Vera, S.; Wylie, A.; Takeda, H.; Tang, L.; Crowe, M.A.; Wathes, D.C.; et al. Global transcriptomic profiles of circulating leucocytes in early lactation cows with clinical or subclinical mastitis. Mol. Biol. Rep. 2021, 48, 4611–4623. [Google Scholar] [CrossRef]
- Blyth, G.A.D.; Connors, L.; Fodor, C.; Cobo, E.R. The Network of Colonic Host Defense Peptides as an Innate Immune Defense Against Enteropathogenic Bacteria. Front. Immunol. 2020, 11, 965. [Google Scholar] [CrossRef]
- Donato, R. Intracellular and extracellular roles of S100 proteins. Microsc. Res. Tech. 2003, 60, 540–551. [Google Scholar] [CrossRef]
- Pyorala, S.; Hovinen, M.; Simojoki, H.; Fitzpatrick, J.; Eckersall, P.D.; Orro, T. Acute phase proteins in milk in naturally acquired bovine mastitis caused by different pathogens. Vet. Rec. 2011, 168, 535. [Google Scholar] [CrossRef]
- Kutila, T.; Pyorala, S.; Saloniemi, H.; Kaartinen, L. Antibacterial effect of bovine lactoferrin against udder pathogens. Acta Vet. Scand. 2003, 44, 35–42. [Google Scholar] [CrossRef]
- Ratledge, C.; Dover, L.G. Iron metabolism in pathogenic bacteria. Annu. Rev. Microbiol. 2000, 54, 881–941. [Google Scholar] [CrossRef]
- Thurm, C.; Schraven, B.; Kahlfuss, S. ABC Transporters in T Cell-Mediated Physiological and Pathological Immune Responses. Int. J. Mol. Sci. 2021, 22, 9186. [Google Scholar] [CrossRef]
- Wathes, D.C.; Becker, F.; Buggiotti, L.; Crowe, M.A.; Ferris, C.; Foldager, L.; Grelet, C.; Hostens, M.; Ingvartsen, K.L.; Marchitelli, C.; et al. Associations between Circulating IGF-1 Concentrations, Disease Status and the Leukocyte Transcriptome in Early Lactation Dairy Cows. Ruminants 2021, 1, 147–177. [Google Scholar] [CrossRef]
- Inami, A.; Kiyono, H.; Kurashima, Y. ATP as a Pathophysiologic Mediator of Bacteria-Host Crosstalk in the Gastrointestinal Tract. Int. J. Mol. Sci. 2018, 19, 2371. [Google Scholar] [CrossRef] [Green Version]
- Proietti, M.; Perruzza, L.; Scribano, D.; Pellegrini, G.; D’Antuono, R.; Strati, F.; Raffaelli, M.; Gonzalez, S.F.; Thelen, M.; Hardt, W.D.; et al. ATP released by intestinal bacteria limits the generation of protective IgA against enteropathogens. Nat. Commun. 2019, 10, 250. [Google Scholar] [CrossRef]
- Sun, Y.; Sheshadri, N.; Zong, W.X. SERPINB3 and B4: From biochemistry to biology. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA, 2017; Volume 62, pp. 170–177. [Google Scholar] [CrossRef] [Green Version]
- Yaron, J.R.; Zhang, L.; Guo, Q.; Haydel, S.E.; Lucas, A.R. Fibrinolytic Serine Proteases, Therapeutic Serpins and Inflammation: Fire Dancers and Firestorms. Front. Cardiovasc. Med. 2021, 8, 648947. [Google Scholar] [CrossRef]
- Greer, J.P.; Arber, D.A.; Glader, B.E.; List, A.F.; Means, R.M.; Rodgers, G.M. Wintrobe’s Clinical Hematology; Lippincott, Williams and Wilkins: Philadelphia, PA, USA, 2013. [Google Scholar]
- Lee, E.; Choi, H.S.; Hwang, J.H.; Hoh, J.K.; Cho, Y.H.; Baek, E.J. The RNA in reticulocytes is not just debris: It is necessary for the final stages of erythrocyte formation. Blood Cells Mol. Dis. 2014, 53, 1–10. [Google Scholar] [CrossRef]
- Isobe, N. Control mechanisms for producing antimicrobial factors in ruminant mammary gland. Anim. Sci. J. 2017, 88, 937–943. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Ruiz, S.R.; Torres-Aguilar, H.; Gonzalez-Dominguez, E.; Narvaez, J.; Gonzalez-Perez, G.; Vargas-Ayala, G.; Meraz-Rios, M.A.; Garcia-Zepeda, E.A.; Sanchez-Torres, C. Human CD16+ and CD16- monocyte subsets display unique effector properties in inflammatory conditions in vivo. J. Leukoc. Biol. 2011, 90, 1119–1131. [Google Scholar] [CrossRef]
- Hussen, J.; Duvel, A.; Sandra, O.; Smith, D.; Sheldon, I.M.; Zieger, P.; Schuberth, H.J. Phenotypic and functional heterogeneity of bovine blood monocytes. PLoS ONE 2013, 8, e71502. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Pamer, E.G. Monocyte recruitment during infection and inflammation. Nat. Rev. Immunol. 2011, 11, 762–774. [Google Scholar] [CrossRef] [Green Version]
- Bokoch, G.M.; Knaus, U.G. NADPH oxidases: Not just for leukocytes anymore! Trends Biochem. Sci. 2003, 28, 502–508. [Google Scholar] [CrossRef]
- Quinn, M.T.; Gauss, K.A. Structure and regulation of the neutrophil respiratory burst oxidase: Comparison with nonphagocyte oxidases. J. Leukoc. Biol. 2004, 76, 760–781. [Google Scholar] [CrossRef]
- Jungi, T.W.; Adler, H.; Adler, B.; Thony, M.; Krampe, M.; Peterhans, E. Inducible nitric oxide synthase of macrophages. Present knowledge and evidence for species-specific regulation. Vet. Immunol. Immunopathol. 1996, 54, 323–330. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef]
- Forman, H.J.; Torres, M. Redox signaling in macrophages. Mol. Aspects Med. 2001, 22, 189–216. [Google Scholar] [CrossRef]
- Liu, D.; Chen, Z.; Zhao, W.; Guo, L.; Sun, H.; Zhu, K.; Liu, G.; Shen, X.; Zhao, X.; Wang, Q.; et al. Genome-wide selection signatures detection in Shanghai Holstein cattle population identified genes related to adaption, health and reproduction traits. BMC Genom. 2021, 22, 747. [Google Scholar] [CrossRef]
- Akinduro, O.; Sully, K.; Patel, A.; Robinson, D.J.; Chikh, A.; McPhail, G.; Braun, K.M.; Philpott, M.P.; Harwood, C.A.; Byrne, C.; et al. Constitutive Autophagy and Nucleophagy during Epidermal Differentiation. J. Investig. Dermatol. 2016, 136, 1460–1470. [Google Scholar] [CrossRef] [Green Version]
- Hugo, S.E.; Cruz-Garcia, L.; Karanth, S.; Anderson, R.M.; Stainier, D.Y.; Schlegel, A. A monocarboxylate transporter required for hepatocyte secretion of ketone bodies during fasting. Genes Dev. 2012, 26, 282–293. [Google Scholar] [CrossRef] [Green Version]
- Lougaris, V.; Baronio, M.; Castagna, A.; Tessarin, G.; Rossi, S.; Gazzurelli, L.; Benvenuto, A.; Moratto, D.; Chiarini, M.; Cattalini, M.; et al. Paediatric MAS/HLH caused by a novel monoallelic activating mutation in p110delta. Clin. Immunol. 2020, 219, 108543. [Google Scholar] [CrossRef]
- Valtcheva, N.; Primorac, A.; Jurisic, G.; Hollmen, M.; Detmar, M. The orphan adhesion G protein-coupled receptor GPR97 regulates migration of lymphatic endothelial cells via the small GTPases RhoA and Cdc42. J. Biol. Chem. 2013, 288, 35736–35748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kour, A.; Deb, S.M.; Nayee, N.; Raina, V.S.; Yadav, V.; Niranjan, S.K. Understanding the genomic architecture of clinical mastitis in Bos indicus. 3 Biotech 2021, 11, 466. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Zhong, R.; Qiu, C.; Zou, B.B. The Prognostic Value of GNG7 in Colorectal Cancer and Its Relationship With Immune Infiltration. Front. Genet. 2022, 13, 833013. [Google Scholar] [CrossRef] [PubMed]
- Evstafieva, A.G.; Kovaleva, I.E.; Shoshinova, M.S.; Budanov, A.V.; Chumakov, P.M. Implication of KRT16, FAM129A and HKDC1 genes as ATF4 regulated components of the integrated stress response. PLoS ONE 2018, 13, e0191107. [Google Scholar] [CrossRef] [Green Version]
- Telang, S.; Clem, B.F.; Klarer, A.C.; Clem, A.L.; Trent, J.O.; Bucala, R.; Chesney, J. Small molecule inhibition of 6-phosphofructo-2-kinase suppresses t cell activation. J. Transl. Med. 2012, 10, 95. [Google Scholar] [CrossRef] [Green Version]
- Ramezani-Rad, P.; Chen, C.; Zhu, Z.; Rickert, R.C. Cyclin D3 Governs Clonal Expansion of Dark Zone Germinal Center B Cells. Cell. Rep. 2020, 33, 108403. [Google Scholar] [CrossRef]
- Hinton, T.V.; Batelu, S.; Gleason, N.; Stemmler, T.L. Molecular characteristics of proteins within the mitochondrial Fe-S cluster assembly complex. Micron 2022, 153, 103181. [Google Scholar] [CrossRef]
- Rupp, R.; Boichard, D. Genetics of resistance to mastitis in dairy cattle. Vet. Res. 2003, 34, 671–688. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Sonstegard, T.S.; Cole, J.B.; VanTassell, C.P.; Wiggans, G.R.; Crooker, B.A.; Tan, C.; Prakapenka, D.; Liu, G.E.; Da, Y. Genome changes due to artificial selection in U.S. Holstein cattle. BMC Genom. 2019, 20, 128. [Google Scholar] [CrossRef]
- Brown, D.; Trowsdale, J.; Allen, R. The LILR family: Modulators of innate and adaptive immune pathways in health and disease. Tissue Antigens 2004, 64, 215–225. [Google Scholar] [CrossRef]
- Tsuchiya, H.; Shiota, G. Immune evasion by cancer stem cells. Regen. Ther. 2021, 17, 20–33. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| E. coli(−) * CONT | E. coli(+) EARLY ‡ | E. coli(+) LATE ‡ | |
|---|---|---|---|
| n | 6 | 6 | 3 |
| Control NO production (µM) | 8.7 ± 0.37 | 10.3 ± 1.72 | 8.6 ± 0.06 |
| LPS stimulated NO production (µM) | 8.5 ± 0.18 # | 8.5 ± 0.13 | 9.4 ± 0.56 # |
| Control IL1B production (pg/mL) | 87.0 ± 18.58 a | 59.8 ±11.87 a | 61.3 ± 14.83 a |
| LPS stimulated IL1B production (pg/mL) | 258.3 ± 86.90 b | 195.5 ± 40.08 b | 205.7 ± 38.84 b |
| Category | DEGs |
|---|---|
| NOD-like receptor signalling pathway | BCL2L1, CAMP, CATHL1, CATHL2, CATHL3, CATHL4, CATHL5, CATHL6, CXCL2, DEFB1, DEFB10, GABARAP, GABARAPL1, IFNAR1, IFNAR2, IL1B, IL18, LOC100301263, LOC112441458, LOC112443481, MAP1LC3A, MAPK13, MAPK14, MAPK3, MEFV, MYD88, NAIP, NFKBIA, NLRX1, PLCB1, STAT2, TXNIP |
| Amoebobiasis | ACTN1, ARG1, ARG2, CASP3, COL1A1, COL1A2, COL3A1, COL4A2, COL4A2, CXCL2, FN1, IL1B, IL1R1, IL1R2, IL12B, LAMB1, LAMC1, LAMC2, LOC505658, LOC511106, LOC786348, PIK3CD, PLCB1, RAB5C, SERPINB4 |
| Chemokine signalling pathway | ARRB2, CCL16, CCR1, CCR6, CXCL13, CXCL2, CXCR1, CXCR2, CXCR4, FGR, FOXO3, GNG2, GNG7, JAK2, LOC100297044, MAPK3, NCF1, NFKBIA, PAK1, PARD3, PIK3CD, PIK3CG, PLCB1, PREX1, PTK2B, PXN, RAC2, STAT2, STAT3, TIAM1, XCR1 |
| Calcium | ACTN1, ADGRE5, ALOX15, ALPL, ANXA1, ANXA9, CAPN1, CAPN3, CDH13, COL1A1, COL1A2, CPNE2, DYSF, EHD1, ENTPD1, F5, FBN1, FGG, ITGA3, LOXL2, MMP2, MMP8, MMP9, NOTCH2, PADI3, PADI4, PLA2G4A, PLA2G4F, PLCB1, PLCD1, PRSS2, PVALB, RASGRP4, RELN, RPH3A, RYR1, S100A12, S100A8, S100A9, SELL, SLC24A3, SPARC, SVIL, TGM1, TGM2, TGM3, TKT, TRPC5, TRPC6, TYROBP |
| Interleukin receptors | IL1R1, IL1R2, IL1RAP, IL1RL1, IL18R1, IL18RAP, MYD88, TGM2 |
| Cathelicidins and other antimicrobials | CAMP, CATHL1, CATHL2, CATHL3, CATHL4, CATHL5, CATHL6, CHI3L1, COL1A1, COL1A2, CXCL13, CXCL2, DEFB1, DEFB10, DEFB4A, DEFB7, DPT, FN1, HP, LTF, PGLYRP1, PGLYRP4, PTAFR, S100A8, S100A9, S100A12, SCARB1 |
| Peptide cross-linking | ANXA1, COL3A1, DSP, EPB42, FN1, TGM1, TGM2, TGM3 |
| Wound healing | ALOX15, AQP1, CNN2, COL3A1, DSP, FN1, NOTCH2, PAK1, PARD, PTK7, SDC1, SLC11A1, YAP1 |
| Protease binding | A2M, ANXA9, ATP9A, CDK5R1, COL1A1, COL1A2, COL3A1, ELANE, FLOT1, FN1, IL1R1, ITGA3, LOC506828, SELL |
| Antifolate resistance and ABC transporters | ABCA6, ABCA7, ABCB11, ABCA13, FOLR3, IL1B, LOC509854, LOC520016, LOC522174, LOC100337053, LOC100847574, LOC107131218, LOC107131247, LOC107131259, LOC107131271, LOC107131273 |
| Collagen | COL1A1, COL1A2, COL3A1, COL4A2, COL6A1, COL6A2, COL6A3, CTHRC1, MMP2, PCOLCE2, PLOD3 |
| Gene Symbol | Max Group Mean | Fold Change | p (BH) | Group # | BTA | Gene Position |
|---|---|---|---|---|---|---|
| EYA3 | 2.616 | 1.511 | 0.025 | A | 2 | NC_037329.1 (125201889..125390272) |
| RAC2 | 6.971 | 1.871 | 0.000 | A | 5 | NC_037332.1 (75656456..75673385) |
| GNG7 | 3.301 | 1.875 | 0.001 | A | 7 | NC_037334.1 (21002517..21097631) |
| ARHGAP26 | 4.428 | 1.539 | 0.005 | B | 7 | NC_037334.1 (53811496..54284856) |
| EBF1 * | 2.895 | −1.563 | 0.017 | A/B | 7 | NC_037334.1 (70284253..70694732 |
| FAM129A | 31.780 | 2.061 | 0.000 | A | 16 | NC_037343.1 (65828044..66012042, |
| WIPI1 | 2.243 | 3.271 | 0.000 | A | 19 | NC_037346.1 (61752501..61782685) |
| ARSG * | 1.651 | 1.929 | 0.008 | A/B | 19 | NC_037346.1 (61781773..61886526) |
| SLC16A6 | 19.480 | 1.671 | 0.010 | B | 19 | NC_037346.1 (61867803..61877805) |
| PFKFB4 | 9.899 | 1.883 | 0.001 | B | 22 | NC_037349.1 (51321977..51363429) |
| BOLA-DOA | 23.840 | −1.539 | 0.006 | A | 23 | NC_037350.1 (7314757..7323452) |
| CCND3 | 29.144 | 1.594 | 0.014 | B | 23 | NC_037350.1 (15698650..15793268 |
| Category | Term | Genes | Fold Enrichment | p-Value | FDR |
|---|---|---|---|---|---|
| KEGG_PATHWAY | bta04310:Wnt signalling pathway | GSK3B, TCF7L2, CAMK2D, PPP3R1, CCND3, RAC2 | 8.06 | 0.0007 | 0.113 |
| UP_KEYWORDS | 4Fe-4S | DPYD, ACO2, NARFL | 34.25 | 0.0033 | 0.320 |
| KEGG_PATHWAY | bta05200:Pathways in cancer | GSK3B, TCF7L2, DAPK1, GNG7, RASSF5, TPR, RAC2, AKT1 | 3.70 | 0.0044 | 0.230 |
| KEGG_PATHWAY | bta05210:Colorectal cancer | GSK3B, TCF7L2, RAC2, AKT1 | 11.16 | 0.0050 | 0.230 |
| KEGG_PATHWAY | bta04662:B cell receptor signalling pathway | GSK3B, PPP3R1, RAC2, AKT1 | 10.52 | 0.0059 | 0.230 |
| GOTERM_CC_DIRECT | GO:0005925~focal adhesion | CCND3, PPP1R12A, RPLP0, RDX, RAC2, MYH9 | 4.94 | 0.0067 | 0.391 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Z.; Palma-Vera, S.; Buggiotti, L.; Salavati, M.; Becker, F.; Werling, D.; Wathes, D.C.; GplusE Consortium. Transcriptomic Analysis of Circulating Leukocytes Obtained during the Recovery from Clinical Mastitis Caused by Escherichia coli in Holstein Dairy Cows. Animals 2022, 12, 2146. https://doi.org/10.3390/ani12162146
Cheng Z, Palma-Vera S, Buggiotti L, Salavati M, Becker F, Werling D, Wathes DC, GplusE Consortium. Transcriptomic Analysis of Circulating Leukocytes Obtained during the Recovery from Clinical Mastitis Caused by Escherichia coli in Holstein Dairy Cows. Animals. 2022; 12(16):2146. https://doi.org/10.3390/ani12162146
Chicago/Turabian StyleCheng, Zhangrui, Sergio Palma-Vera, Laura Buggiotti, Mazdak Salavati, Frank Becker, Dirk Werling, D. Claire Wathes, and GplusE Consortium. 2022. "Transcriptomic Analysis of Circulating Leukocytes Obtained during the Recovery from Clinical Mastitis Caused by Escherichia coli in Holstein Dairy Cows" Animals 12, no. 16: 2146. https://doi.org/10.3390/ani12162146