
Development and Preliminary Evaluation of a Nanoparticle-Assisted PCR Assay for the Detection of Cryptosporidium parvum in Calves

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Parasites, Clinical Samples, and gDNA Samples
2.2. PCR Primers
2.3. Nested PCR
2.4. Optimization of the Normal PCR Assay for C. parvum
2.5. Optimization of the Nano-PCR Assay for C. parvum
2.6. Sensitivities of the Normal PCR and the Nano-PCR Assays
2.7. Specificities of the Normal PCR and the Nano-PCR Assays
2.8. Assessment of the Nano-PCR in Faecal Samples
3. Results
3.1. Optimization of the Normal PCR and the Nano-PCR for C. parvum
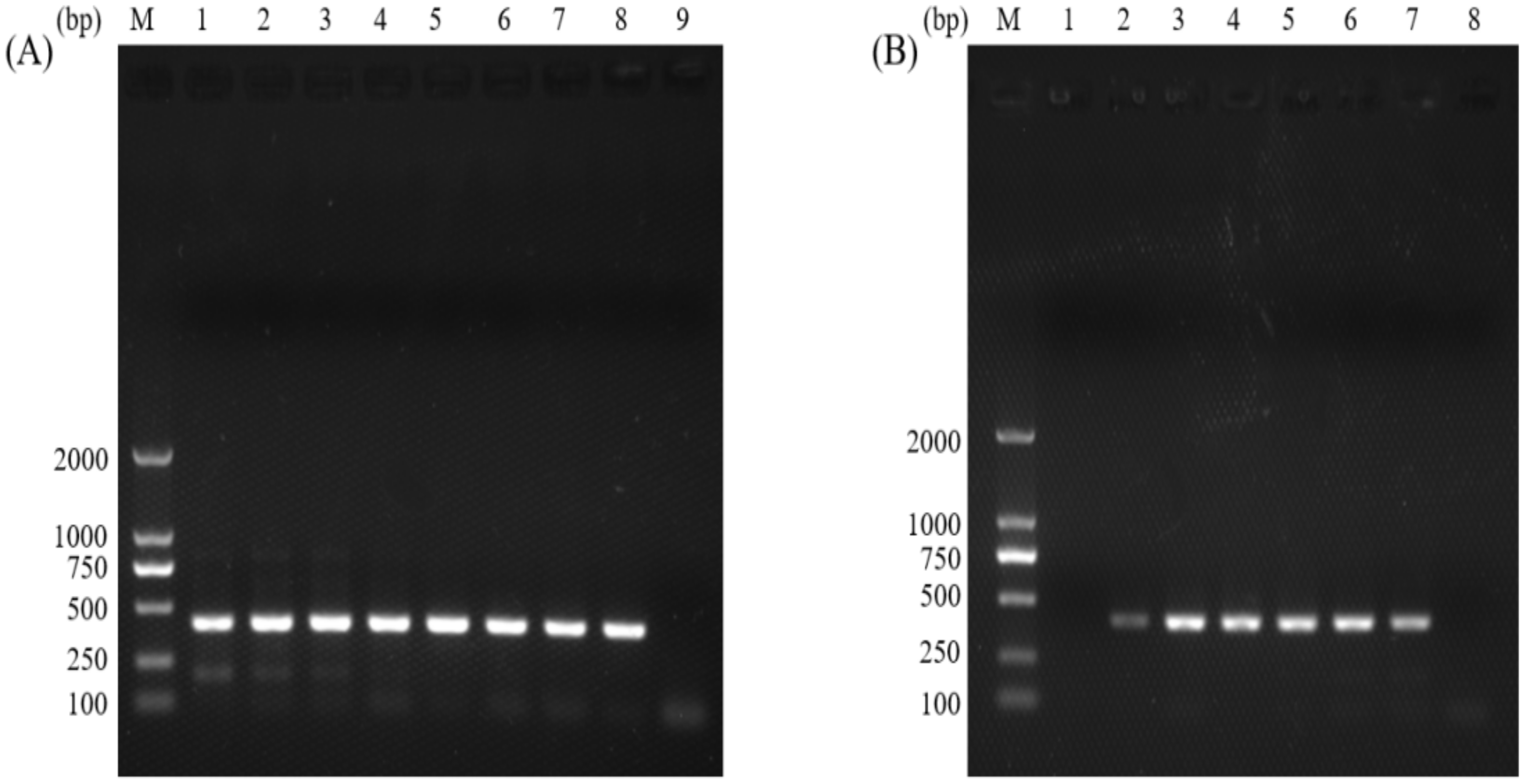
3.2. Sensitivities of the Normal PCR and the Nano-PCR Assays
3.3. Specificities of the Normal PCR and the Nano-PCR Assays
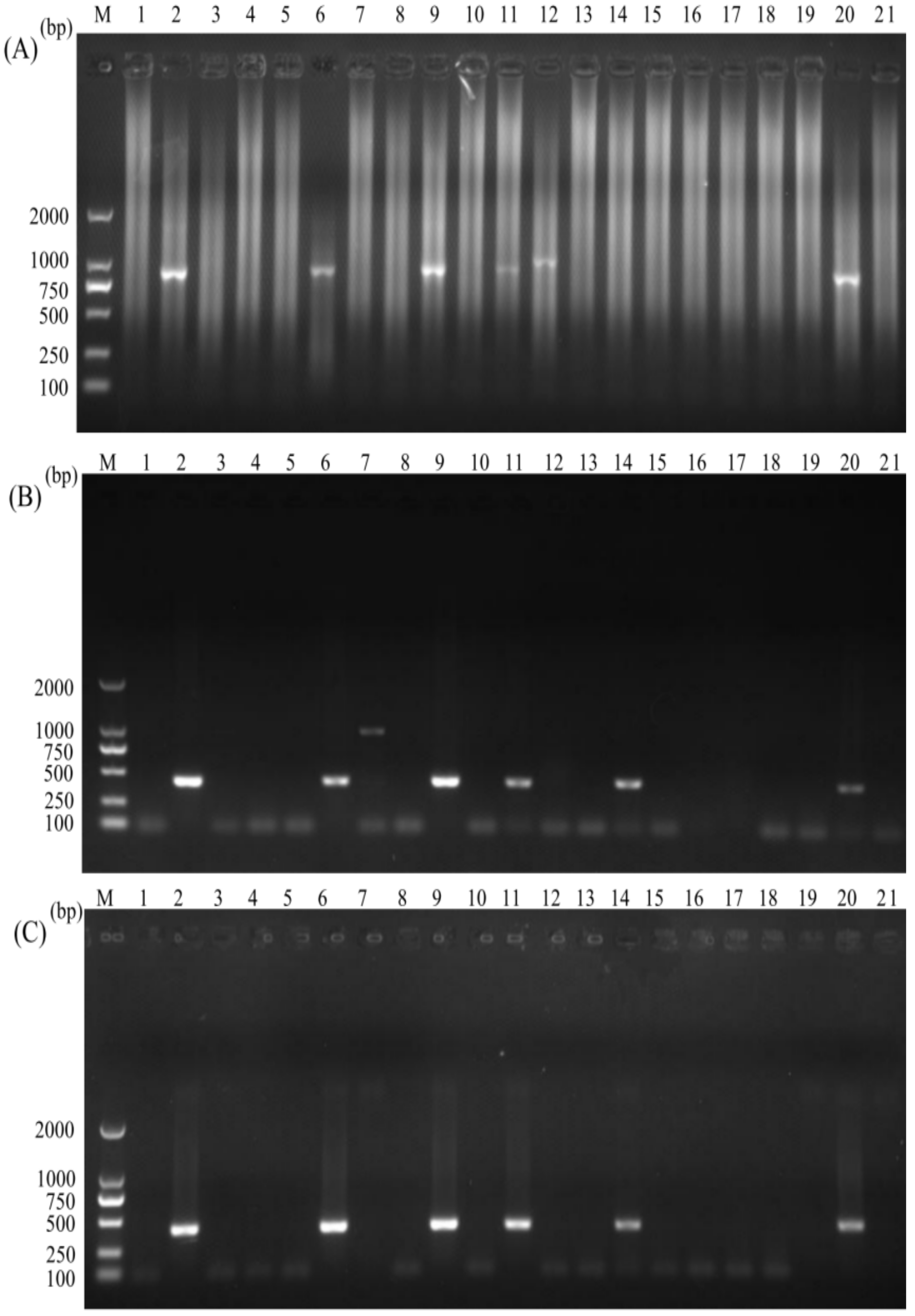
3.4. Assessment of the Nano-PCR for C. parvum in Clinical Settings
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de Graaf, D.C.; Vanopdenbosch, E.; Ortega-Mora, L.M.; Abbassi, H.; Peeters, J.E. A review of the importance of cryptosporidiosis in farm animals. Int. J. Parasitol. 1999, 29, 1269–1287. [Google Scholar] [CrossRef]
- Santín, M. Cryptosporidium and Giardia in ruminants. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 223–238. [Google Scholar] [CrossRef]
- Santín, M.; Trout, J.M.; Xiao, L.; Zhou, L.; Greiner, E.; Fayer, R. Prevalence and age-related variation of Cryptosporidium species and genotypes in dairy calves. Vet. Parasitol. 2004, 122, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Fayer, R.; Santín, M.; Trout, J.M.; Greiner, E. Prevalence of species and genotypes of Cryptosporidium found in 1–2-year-old dairy cattle in the eastern United States. Vet. Parasitol. 2006, 135, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Santín, M.; Trout, J.M.; Fayer, R. A longitudinal study of cryptosporidiosis in dairy cattle from birth to 2 years of age. Vet. Parasitol. 2008, 155, 15–23. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, J.; Chang, Y.; Yu, F.; Zhang, S.; Wang, R.; Zhang, L. Prevalence and molecular characterization of Cryptosporidium spp. and Giardia duodenalis in dairy cattle in Gansu, northwest China. Parasite 2020, 27, 62. [Google Scholar] [CrossRef]
- Thomson, S.; Hamilton, C.A.; Hope, J.C.; Katzer, F.; Mabbott, N.A.; Morrison, L.J.; Innes, E.A. Bovine cryptosporidiosis: Impact, host-parasite interaction and control strategies. Vet. Res. 2017, 48, 42. [Google Scholar] [CrossRef] [Green Version]
- Khurana, S.; Sharma, P.; Sharma, A.; Malla, N. Evaluation of Ziehl-Neelsen staining, auramine phenol staining, antigen detection enzyme linked immunosorbent assay and polymerase chain reaction, for the diagnosis of intestinal cryptosporidiosis. Trop. Parasitol. 2012, 2, 20–23. [Google Scholar] [CrossRef] [PubMed]
- Koehler, A.V.; Jex, A.R.; Haydon, S.R.; Stevens, M.A.; Gasser, R.B. Giardia/giardiasis-a perspective on diagnostic and analytical tools. Biotechnol. Adv. 2014, 32, 280–289. [Google Scholar] [CrossRef]
- Alves, M.; Xiao, L.; Sulaiman, I.; Lal, A.A.; Matos, O.; Antunes, F. Subgenotype analysis of Cryptosporidium isolates from humans, cattle, and zoo ruminants in Portugal. J. Clin. Microbiol. 2003, 41, 2744–2747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Ryan, U.M. Cryptosporidiosis: An update in molecular epidemiology. Curr. Opin. Infect. Dis. 2004, 17, 483–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L. Molecular epidemiology of cryptosporidiosis: An update. Exp. Parasitol. 2010, 124, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Balatbat, A.B.; Jordan, G.W.; Tang, Y.J.; Silva, J., Jr. Detection of Cryptosporidium parvum DNA in human feces by nested PCR. J. Clin. Microbiol. 1996, 34, 1769–1772. [Google Scholar] [CrossRef] [Green Version]
- Demming, A. The state of research after 25 years of nanotechnology. Nanotechnology 2014, 25, 010201. [Google Scholar] [CrossRef]
- Ali, Z.; Jin, G.; Hu, Z.; Wang, Z.; Khan, M.A.; Dai, J.; Tang, Y. A review on nanoPCR: History, mechanism and applications. J. Nanosci. Nanotechnol. 2018, 18, 8029–8046. [Google Scholar] [CrossRef] [PubMed]
- Gedda, M.R.; Madhukar, P.; Shukla, A.; Mudavath, S.L.; Srivastava, O.N.; Singh, O.P.; Sundar, S. Nanodiagnostics in leishmaniasis: A new frontiers for early elimination. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2021, 13, e1675. [Google Scholar] [CrossRef]
- Li, M.; Lin, Y.C.; Wu, C.C.; Liu, H.S. Enhancing the efficiency of a PCR using gold nanoparticles. Nucleic Acids Res. 2005, 33, e184. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, S.; Rasheed, A.K.; Siddiqui, R.; Appaturi, J.N.; Fen, L.B.; Khan, N.A. Development of nanoparticle-assisted PCR assay in the rapid detection of brain-eating amoebae. Parasitol. Res. 2018, 117, 1801–1811. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Sarwar, Y.; Raza, Z.A.; Hussain, S.Z.; Mustafa, T.; Khan, W.S.; Ghauri, M.A.; Haque, A.; Hussain, I. Metal nanoparticle assisted polymerase chain reaction for strain typing of Salmonella typhi. Analyst 2015, 140, 7366–7372. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Liang, L.; Luo, Y.; Wang, G.; Wang, C.; Cui, Y.; Ai, X.; Cui, S. A sensitive duplex nanoparticle-assisted PCR assay for identifying porcine epidemic diarrhea virus and porcine transmissible gastroenteritis virus from clinical specimens. Virus Genes 2017, 53, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, J.; Liu, Z.; Li, J.; Li, Z.; Wang, C.; Wang, J.; Guo, L. Development of a nanoparticle-assisted PCR assay for detection of bovine respiratory syncytial virus. BMC Vet. Res. 2019, 15, 110. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.L.; Wang, Y.; Lai, P.; Yao, Q.; Li, Y.; Zhang, L.X.; Yang, X.; Song, J.K.; Zhao, G.H. Establishment and preliminary application of nanoparticle-assisted PCR assay for detection of Cryptosporidium spp. Parasitol. Res. 2021, 120, 1837–1844. [Google Scholar] [CrossRef] [PubMed]
- Luan, Q.; Jiang, Z.; Wang, D.; Wang, S.; Yin, Y.; Wang, J. A sensitive triple nanoparticle-assisted PCR assay for detection of fowl adenovirus, infectious bursal disease virus and chicken anemia virus. J. Virol. Methods 2022, 303, 114499. [Google Scholar] [CrossRef]
- Zhao, S.S.; Li, Y.H.; Zhang, Y.; Zhou, Q.; Jing, B.; Xu, C.Y.; Zhang, L.X.; Song, J.K.; Qi, M.; Zhao, G.H. Multilocus genotyping of Giardia duodenalis in Bactrian camels (Camelus bactrianus) in China. Parasitol. Res. 2020, 119, 3873–3880. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.H.; Yao, Q.; Dong, H.P.; Wang, S.S.; Chen, R.R.; Song, J.K.; Yan, W.C.; Zhao, G.H. Molecular characterization of Balantioides coli in pigs from Shaanxi province, northwestern China. Parasitol. Res. 2020, 119, 3075–3081. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, Y.H.; Wang, Y.X.; Wang, J.W.; Lai, P.; Li, Y.; Song, J.K.; Qi, M.; Zhao, G.H. Molecular characterization of Blastocystis sp. in Camelus bactrianus in Northwestern China. Animals 2021, 11, 3016. [Google Scholar] [CrossRef]
- Wang, S.S.; Li, J.Q.; Li, Y.H.; Wang, X.W.; Fan, X.C.; Liu, X.; Li, Z.J.; Song, J.K.; Zhang, L.X.; Zhao, G.H. Novel genotypes and multilocus genotypes of Enterocytozoon bieneusi in pigs in northwestern China: A public health concern. Infect. Genet. Evol. 2018, 63, 89–94. [Google Scholar] [CrossRef]
- Wang, X.T. Study on Population Structure of Cryptosporidium spp. and Giardia lamblia in in Calves from Partial Areas of Shaanxi. Master Thesis, Northwest A&F University, Xianyang, China, 2017. (In Chinese). [Google Scholar]
- Adeyemo, F.E.; Singh, G.; Reddy, P.; Stenström, T.A. Methods for the detection of Cryptosporidium and Giardia: From microscopy to nucleic acid based tools in clinical and environmental regimes. Acta Trop. 2018, 184, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Destura, R.V.; Rohani, C.B.; Ma, J.; Sevilleja, J.E.A.D. Advancing Cryptosporidium diagnostics from bench to bedside. Curr. Trop. Med. Rep. 2015, 2, 150–160. [Google Scholar] [CrossRef] [Green Version]
- Cunha, F.S.; Peralta, R.H.S.; Peralta, J.M. New insights into the detection and molecular characterization of Cryptosporidium with emphasis in Brazilian studies: A review. Rev. Inst. Med. Trop. Sao Paulo 2019, 61, e28. [Google Scholar] [CrossRef] [PubMed]
- Mesa, L.E.; Manrique, R.; Muskus, C.; Robledo, S.M. Test accuracy of polymerase chain reaction methods against conventional diagnostic techniques for Cutaneous Leishmaniasis (CL) in patients with clinical or epidemiological suspicion of CL: Systematic review and meta-analysis. PLoS Negl. Trop. Dis. 2020, 14, e0007981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezzaty Mirhashemi, M.; Zintl, A.; Grant, T.; Lucy, F.E.; Mulcahy, G.; De Waal, T. Comparison of diagnostic techniques for the detection of Cryptosporidium oocysts in animal samples. Exp. Parasitol. 2015, 151–152, 14–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Bern, C.; Limor, J.; Sulaiman, I.; Roberts, J.; Checkley, W.; Cabrera, L.; Gilman, R.H.; Lal, A.A. Identification of 5 types of Cryptosporidium parasites in children in Lima, Peru. J. Infect. Dis. 2001, 183, 492–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, S.C.; Laperriere, G.; Germain, H. Droplet digital PCR versus qPCR for gene expression analysis with low abundant targets: From variable nonsense to publication quality data. Sci. Rep. 2017, 7, 2409. [Google Scholar] [CrossRef] [Green Version]
- Hadfield, S.J.; Robinson, G.; Elwin, K.; Chalmers, R.M. Detection and differentiation of Cryptosporidium spp. in human clinical samples by use of real-time PCR. J. Clin. Microbiol. 2011, 49, 918–924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, R.; Paparini, A.; Monis, P.; Ryan, U. Comparison of next-generation droplet digital PCR (ddPCR) with quantitative PCR (qPCR) for enumeration of Cryptosporidium oocysts in faecal samples. Int. J. Parasitol. 2014, 44, 1105–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakheit, M.A.; Torra, D.; Palomino, L.A.; Thekisoe, O.M.; Mbati, P.A.; Ongerth, J.; Karanis, P. Sensitive and specific detection of Cryptosporidium species in PCR-negative samples by loop-mediated isothermal DNA amplification and confirmation of generated LAMP products by sequencing. Vet. Parasitol. 2008, 158, 11–22. [Google Scholar] [CrossRef]
- Karanis, P.; Thekisoe, O.; Kiouptsi, K.; Ongerth, J.; Igarashi, I.; Inoue, N. Development and preliminary evaluation of a loop-mediated isothermal amplification procedure for sensitive detection of Cryptosporidium oocysts in fecal and water samples. Appl. Environ. Microbiol. 2007, 73, 5660–5662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichols, R.A.B.; Campbell, B.M.; Smith, H.V. Identification of Cryptosporidium spp. oocysts in United Kingdom noncarbonated natural mineral waters and drinking waters by using a modified nested PCR-restriction fragment length polymorphism assay. Appl. Environ. Microbiol. 2003, 69, 4183–4189. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Wang, R.; Cai, M.; Jiang, W.; Feng, Y.; Xiao, L. Outbreak of cryptosporidiosis due to Cryptosporidium parvum subtype IIdA19G1 in neonatal calves on a dairy farm in China. Int. J. Parasitol. 2019, 49, 569–577. [Google Scholar] [CrossRef]
- Li, N.; Zhao, W.; Song, S.; Ye, H.; Chu, W.; Guo, Y.; Feng, Y.; Xiao, L. Diarrhoea outbreak caused by coinfections of Cryptosporidium parvum subtype IIdA20G1 and rotavirus in pre-weaned dairy calves. Transbound. Emerg. Dis. 2022; in press. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCR Assays | Primer Names | Sequences (5′–3′) | Product Sizes (bp) |
|---|---|---|---|
| Normal PCR /Nano-PCR | cgd3_330-F | AGTGGTTACAGGTGGGATGAGT | ~413 |
| cgd3_330-R | GCGAGTTTCCTTGATTCATAGC | ||
| Nested PCR | gp60-F1 | TTACTCTCCGTTATAGTCTCC | ~915 |
| gp60-R1 | GGAAGGAACGATGTATCTGA | ||
| gp60-F2 | TCCGCTGTATTCTCAGCC | ~800 | |
| gp60-R2 | GCAGAGGAACCAGCATC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, Q.; Yang, X.; Wang, Y.; Wang, J.; Huang, S.; Song, J.; Zhao, G. Development and Preliminary Evaluation of a Nanoparticle-Assisted PCR Assay for the Detection of Cryptosporidium parvum in Calves. Animals 2022, 12, 1953. https://doi.org/10.3390/ani12151953
Yao Q, Yang X, Wang Y, Wang J, Huang S, Song J, Zhao G. Development and Preliminary Evaluation of a Nanoparticle-Assisted PCR Assay for the Detection of Cryptosporidium parvum in Calves. Animals. 2022; 12(15):1953. https://doi.org/10.3390/ani12151953
Chicago/Turabian StyleYao, Qian, Xin Yang, Yi Wang, Junwei Wang, Shuang Huang, Junke Song, and Guanghui Zhao. 2022. "Development and Preliminary Evaluation of a Nanoparticle-Assisted PCR Assay for the Detection of Cryptosporidium parvum in Calves" Animals 12, no. 15: 1953. https://doi.org/10.3390/ani12151953