
Identification and Characterization of Copy Number Variations Regions in West African Taurine Cattle

 ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
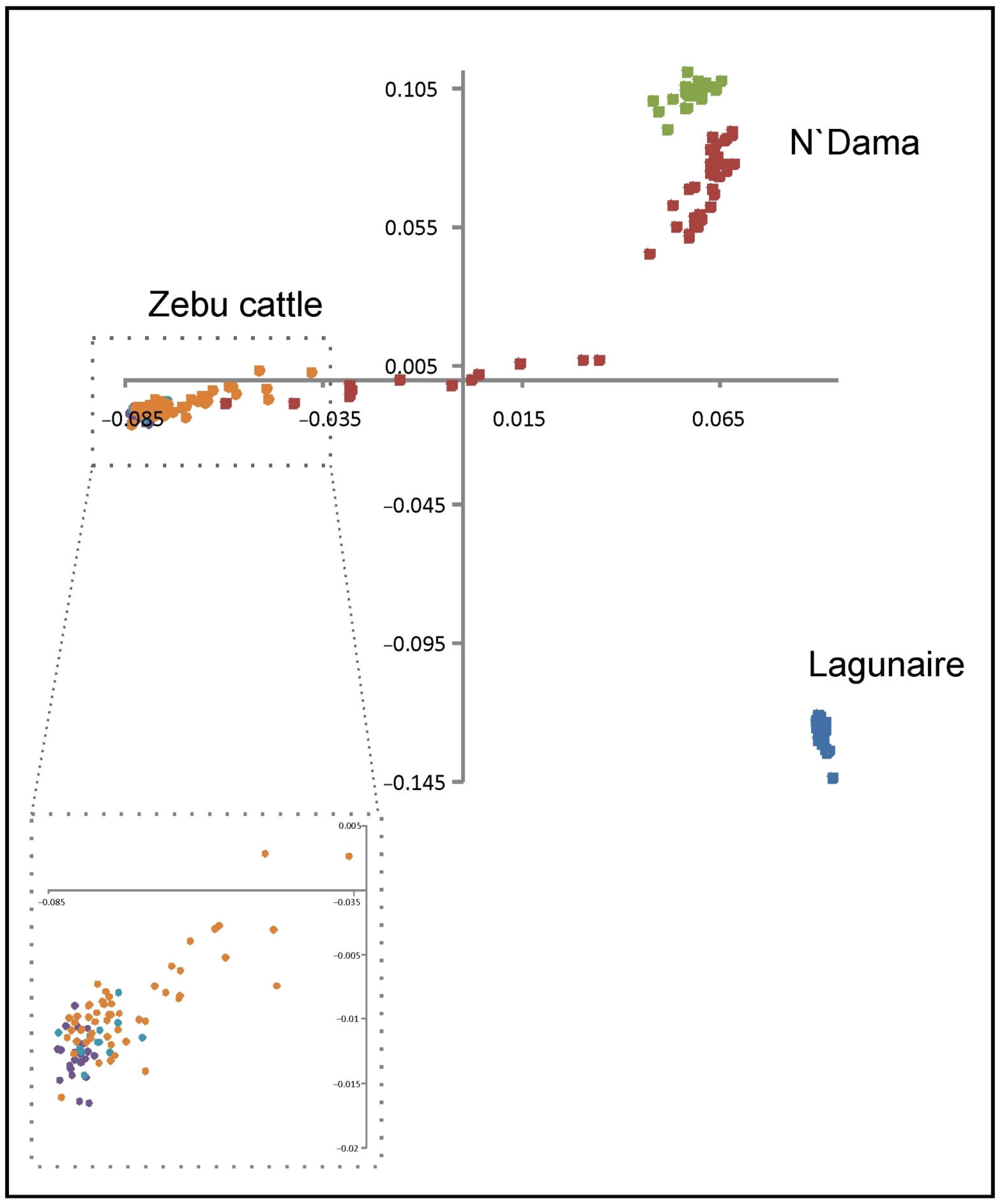
2.1. Samples, Genotyping, and Population Structure Analysis
2.2. CNV Calling, Editing, and Candidate CNVRs
2.3. Enrichment and Functional Annotation Analyses
2.4. Comparison with Previously Mapped QTL
3. Results
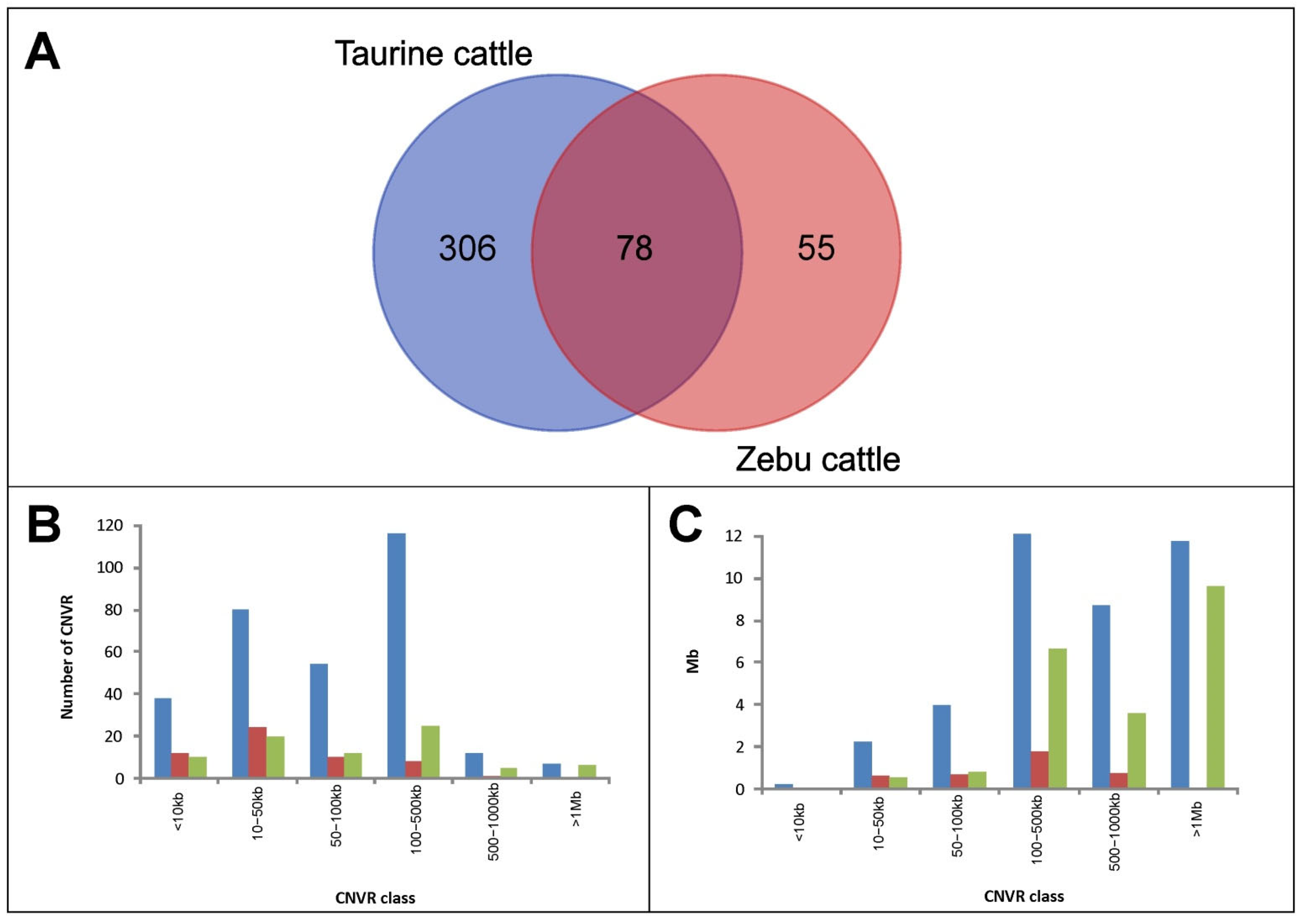
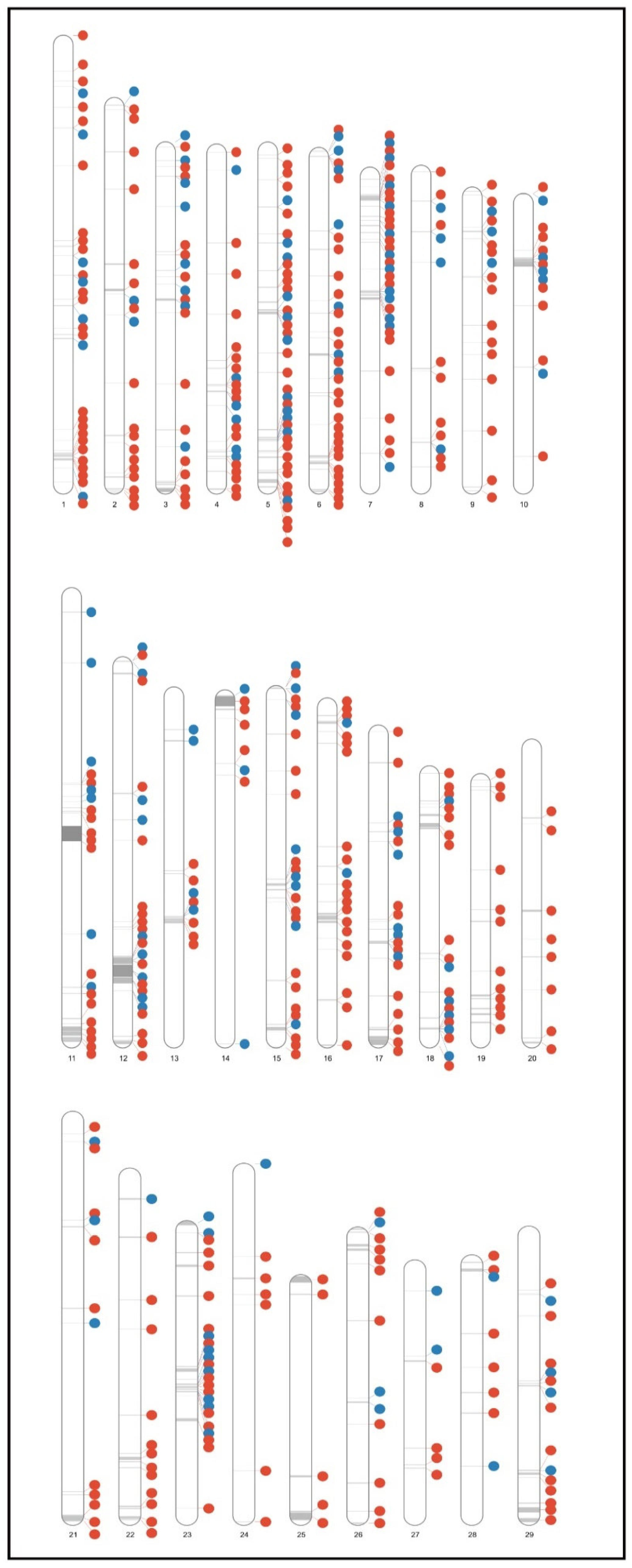
3.1. Construction of Candidate CNV Regions
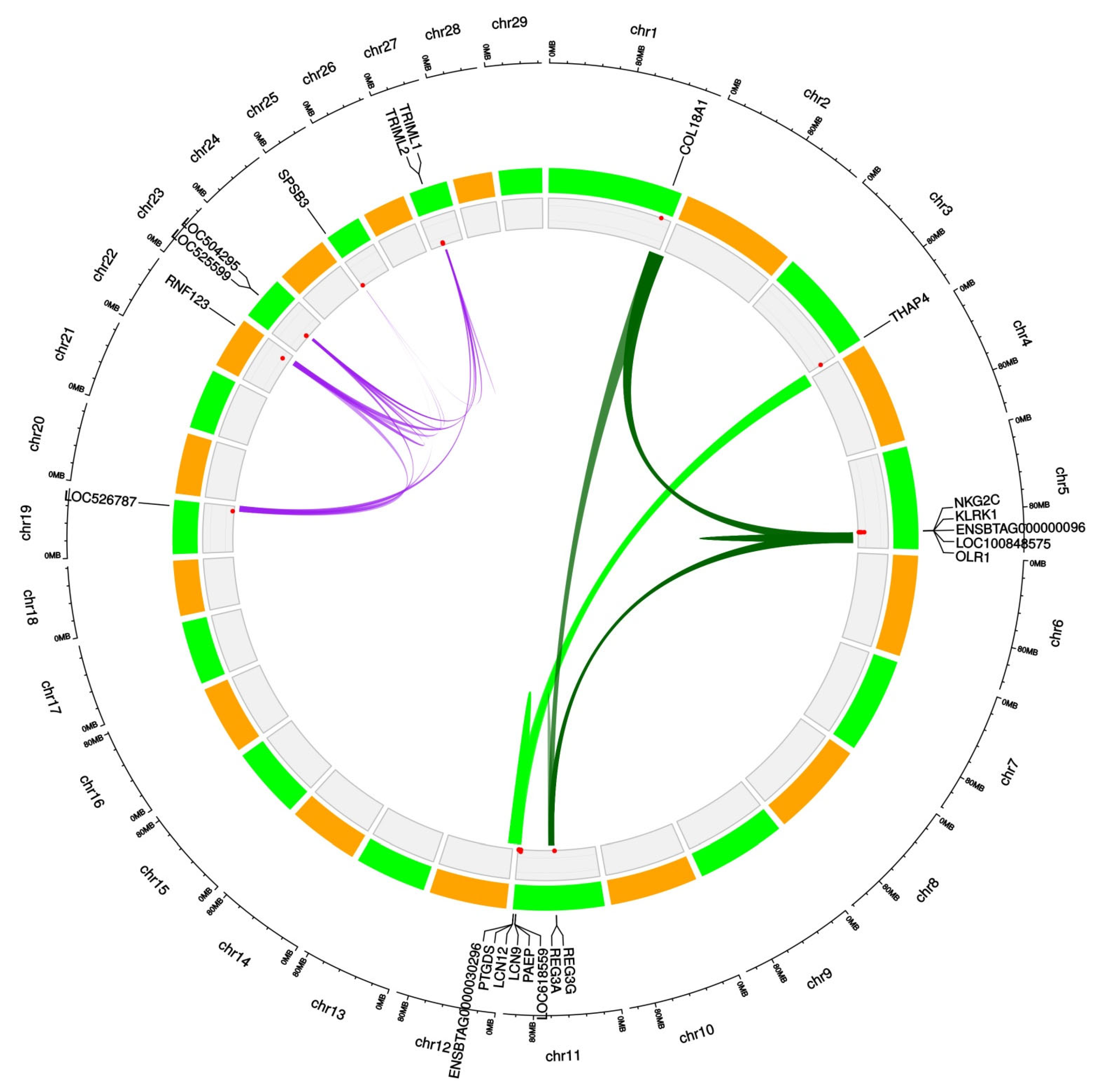
3.2. Enrichment and Functional Annotation Analyses
3.3. Correspondence with Trypanotolerance-Related QTL
4. Discussion
4.1. Identification of CNV Regions
4.2. Biological Importance of the CNVRs Identified in Taurine Cattle
4.3. Relationship between Trypanotolerance-Related QTL
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gifford-Gonzalez, D. Animal Disease Challenges to the Emergence of Pastoralism in Sub-Saharan Africa. Afr. Archaeol. Rev. 2000, 17, 45. [Google Scholar] [CrossRef]
- Flori, L.; Thevenon, S.; Dayo, G.K.; Senou, M.; Sylla, S.; Berthier, D.; Moazami-Goudarzi, K.; Gautier, M. Adaptive Admixture in the West African Bovine Hybrid Zone: Insight from the Borgou Population. Mol. Ecol. 2014, 23, 3241–3257. [Google Scholar] [CrossRef] [PubMed]
- Mwai, O.; Hanotte, O.; Kwon, Y.-J.; Cho, S. African Indigenous Cattle: Unique Genetic Resources in a Rapidly Changing World. Asian-Australas. J. Anim. Sci. 2015, 28, 911–921. [Google Scholar] [CrossRef] [PubMed]
- Geerts, S.; Osaer, S.; Goossens, B.; Faye, D. Trypanotolerance in Small Ruminants of Sub-Saharan Africa. Trends Parasitol. 2009, 25, 132–138. [Google Scholar] [CrossRef]
- Soudré, A.; Ouédraogo-Koné, S.; Wurzinger, M.; Müller, S.; Hanotte, O.; Ouédraogo, A.G.; Sölkner, J. Trypanosomosis: A Priority Disease in Tsetse-Challenged Areas of Burkina Faso. Trop. Anim. Health Prod. 2013, 45, 497–503. [Google Scholar] [CrossRef]
- Mattioli, R.C.; Pandey, V.S.; Murray, M.; Fitzpatrick, J.L. Immunogenetic Influences on Tick Resistance in African Cattle with Particular Reference to Trypanotolerant N’Dama (Bos Taurus) and Trypanosusceptible Gobra Zebu (Bos Indicus) Cattle. Acta Trop. 2000, 75, 263–277. [Google Scholar] [CrossRef]
- Berthier, D.; Peylhard, M.; Dayo, G.K.; Flori, G.K.; Sylla, S.; Bolly, S.; Sakande, H.; Chantal, I.; Thevenon, S. A Comparison of Phenotypic Traits Related to Trypanotolerance in Five West African Cattle Breeds Highlights the Value of Shorthorn Taurine Breeds. PLoS ONE 2015, 10, e0126498. [Google Scholar] [CrossRef]
- Orenge, C.O.; Munga, L.; Kimwele, C.N.; Kemp, S.; Korol, A.; Gibson, J.P.; Hanotte, O.; Soller, M. Trypanotolerance in N’Dama x Boran Crosses under Natural Trypanosome Challenge: Effect of Test-Year Environment, Gender, and Breed Composition. BMC Genet. 2012, 13, 87. [Google Scholar] [CrossRef]
- Dayo, G.K.; Gautier, M.; Berthier, D.; Poivey, J.P.; Sidibe, I.; Bengaly, Z.; Eggen, A.; Boichard, D.; Thevenon, S. Association Studies in QTL Regions Linked to Bovine Trypanotolerance in a West African Crossbred Population. Anim. Genet. 2011, 43, 123–132. [Google Scholar] [CrossRef]
- Hanotte, O.; Ronin, Y.; Agaba, M.; Nilsson, P.; Gelhaus, A.; Horstmann, R.; Sugimoto, Y.; Kemp, S.; Gibson, J.; Korol, A.; et al. Mapping of Quantitative Trait Loci Controlling Trypanotolerance in a Cross of Tolerant West African N’Dama and Susceptible East African Boran Cattle. Proc. Natl. Acad. Sci. USA 2003, 100, 7443–7448. [Google Scholar] [CrossRef]
- Álvarez, I.; Pérez-Pardal, L.; Traoré, A.; Fernández, I.; Goyache, F. African Cattle Do not Carry Unique Mutations on the Exon 9 of the ARHGAP15 Gene. Anim. Biotechnol. 2016, 27, 9–12. [Google Scholar] [CrossRef]
- Álvarez, I.; Pérez-Pardal, L.; Traoré, A.; Fernández, I.; Goyache, F. Lack of Haplotype Structuring for Two Candidate Genes for Trypanotolerance in Cattle. J. Anim. Breed. Genet. 2016, 133, 105–114. [Google Scholar] [CrossRef]
- Álvarez, I.; Pérez-Pardal, L.; Traoré, A.; Fernández, I.; Goyache, F. Lack of Specific Alleles for the Bovine Chemokine (C-X-C) Receptor Type 4 (CXCR4) Gene in West African Cattle Questions Its Role as a Candidate for Trypanotolerance. Infect. Genet. Evol. 2016, 42, 30–33. [Google Scholar] [CrossRef]
- Dayo, G.K.; Thevenon, S.; Berthier, D.; Moazami-Goudarzi, K.; Denis, C.; Cuny, G.; Eggen, A.; Gautier, M. Detection of Selection Signatures within Candidate Regions Underlying Trypanotolerance in Outbred Cattle Populations. Mol. Ecol. 2009, 18, 1801–1813. [Google Scholar] [CrossRef]
- Goyache, F.; Pérez-Pardal, L.; Fernández, I.; Traoré, A.; Menéndez-Arias, N.A.; Álvarez, I. Ancient Autozygous Segments Subject to Positive Selection Suggest Adaptive Immune Responses in West African Cattle. Gene 2021, 803, 145899. [Google Scholar] [CrossRef]
- Noyes, H.; Brass, A.; Obara, I.; Anderson, S.; Archibald, A.L.; Bradley, D.G.; Fisher, P.; Freeman, A.; Gibson, J.; Gicheru, M.; et al. Genetic and Expression Analysis of Cattle Identifies Candidate Genes in Pathways Responding to Trypanosoma Congolense Infection. Proc. Natl. Acad. Sci. USA 2011, 108, 9304–9309. [Google Scholar] [CrossRef]
- Feuk, L.; Carson, A.R.; Scherer, S.W. Structural Variation in the Human Genome. Nat. Rev. Genet. 2006, 7, 85–97. [Google Scholar] [CrossRef]
- Scherer, S.W.; Lee, C.; Birney, E.; Altshuler, D.M.; Eichler, E.E.; Carter, N.P.; Hurles, M.E.; Feuk, L. Challenges and Standards in Integrating Surveys of Structural Variation. Nat. Genet. 2007, 39, S7–S15. [Google Scholar] [CrossRef]
- Conrad, D.F.; Pinto, D.; Redon, R.; Feuk, L.; Gokcumen, O.; Zhang, Y.; Aerts, J.; Andrews, T.D.; Barnes, C.; Campbell, P.; et al. Origins and Functional Impact of Copy Number Variation in the Human Genome. Nature 2010, 464, 704–712. [Google Scholar] [CrossRef]
- Yang, L.; Niu, Q.; Zhang, T.; Zhao, G.; Zhu, B.; Chen, Y.; Zhang, L.; Gao, X.; Gao, H.; Liu, G.E.; et al. Genomic Sequencing Analysis Reveals Copy Number Variations and Their Associations with Economically Important Traits in Beef Cattle. Genomics 2021, 113, 812–820. [Google Scholar] [CrossRef]
- Da Silva, J.M.; Giachetto, P.F.; da Silva, L.O.; Cintra, L.C.; Paiva, S.R.; Yamagishi, M.E.B.; Caetano, A.R. Genome-Wide Copy Number Variation (CNV) Detection in Nelore Cattle Reveals Highly Frequent Variants in Genome Regions Harboring QTLs Affecting Production Traits. BMC Genom. 2016, 17, 454. [Google Scholar] [CrossRef]
- Traoré, A.; Koudandé, D.O.; Fernández, I.; Soudré, A.; Granda, V.; Álvarez, I.; Diarra, S.; Diarra, F.; Kaboré, A.; Sanou, M.; et al. Geographical Assessment of Body Measurements and Qualitative Traits in West African Cattle. Trop. Anim. Health Prod. 2015, 47, 1505–1513. [Google Scholar] [CrossRef]
- Traoré, A.; Koudandé, D.O.; Fernández, I.; Soudré, A.; Álvarez, I.; Diarra, S.; Diarra, F.; Kaboré, A.; Sanou, M.; Tamboura, H.H.; et al. Multivariate Characterization of Morphological Traits in West African Cattlesires. Arch. Anim. Breed. 2016, 59, 337–344. [Google Scholar] [CrossRef]
- Moussa, M.M.A.; Issa, M.; Traoré, A.; Grema, M.; Hamani, M.; Fernández, I.; Soudré, A.; Álvarez, I.; Sanou, M.; Tamboura, H.H.; et al. Morphological Assessment of the Zebu Bororo (Wodaabé) Cattle of Niger in the West African Zebu Framework. Arch. Anim. Breed. 2017, 60, 363–371. [Google Scholar] [CrossRef]
- Álvarez, I.; Traoré, A.; Fernández, I.; Cuervo, M.; Lecomte, T.; Soudré, A.; Kaboré, A.; Tamboura, H.H.; Goyache, F. Assessing Introgression of Sahelian Zebu Genes into Native Bos Taurus Breeds in Burkina Faso. Mol. Biol. Rep. 2014, 41, 3745–3754. [Google Scholar] [CrossRef]
- Coordinate Remapping Service: NCBI. Available online: https://www.ncbi.nlm.nih.gov/genome/tools/remap (accessed on 21 June 2022).
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-Generation PLINK: Rising to the Challenge of Larger and Richer Datasets. GigaScience 2015, 4, 7. [Google Scholar] [CrossRef]
- Wickham, H. Ggplo2: Elegant Data Visualisations Using the Grammar of Graphics. Available online: https://ggplot2.tidyverse.org/ (accessed on 20 June 2022).
- The Comprehensive R Archive Network. Available online: https://cran.r-project.org/ (accessed on 21 June 2022).
- Goyache, F.; Fernández, I.; Tapsoba, A.S.R.; Traoré, A.; Menéndez-Arias, N.A.; Álvarez, I. Functional Characterization of Copy Number Variations Regions in Djallonké Sheep. J. Anim. Breed. Genet. 2021, 138, 600–612. [Google Scholar] [CrossRef]
- Wang, K.; Li, M.; Hadley, D.; Liu, R.; Glessner, J.; Grant, S.F.A.; Hakonarson, H.; Bucan, M. PennCNV: An Integrated Hidden Markov Model Designed for High-Resolution Copy Number Variation Detection in Whole-Genome SNP Genotyping Data. Genome Res. 2007, 17, 1665–1674. [Google Scholar] [CrossRef]
- Colella, S.; Yau, C.; Taylor, J.M.; Mirza, G.; Butler, H.; Clouston, P.; Bassett, A.S.; Seller, A.; Holmes, C.C.; Ragoussis, J. QuantiSNP: An Objective Bayes Hidden-Markov Model to Detect and Accurately Map Copy Number Variation Using SNP Genotyping Data. Nucleic Acids Res. 2007, 35, 2013–2025. [Google Scholar] [CrossRef]
- Quinlan, A.R.; Hall, I.M. BEDTools: A Flexible Suite of Utilities for Comparing Genomic Features. Bioinformatics 2010, 26, 841–842. [Google Scholar] [CrossRef]
- Hou, Y.; Liu, G.E.; Bickhart, D.M.; Cardone, M.F.; Wang, K.; Kim, E.; Matukumalli, L.K.; Ventura, M.; Song, J.; VanRaden, P.M.; et al. Genomic Characteristics of Cattle Copy Number Variations. BMC Genom. 2011, 12, 127. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Lv, D.; Ge, Y.; Shi, J.; Weijers, D.; Yu, G.; Chen, J. RIdeogram: Drawing SVG Graphics to Visualize and Map Genome-Wide Data on the Idiograms. PeerJComput. Sci. 2020, 6, e251. [Google Scholar] [CrossRef] [PubMed]
- Kinsella, R.J.; Kahari, A.; Haider, S.; Zamora, J.; Proctor, G.; Spudich, G.; Almeida-King, J.; Staines, D.; Derwent, P.; Kerhornou, A.; et al. EnsemblBioMarts: A Hub for Data Retrieval across Taxonomic Space. Database 2011, 2011, bar030. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Ouyang, Y.; Yao, W. ShinyCircos: An R/Shiny Application for Interactive Creation of Circos Plot. Bioinformatics 2018, 34, 1229–1231. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.-L.; Fritz, E.R.; Reecy, J.M. AnimalQTLdb: A Livestock QTL Database Tool Set for Positional QTL Information Mining and Beyond. Nucleic Acids Res. 2007, 35, D604–D609. [Google Scholar] [CrossRef] [PubMed]
- Cattle QTL Database. Available online: https://www.animalgenome.org/cgi-bin/QTLdb/BT/summary (accessed on 21 June 2022).
- Draw Venn Diagram. Available online: http://bioinformatics.psb.ugent.be/webtools/Venn/ (accessed on 21 June 2022).
- Machado, M.A.; Azevedo, A.L.S.; Teodoro, R.L.; Pires, M.A.; Peixoto, M.G.; de Freitas, C.; Prata, M.C.; Furlong, J.; da Silva, M.V.; Guimarães, S.E.; et al. Genome Research Article Wide Scan for Quantitative Trait Loci Affecting Tick Resistance in Cattle (Bos Taurus × Bos Indicus). BMC Genom. 2010, 11, 280. [Google Scholar] [CrossRef]
- Otto, P.I.; Guimarães, S.E.F.; Verardo, L.L.; Azevedo, A.L.S.; Vandenplas, J.; Soares, A.C.C.; Sevillano, C.A.; Veroneze, R.; Pires, M.F.A.; de Freitas, C.; et al. Genome-Wide Association Studies for Tick Resistance in Bos Taurus × Bos Indicus Crossbred Cattle: A Deeper Look into This Intricate Mechanism. J. Dairy Sci. 2018, 101, 11020–11032. [Google Scholar] [CrossRef]
- Mapholi, N.O.; Maiwashe, A.; Matika, O.; Riggio, V.; Bishop, S.C.; MacNeil, M.D.; Banga, C.; Taylor, J.F.; Dzama, K. Genome-Wide Association Study of Tick Resistance in South African Nguni Cattle. Ticks Tick-Borne Dis. 2016, 7, 487–497. [Google Scholar] [CrossRef]
- Álvarez, I.; Fernández, I.; Traoré, A.; Menéndez-Arias, N.A.; Goyache, F. Population Structure Assessed Using Microsatellite and SNP Data: An Empirical Comparison in West African Cattle. Animals 2021, 11, 151. [Google Scholar] [CrossRef]
- Upadhyay, M.; Derks, M.F.L.; Andersson, G.; Medugorac, I.; Groenen, M.A.M.; Crooijmans, R.P.M.A. Introgression Contributes to Distribution of Structural Variations in Cattle. Genomics 2021, 113, 3092–3102. [Google Scholar] [CrossRef]
- Hu, Y.; Xia, H.; Li, M.; Xu, C.; Ye, X.; Su, R.; Zhang, M.; Nash, O.; Sonstegard, T.S.; Yang, L.; et al. Comparative Analyses of Copy Number Variations between Bos Taurus and Bos Indicus. BMC Genom. 2020, 21, 682. [Google Scholar] [CrossRef]
- Low, W.Y.; Tearle, R.; Liu, R.; Koren, S.; Rhie, A.; Bickhart, D.M.; Rosen, B.D.; Kronenberg, Z.N.; Kingan, S.B.; Tseng, E.; et al. Haplotype-Resolved Genomes Provide Insights into Structural Variation and Gene Content in Angus and Brahman Cattle. Nat. Commun. 2020, 11, 2071. [Google Scholar] [CrossRef]
- Rafter, P.; Purfield, D.C.; Berry, D.P.; Parnell, A.C.; Gormley, I.C.; Kearney, J.F.; Coffey, M.P.; Carthy, T.R. Characterization of Copy Number Variants in a Large Multibreed Population of Beef and Dairy Cattle Using High-Density Single Nucleotide Polymorphism Genotype Data1. J. Anim. Sci. 2018, 96, 4112–4124. [Google Scholar] [CrossRef]
- Liu, G.E.; Hou, Y.; Zhu, B.; Cardone, M.F.; Jiang, L.; Cellamare, A.; Mitra, A.; Alexander, L.J.; Coutinho, L.L.; Dell’Aquila, M.E.; et al. Analysis of Copy Number Variations among Diverse Cattle Breeds. Genome Res. 2010, 20, 693–703. [Google Scholar] [CrossRef]
- Upadhyay, M.; da Silva, V.H.; Megens, H.J.; Visker, M.H.P.W.; Ajmone-Marsan, P.; Bâlteanu, V.A.; Dunner, S.; Garcia, J.F.; Ginja, C.; Kantanen, J.; et al. Distribution and Functionality of Copy Number Variation across European Cattle Populations. Front. Genet. 2017, 8, 108. [Google Scholar] [CrossRef]
- Pérez-Pardal, L.; Sánchez-Gracia, A.; Álvarez, I.; Traoré, A.; Ferraz, J.B.S.; Fernández, I.; Costa, V.; Chen, S.; Tapio, M.; Cantet, R.J.C.; et al. Legacies of Domestication, Trade and Herder Mobility Shape Extant Male Zebu Cattle Diversity in South Asia and Africa. Sci. Rep. 2018, 8, 18027. [Google Scholar] [CrossRef]
- Pérez-Pardal, L.; Royo, L.J.; Beja-Pereira, A.; Curik, I.; Traoré, A.; Fernández, I.; Sölkner, J.; Alonso, J.; Álvarez, I.; Bozzi, R.; et al. Y-Specific Microsatellites Reveal an African Subfamily in Taurine (Bos Taurus) Cattle: Y-Chromosome Diversity in Taurine Cattle. Anim. Genet. 2010, 41, 232–241. [Google Scholar] [CrossRef]
- Jang, J.; Terefe, E.; Kim, K.; Lee, Y.H.; Belay, G.; Tijjani, A.; Han, J.-L.; Hanotte, O.; Kim, H. Population Differentiated Copy Number Variation of Bos Taurus, Bos Indicus and Their African Hybrids. BMC Genom. 2021, 22, 531. [Google Scholar] [CrossRef]
- Hughes, G.M.; Boston, E.S.M.; Finarelli, J.A.; Murphy, W.J.; Higgins, D.G.; Teeling, E.C. The Birth and Death of Olfactory Receptor Gene Families in Mammalian Niche Adaptation. Mol. Biol. Evol. 2018, 35, 1390–1406. [Google Scholar] [CrossRef]
- Jensen-Jarolim, E.; Pacios, L.F.; Bianchini, R.; Hofstetter, G.; Roth-Walter, F. Structural Similarities of Human and Mammalian Lipocalins, and Their Function in Innate Immunity and Allergy. Allergy 2016, 71, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Kongsuwan, K.; Josh, P.; Colgrave, M.L.; Bagnall, N.H.; Gough, J.; Burns, B.; Pearson, R. Activation of Several Key Components of the Epidermal Differentiation Pathway in Cattle Following Infestation with the Cattle Tick, Rhipicephalus (Boophilus) Microplus. Int. J. Parasitol. 2010, 40, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, S. TRIM Family Proteins: Roles in Autophagy, Immunity, and Carcinogenesis. Trends Biochim. Sci. 2017, 42, 297–311. [Google Scholar] [CrossRef] [PubMed]
- Ozato, K.; Shin, D.-M.; Chang, T.-H.; Morse, H.C. TRIM Family Proteins and Their Emerging Roles in Innate Immunity. Nat. Rev. Immunol. 2008, 8, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Malinowska, M.; Tokarz-Deptuła, B.; Deptuła, W. Butyrophilins: An Important New Element of Resistance. Cent. Eur. J. Immunol. 2017, 42, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Tone, K.; Stappers, M.H.T.; Willment, J.A.; Brown, G.D. C-Type Lectin Receptors of the Dectin-1 Cluster: Physiological Roles and Involvement in Disease. Eur. J. Immunol. 2019, 49, 2127–2133. [Google Scholar] [CrossRef] [PubMed]
- Carvalheiro, R.; Costilla, R.; Neves, H.H.R.; Albuquerque, L.G.; Moore, S.; Hayes, B.J. Unraveling Genetic Sensitivity of Beef Cattle to Environmental Variation under Tropical Conditions. Genet. Sel. Evol. 2019, 51, 29. [Google Scholar] [CrossRef]
- Granados, D.P.; Sriranganadane, D.; Daouda, T.; Zieger, A.; Laumont, C.M.; Caron-Lizotte, O.; Boucher, G.; Hardy, M.P.; Gendron, P.; Côté, C.; et al. Impact of Genomic Polymorphisms on the Repertoire of Human MHC Class I-Associated Peptides. Nat. Commun. 2014, 5, 3600. [Google Scholar] [CrossRef]
- Benavides, M.V.; Sonstegard, T.S.; van Tassell, C. Genomic Regions Associated with Sheep Resistance to Gastrointestinal Nematodes. Trends Parasitol. 2016, 32, 470–480. [Google Scholar] [CrossRef]
- Thompson-Crispi, K.A.; Sargolzaei, M.; Ventura, R.; Abo-Ismail, M.; Miglior, F.; Schenkel, F.; Mallard, B.A. A Genome-Wide Association Study of Immune Response Traits in Canadian Holstein Cattle. BMC Genom. 2014, 15, 559. [Google Scholar] [CrossRef]
- Babcock, J.; Li, M. Deorphanizing the Human Transmembrane Genome: A Landscape of Uncharacterized Membrane Proteins. Acta Pharmacol. Sin. 2014, 35, 11–23. [Google Scholar] [CrossRef]
- Pérez-Enciso, M.; Varona, L. Quantitative Trait Loci Mapping in F2 Crosses Between Outbred Lines. Genetics 2000, 155, 391–405. [Google Scholar] [CrossRef]
- Gautier, M.; Flori, L.; Riebler, A.; Jaffrézic, F.; Laloé, D.; Gut, I.; Moazami-Goudarzi, K.; Foulley, J.L. A Whole Genome Bayesian Scan for Adaptive Genetic Divergence in West African Cattle. BMC Genom. 2009, 10, 550. [Google Scholar] [CrossRef]
- Kim, K.; Kwon, T.; Dessie, T.; Yoo, D.; Mwai, O.A.; Jang, J.; Sung, S.; Lee, S.; Salim, B.; Jung, J.; et al. The Mosaic Genome of Indigenous African Cattle as a Unique Genetic Resource for African Pastoralism. Nat. Genet. 2020, 52, 1099–1110. [Google Scholar] [CrossRef]
- Yougbaré, B.; Soudré, A.; Ouédraogo, D.; Zoma, B.L.; Tapsoba, A.S.R.; Sanou, M.; Ouédraogo-Koné, S.; Burger, P.A.; Wurzinger, M.; Khayatzadeh, N.; et al. Genome-Wide Association Study of Trypanosome Prevalence and Morphometric Traits in Purebred and Crossbred Baoulé Cattle of Burkina Faso. PLoS ONE 2021, 16, e0255089. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Cluster (Enrichment Score) | Category | Term and Description | Genes a | p-Value | Fold Enrichment |
|---|---|---|---|---|---|
| Annotation Cluster 1 (2.27) | INTERPRO | IPR011038: Calycin-like | THAP4, LOC618559, PAEP, LCN9, LCN12, PTGDS, ENSBTAG00000030296 | 2.28 × 10−3 | 5.08 |
| INTERPRO | IPR000566: Lipocalin/cytosolic fatty-acid binding protein domain | 7.28 × 10−3 | 4.85 | ||
| INTERPRO | IPR012674: Calycin | 9.22 × 10−3 | 4.59 | ||
| Annotation Cluster 2 (1.47) | SMART | SM00449: SPRY | LOC526787, RNF123, LOC525599, LOC504295, SPSB3, TRIML2, TRIML1 | 2.86 × 10−2 | 2.99 |
| INTERPRO | IPR003877: SPla/RYanodine receptor SPRY | 0.031 | 2.96 | ||
| INTERPRO | IPR001870:B30.2/SPRY domain | 0.044 | 2.71 | ||
| Annotation Cluster 3 (1.44) | INTERPRO | IPR016186:C-type lectin-like | COL18A1, NKG2C, KLRD1, KLRK1, ENSBTAG00000000966, LOC100848575, OLR1, REG3G, REG3A | 2.93× 10−2 | 2.47 |
| SMART | SM00034: CLECT | 0.036 | 2.56 | ||
| INTERPRO | IPR001304:C-type lectin | 3.98 × 10−2 | 2.51 | ||
| INTERPRO | IPR016187:C-type lectin fold | 4.28 × 10−2 | 2.29 |
| Candidate CNVR | BTA | QTL Start (bp) | QTL End (bp) | QTL_ID | Trait | Candidate Genes |
|---|---|---|---|---|---|---|
| CNVRt4 | 1 | 12,484,941 | 34,327,730 | 10506 | Initial packed red blood cell volume a | ROBO2 (AC17, AC30) |
| CNVRt195 | 11 | 104,664,393 | 104,664,433 | 101155 | Tick resistance c | VAV2 (AC22, AC23) |
| 101169 | Tick resistance c | |||||
| CNVRt214 | 13 | 17,709,118 | 53,561,417 | 10524 | Percentage decrease in PCV up to day 100 after challenge a | NET1 (AC23) |
| 10525 | Percentage decrease in PCV up to day 100 after challenge a | |||||
| 10526 | Parasite detection rate a | |||||
| CNVRt303 | 20 | 12,158,768 | 22,679,451 | 10537 | PCVI minus PCVM a | ELOVL7 (AC30), LOC107131564, DEPDC1B (AC18) |
| 10538 | PCV variance a | |||||
| 10539 | Percentage decrease in PCV up to day 100 after challenge a | |||||
| 10540 | Percentage decrease in body weight up today 150 after challenge a | |||||
| 10541 | Parasite natural logarithm of mean number a | |||||
| CNVRt321 | 22 | 4,775,998 | 28,467,455 | 10542 | Initial packed red blood cell volume a | SCN5A (AC30) |
| CNVRt334 | 23 | 4,584,848 | 29,554,995 | 10543 | Parasite detection rate a | FAM83B |
| CNVRt335 | 23 | 4,584,848 | 29,554,995 | 10543 | Parasite detection rate a | BAK1, LOC516410, ITPR3 (AC23, AC30), MNF1, IP6K3 (AC30), LEMD2 (AC30), MLN |
| CNVRt336 | 23 | 4,584,848 | 29,554,995 | 10543 | Parasite detection rate a | KCNK5 (AC30, AC5), KCNK17 (AC30, AC5) |
| 13,002,524 | 13,489,143 | 164932 | Tick resistance d | |||
| 13,002,524 | 13,489,143 | 164933 | Tick resistance d | |||
| CNVRt338 | 23 | 4,584,848 | 29,554,995 | 10543 | Parasite detection rate a | ENSBTAG00000048364, ENSBTAG00000038397, ENSBTAG00000015565 (AC30), BOLA-DRA (AC30), BTNL2 (AC30), LOC525599 (AC2, AC30), LOC504295 (AC2), ENSBTAG00000026163, ENSBTAG00000050817 |
| 22,432,428 | 29,658,279 | 12223 | Tick resistance b | |||
| CNVRt341 | 23 | 4,584,848 | 29,554,995 | 10543 | Parasite detection rate a | ENSBTAG00000054439, RNF39, PPP1R11, POLR1H (AC31), ENSBTAG00000054588, ZFP57 (AC37), MOG (AC30), GABBR1(AC30, AC8), ENSBTAG00000031825, LOC504548, ENSBTAG00000052703, OR2H1D (AC30, AC40,AC41), OR2H1 (AC30,AC40,AC41) |
| 22,432,428 | 29,658,279 | 12223 | Tick resistance b | |||
| 28,511,975 | 39,129,003 | 12224 | Tick resistance b | |||
| CNVRt342 | 23 | 4,584,848 | 29,554,995 | 10543 | Parasite detection rate a | OR12D23, OR12D18, OR12D2H, OR12D2E (AC30, AC40, AC41) |
| 22,432,428 | 29,658,279 | 12223 | Tick resistance b | |||
| 28,511,975 | 39,129,003 | 12224 | Tick resistance b | |||
| CNVRt345 | 23 | 28,511,975 | 39,129,003 | 12224 | Tick resistance b | ZKSCAN8 (AC31, AC37) |
| CNVRt346 | 23 | 28,511,975 | 39,129,003 | 12224 | Tick resistance b | ENSBTAG00000038430, PRP4 (AC9), PRP8 (AC9), PRP6 (AC9), PRP1, LOC100298767 (AC9), PRP2 |
| CNVRt349 | 24 | 14,587,194 | 22,401,178 | 10544 | Initial packed red blood cell volume a | ENSBTAG00000044087 |
| 10544 | Percentage decrease in PCV up to day 100 after challenge a | |||||
| CNVRt364 | 26 | 5,469,941 | 18,747,171 | 10548 | Percentage decrease in PCV up to day 100 after challenge a | LOC540627 |
| 10549 | Percentage decrease in PCV up to day 100 after challenge a | |||||
| 10550 | BWF scaled by BWI a | |||||
| 10551 | Body weight (mean) a | |||||
| 10552 | Percentage decrease in body weight up today 150 after challenge a | |||||
| CNVRt369 | 27 | 13,701,246 | 20,999,044 | 10553 | PCVI minus PCVM a | ZFP42 (AC37), TRIML2 (AC2), TRIML1 (AC2) |
| 10554 | PCV variance a | |||||
| 10555 | Percentage decrease in PCV up to day 100 after challenge a | |||||
| 10556 | Percentage decrease in PCV up to day 100 after challenge a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goyache, F.; Pérez-Pardal, L.; Fernández, I.; Traoré, A.; Menéndez-Arias, N.A.; Arias, K.D.; Álvarez, I. Identification and Characterization of Copy Number Variations Regions in West African Taurine Cattle. Animals 2022, 12, 2130. https://doi.org/10.3390/ani12162130
Goyache F, Pérez-Pardal L, Fernández I, Traoré A, Menéndez-Arias NA, Arias KD, Álvarez I. Identification and Characterization of Copy Number Variations Regions in West African Taurine Cattle. Animals. 2022; 12(16):2130. https://doi.org/10.3390/ani12162130
Chicago/Turabian StyleGoyache, Félix, Lucía Pérez-Pardal, Iván Fernández, Amadou Traoré, Nuria A. Menéndez-Arias, Katherine D. Arias, and Isabel Álvarez. 2022. "Identification and Characterization of Copy Number Variations Regions in West African Taurine Cattle" Animals 12, no. 16: 2130. https://doi.org/10.3390/ani12162130