1. Introduction
Broilers make a major contribution to human protein consumption in many regions of the world; to achieve this, the poultry industry has adopted high stocking densities to maximize output and profitability [
1,
2]. Compared with their wild ancestors, the ability of chickens to cope with the stressors in an outdoor environment appears to have decreased [
3]. Temperatures above 30 °C and relative humidity levels above 80% inside the housing, as observed in tropical environments during summer, can cause heat stress in fast-growing chickens, which adversely affects growth rates, immune functions, disease susceptibility, and potentially death by heat exhaustion [
4,
5]. Coping strategies during stressful circumstances require major physiological and behavioral adjustments [
6]. Physiological responses include an increase in the heterophil to lymphocyte (H/L) ratio, due to heterophilia and lymphopenia [
7], and increased weight of lymphoid organs, through immune activation [
8]. Both are good indicators of stress and immune status.
Behavioral responses, such as shade seeking or increased standing to allow increased ventilation, require the birds to walk, run, and stand. This could be dependent on leg health, which can be assessed by the Seedor index of bone density [
9].
Despite these potential problems in the tropics, there is a growing body of literature that recognizes the importance of allowing chickens to express their natural behavior by providing outdoor access [
10,
11,
12], particularly at low stocking densities [
13]. Indoor stocking densities may influence whether outdoor access is beneficial or not. However, there have been few studies investigating the effects of offering outdoor access to broilers housed under tropical climatic conditions. In an earlier study of the provision of outdoor area for broilers in the winter in the tropics [
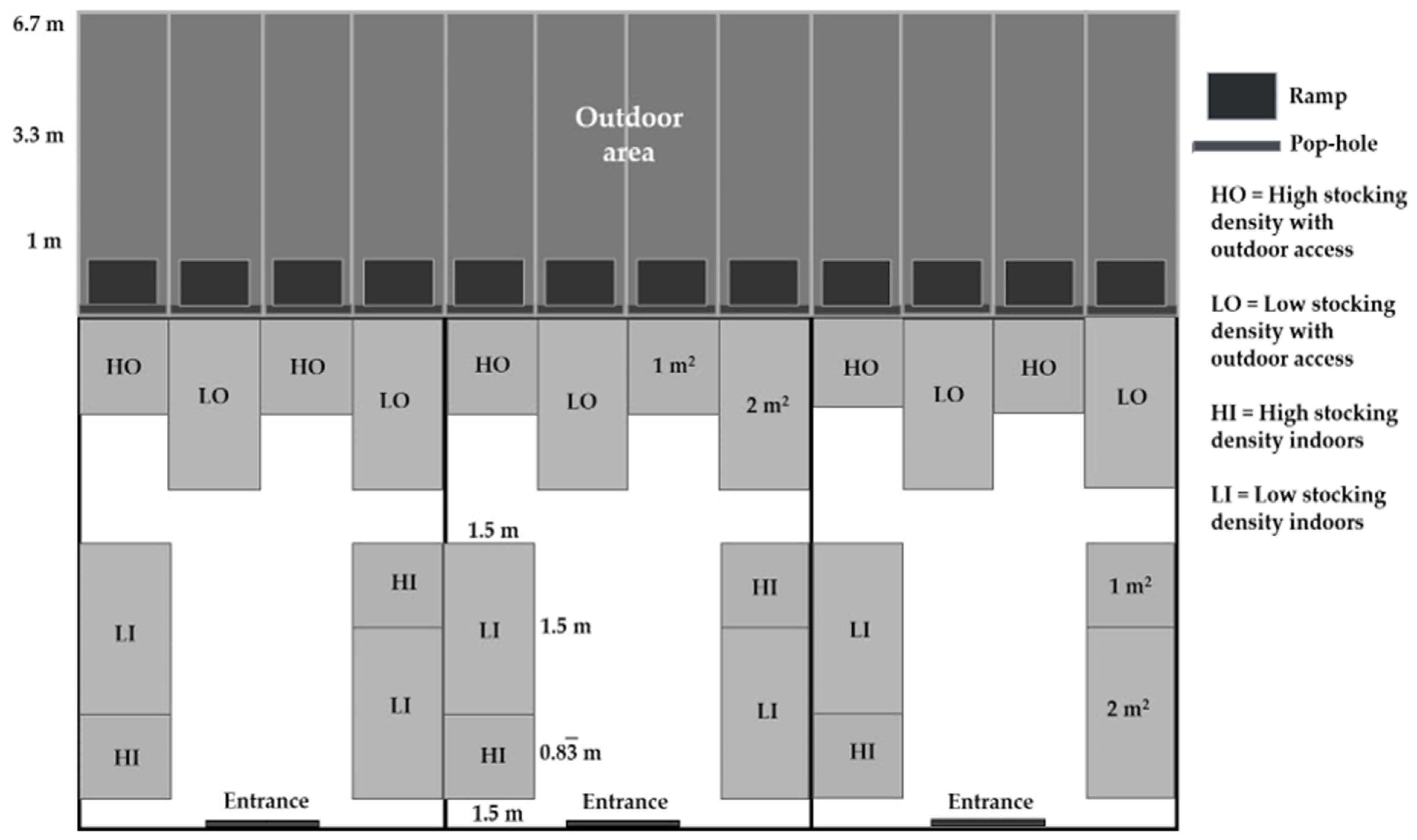
13], we found that birds at a high stocking density with no outdoor pens walked and preened themselves less and laid down more. Outdoor access increased foraging but only when birds were stocked at a low stocking density indoors; these birds also appeared more responsive to stressors, with elevated corticosterone and reduced spleen and bursa of Fabricius weights. Therefore, the aim of this study was to evaluate, in two contrasting seasons of the year, the effects on behavior and stress responses to allow chickens outdoor access at two indoor stocking densities. The effects on bird growth and carcass composition were published previously [
14], showing that the low stocking density and outdoor access had positive effects on chicken growth in winter. Summer temperatures were sufficient to cause heat stress, as evidenced by an increase in mortality. Birds reared in summer without outdoor access, or with outdoor access but a high stocking density inside, had reduced growth and feed conversion efficiency compared with birds reared in winter. Because of the evident effects of outdoor access on walking and lying that we previously recorded [
13], the effects on leg health (footpad dermatitis and bone conformation) were included in this study.
4. Discussion
This study was conducted with typical conditions for the tropics. Temperatures were high in both summer and winter, above the upper critical temperature, which has been estimated between 23.9 and 25.5 °C [
21], but in summer the heat stress was exacerbated by high humidity. In our facility, the outdoor temperatures tended to be higher than those recorded indoors.
Outdoor access mainly affected resting behavior and tibiae conformation. A reduction in resting behavior was most evident in summer, both in older birds and in birds at low densities. The potential for heat stress in summer is sufficient to cause increased mortality in this climate [
13,
14]. The reduced resting time in outdoor birds in summer, compared with indoor birds, suggests that the high outdoor temperatures stimulated activity. This was most evident for birds at low densities and older birds. If outdoor access was sufficient to cause heat stress, it is possible that there was sufficient space for some subordinate birds to be forced by dominant birds to stay outside at low densities, where the high temperatures stimulated activity and reduced growth and feed conversion efficiency [
14]. High-density indoors would probably offer less opportunity for agonistic bird interactions.
Low densities had welfare benefits of encouraging more activity in the birds, locomotion, preening, and foraging, and high densities encouraged polydipsia and reduced activity. The decreased drinking in birds at high densities in summer may have been connected to the increased resting, standing, and locomotion of these birds at this time. The close proximity of other birds is likely to have stimulated locomotion and standing. This in turn would have necessitated more time spent resting. Conversely, birds at low densities spent more time foraging, conceivably because they were less stressed, and in winter they spent more time preening, probably because of reduced disturbances by other birds.
As expected, drinking behavior was particularly high in week 6 of the experimental period (42 d old birds) in summer, when high temperatures were recorded. This is in accordance with the findings of Bruno et al. [
22], who reported that high environmental temperatures increase this behavior. The bell-type drinkers used in our study promoted high water intake, which is important for broilers in terms of biochemical and physiological function for homeostasis and growth.
The resting behavior observed more frequently in both outdoor access treatments and densities during winter is in accordance with the findings of Sanchez-Casanova et al. [
13], demonstrating a dependency of this response on season. Since resting is a comfort behavior, chickens spend up to 76% of their time in it [
23], but it can be a response to restricted space or leg disease [
24,
25]. However, despite the reduced percentage of foot pad dermatitis observed in winter, the fact that chickens at 42 days old were heavier in low-density treatments, with inferior tibia bone development at this age, could have led to the reduced physical activity, decreasing locomotion and increasing resting behavior.
Standing behavior followed a pattern that could be explained by the opposite effect described for resting, with similar consequences derived from bone quality. This behavior was more frequently observed in summer and the effects of stocking density were also dependent on season since the high- and low-density treatments showed an opposite trend. In summer, more birds were found standing in the high-density treatments. In terms of age, there was also an opposite trend in winter, since more birds were observed standing at 42 days old, in contrast to those at 28 days old. Standing has been described as a sign of good leg health, but is also considered as an avoidance response when discomfort is present [
26], and it can also be a response to heat stress.
There was a higher percentage of FPD lesions, poorer tibia bone properties, and the hardest environmental conditions observed in summer, which could be increased by high stocking densities [
27]. The pain produced by FPD ulcerative lesions in conjunction with high temperatures and humidity observed in week 4 of the experimental period in summer (
Table 2) could have increased the frequency of this avoidance behavior (standing), which was exacerbated by acute and chronic pain derived from an impaired leg bone development [
23,
26].
More birds were observed preening in summer, at both low and high stocking densities, at 28 and 42 days old, but with no difference between outdoor and indoor treatments. Although birds usually prefer to preen themselves indoors [
28], increased preening has been related with mild frustration [
29] which could be provoked by thermal discomfort in summer. However, the fact that this behavior increased at the end of the experimental period agrees with the findings of Fortomaris et al. [
30], who suggested that most husbandry systems allow full expression of this behavior.
Foraging is a highly motivated behavior [
31] and is a good indicator of a comfortable state in chickens. In this study, low-density treatments promoted foraging in birds raised in summer which can be attributable to more space allowance and, therefore, less effects of hard environmental conditions. In addition, litter material could have served as a source of enrichment, leading to a less restricted natural behavior.
Heterophils increased in summer at 42 days old, and the higher H/L ratio observed in winter in the LO treatment, at 28 days old, as a result of heterophilia, was contrary to the findings of Osti et al. [
32], whereby the H/L ratio was greater during summer, in accordance with Sanchez-Casanova et al. [
13], who found a higher H/L ratio in low-density treatments in winter, and Rajkumar et al. [
33], whose findings demonstrated a lower H/L ratio in summer. An increase in the H/L ratio is commonly observed during mild to moderate stress [
34,
35] and is also associated with elevated serum corticosterone concentrations [
36]. Multiple stressors can increase this ratio such as fearfulness and heat stress. Although fearfulness was not measured in this study, it is well known that handling procedures and external factors, such as noise or predators, could trigger fear, primarily around young ages due to little experience being outside [
37]. Heat stress is often observed when metabolic heat production is greater than the amount of heat that can be lost to the environment, so body heat and body temperature increase [
38]. The duration of heat exposure plays an important role in the H/L ratio [
39]. Even though outdoor access could potentially improve chickens’ welfare by providing extra space, it could also expose chickens to high temperatures [
40]. Tropical environmental temperatures over 30 °C, as observed during the week 4 of the experimental period in winter and summer (
Table 2), could have led to thermal discomfort. There was a rise in the H/L ratio after heat exposure over 6 h [
39]; the 11 h of sunlight exposure in our outdoor areas could have caused this response, especially in commercial broilers, which are less tolerant to heat stress [
13,
40].
Decreased spleen and bursa RW observed indoors in summer, as well as decreased bursa RW in high-density treatments, are in accordance with the H/L results discussed above and with previous studies [
7,
8], which confirm that decreased RW of lymphoid organs is associated with chronically elevated corticosterone concentrations. This elicits the involution of lymphoid organs, such as the spleen and bursa of Fabricius, by the depletion of lymphocytes from germinal cells with dysregulated immune responses [
7,
41,
42]. However, other researchers have found decreased weight of the bursa of Fabricius as a result of heat stress [
41]. On the other hand, an increased bursa of Fabricius RW, as observed in winter and at low densities, could indicate the maturation of a greater number of T lymphocytes and B lymphocytes, since the bursa provides a suitable microenvironment for the proliferation and differentiation of these cells [
42,
43]. The larger the spleen, the stronger the immune system. However, the avian spleen size cannot always be used as an indicator of optimal immune status, as it has large seasonal intra-individual variations [
44].
The decreased resting among birds with outdoor access may have been responsible for their different tibia shape. The tibiae were wider and tended to be shorter, suggesting greater activity. Physical activity increases the diameter of the tibiotarsus diaphysis [
45]. This may be beneficial for tibiae strength, with breakages reflecting a potential problem when birds are caught. The better tibia bone properties observed in the LI treatment confirm that low stocking densities promoted behaviors such as walking and running (locomotion), which increased both the thickness and density of the cortical bone, as well as the diameter of the diaphysis [
45], as indicated by the higher Seedor index at low densities in winter. However, as physical activity tends to decrease with age this study and [
28], in combination with the consequences of the fast growth rate in broilers [
23], tibia bone properties could also be reduced over time.






{kind=link}