
CYP19A1 May Influence Lambing Traits in Goats by Regulating the Biological Function of Granulosa Cells



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Sourcing, Cell Collection, and Cell Identification
2.2. Plasmid Construction
2.3. Cell Culture and Transfection
2.4. Total RNA Extraction and Reverse Transcription
2.5. Real-Time PCR
2.6. Cell Proliferation Assay
2.7. Steroid Assay
2.8. Immunohistochemistry
2.9. Statistical Analysis
3. Results
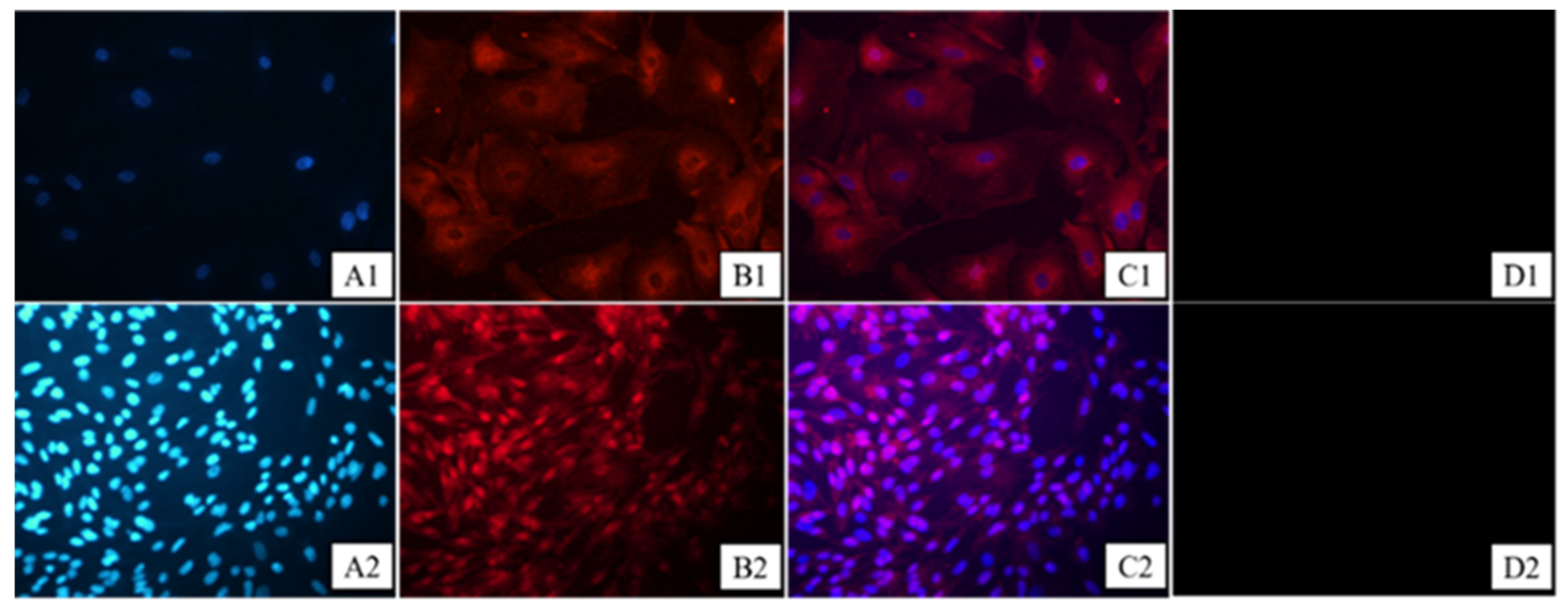
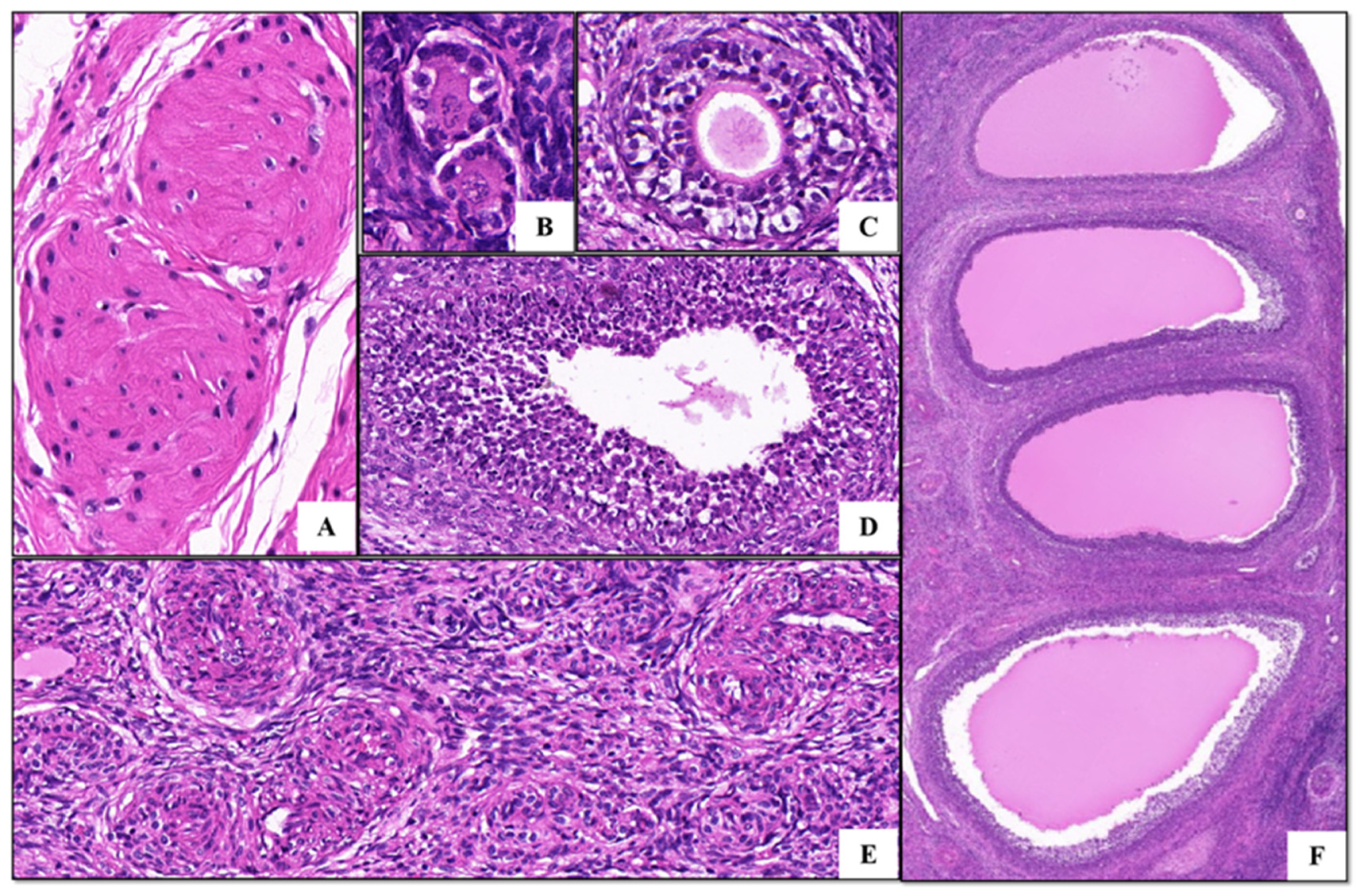
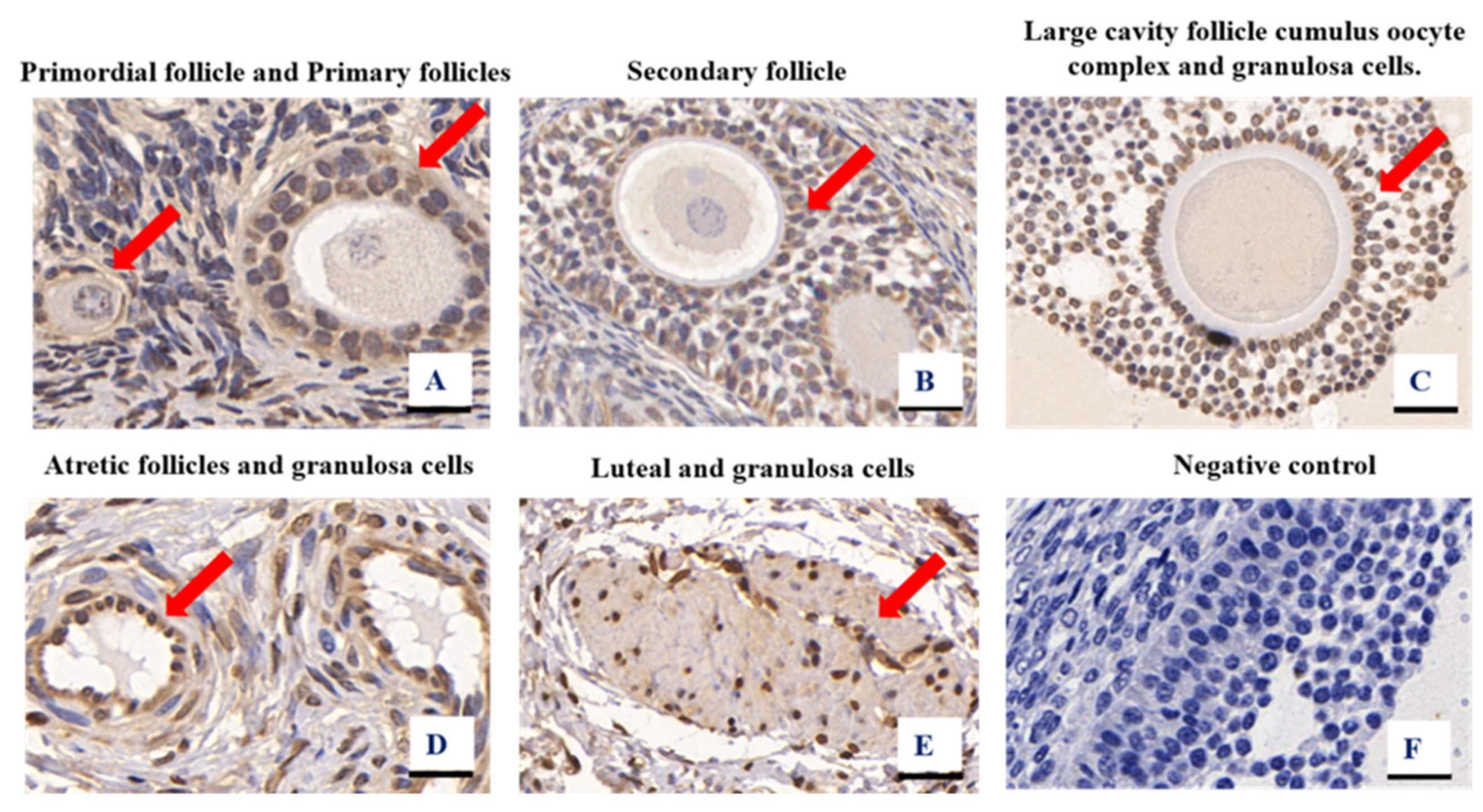
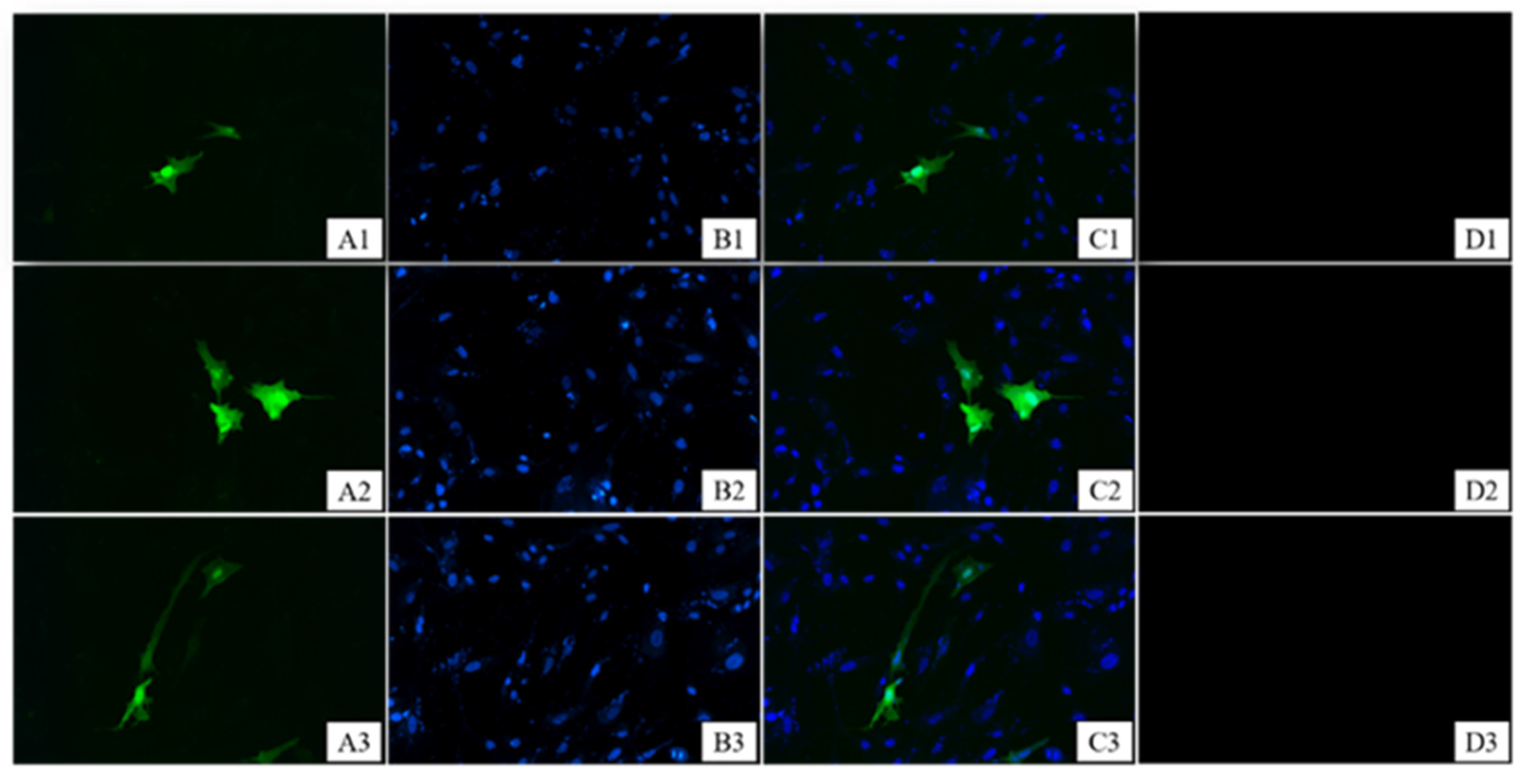
3.1. Granulosa Cell Identification and HE Staining of Ovarian Tissues
3.2. Figures, Tables, and Schemes
3.3. Effects of Overexpression and Silencing of CYP19A1 on the mRNA Expressions of BMPR-IB, FSHR, and INHBA in Granulosa Cells
3.4. Effects of CYP19A1 on the Proliferation of Goat Granulosa Cells
3.5. Effects of CYP19A1 on the Secretion of Steroid Hormones from Goat Granulosa Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tajima, K.; Orisaka, M.; Yata, H.; Goto, K.; Hosokawa, K.; Kotsuji, F. Role of granulosa and theca cell interactions in ovarian follicular maturation. Microsc. Res. Tech. 2006, 69, 450–458. [Google Scholar] [CrossRef]
- Vanderhyden, B.C.; Telfer, E.E.; Eppig, J.J. Mouse oocytes promote proliferation of granulosa cells from preantral and antral follicles in vitro. Biol. Reprod. 1992, 46, 1196–1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havelock, J.C.; Rainey, W.E.; Carr, B.R. Ovarian granulosa cell lines. Mol. Cell. Endocrinol. 2004, 228, 67–78. [Google Scholar] [CrossRef]
- Sanchez, A.M.; Somigliana, E.; Vercellini, P.; Pagliardini, L.; Candiani, M.; Vigano, P. Endometriosis as a detrimental condition for granulosa cell steroidogenesis and development: From molecular alterations to clinical impact. J. Steroid. Biochem. Mol. Biol. 2016, 155, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.E. The role of steroids in follicular growth. Reprod. Biol. Endocrinol. 2006, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Jain, P.; Jain, M.; Haldar, C.; Singh, T.B.; Jain, S. Melatonin and its correlation with testosterone in polycystic ovarian syndrome. J. Hum. Reprod. Sci. 2013, 6, 253–258. [Google Scholar] [CrossRef]
- Mahmoodi, M.; Soleimani Mehranjani, M.; Shariatzadeh, S.M.; Eimani, H.; Shahverdi, A. N-acetylcysteine improves function and follicular survival in mice ovarian grafts through inhibition of oxidative stress. Reprod. Biomed. Online 2015, 30, 101–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macklon, N.S.; Fauser, B.C. Aspects of ovarian follicle development throughout life. Horm. Res. 1999, 52, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, Y.; Zhao, C.; Yu, T.; Liu, Y.; Shi, W.; Shi, F.; Liu, X.; Sheng, J.; Huang, H.; et al. Reduced alternative splicing of estrogen receptor alpha in the endometrium of women with endometriosis. Oncotarget 2017, 8, 110176–110186. [Google Scholar] [CrossRef] [Green Version]
- Siquara De Sousa, A.C.; Capek, S.; Amrami, K.K.; Spinner, R.J. Neural involvement in endometriosis: Review of anatomic distribution and mechanisms. Clin. Anat. 2015, 28, 1029–1038. [Google Scholar] [CrossRef]
- Zhang, W.; Lu, H.; Jiang, H.; Li, M.; Zhang, S.; Liu, Q.; Zhang, L. Isolation and characterization of cyp19a1a and cyp19a1b promoters in the protogynous hermaphrodite orange-spotted grouper (Epinephelus coioides). Gen. Comp. Endocrinol. 2012, 175, 473–487. [Google Scholar] [CrossRef]
- Vega, W.H.O.; Quirino, C.R.; Bartholazzi-Junior, A.; Rua, M.A.S.; Serapiao, R.V.; Oliveira, C.S. Variants in the CYP19A1 gene can affect in vitro embryo production traits in cattle. J. Assist. Reprod. Genet. 2018, 35, 2233–2241. [Google Scholar] [CrossRef]
- Bao, B.; Garverick, H.A. Expression of steroidogenic enzyme and gonadotropin receptor genes in bovine follicles during ovarian follicular waves: A review. J. Anim. Sci. 1998, 76, 1903–1921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juengel, J.L.; Hudson, N.L.; Heath, D.A.; Smith, P.; Reader, K.L.; Lawrence, S.B.; O’Connell, A.R.; Laitinen, M.P.; Cranfield, M.; Groome, N.P.; et al. Growth differentiation factor 9 and bone morphogenetic protein 15 are essential for ovarian follicular development in sheep. Biol. Reprod. 2002, 67, 1777–1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, J.; McAndrew, B.; Penman, D. Cloning of brain aromatase gene and expression of brain and ovarian aromatase genes during sexual differentiation in genetic male and female Nile tilapia Oreochromis niloticus. Mol. Reprod. Dev. 2001, 59, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Hess, M.F.; Roser, J.F. Immunocytochemical localization of cytochrome P450 aromatase in the testis of prepubertal, pubertal, and postpubertal horses. Theriogenology 2004, 61, 293–299. [Google Scholar] [CrossRef]
- Albrecht, B.A.; Daels, P.F. Immunolocalization of 3 beta-hydroxysteroid dehydrogenase, cytochrome P450 17 alpha-hydroxylase/17,20-lyase and cytochrome P450 aromatase in the equine corpus luteum of dioestrus and early pregnancy. J. Reprod. Fertil. 1997, 111, 127–133. [Google Scholar] [CrossRef] [Green Version]
- Zhou, R.; Yang, Y.-l.; Liu, Y.; Chen, Q.-M.; Chen, J.; Li, K. Association of CYP19A1 gene polymorphisms with reproductive traits in pigs. J. Integr. Agric. 2017, 16, 1558–1565. [Google Scholar] [CrossRef]
- Logan, K.A.; Juengel, J.L.; McNatty, K.P. Onset of steroidogenic enzyme gene expression during ovarian follicular development in sheep. Biol. Reprod. 2002, 66, 906–916. [Google Scholar] [CrossRef] [Green Version]
- Quirke, L.D.; Juengel, J.L.; Tisdall, D.J.; Lun, S.; Heath, D.A.; McNatty, K.P. Ontogeny of steroidogenesis in the fetal sheep gonad. Biol. Reprod. 2001, 65, 216–228. [Google Scholar] [CrossRef]
- Luo, M.; Li, L.; Xiao, C.; Sun, Y.; Wang, G.L. Heat stress impairs mice granulosa cell function by diminishing steroids production and inducing apoptosis. Mol. Cell. Biochem. 2016, 412, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, V.; Salvetti, N.R.; Matiller, V.; Ortega, H.H. Developmental programming: Prenatal steroid excess disrupts key members of intraovarian steroidogenic pathway in sheep. Endocrinology 2014, 155, 3649–3660. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Wang, X.; Gao, F.; Du, W. Cyp19a1Role of in the female pathway of a freshwater turtle species (Mauremys reevesii) with temperature-dependent sex determination. Zool. Res. 2022, 43, 81–84. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Chen, X.; Zhu, M.; Wang, W.; Ao, Z.; Zhao, J.; Tang, W.; Hong, L. Cathepsin D knockdown regulates biological behaviors of granulosa cells and affects litter size traits in goats. J. Zhejiang Univ. Sci. B 2021, 22, 893–905. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.Y.; Gao, K.X.; Xin, H.Y.; Han, P.; Zhu, G.Q.; Cao, B.Y. Molecular cloning, expression analysis, and function of decorin in goat ovarian granulosa cells. Domest. Anim. Endocrinol. 2016, 57, 108–116. [Google Scholar] [CrossRef]
- Gong, T.; Wang, W.; Xu, H.; Yang, Y.; Chen, X.; Meng, L.; Xu, Y.; Li, Z.; Wan, S.; Mu, Q. Longitudinal Expression of Testicular TAS1R3 from Prepuberty to Sexual Maturity in Congjiang Xiang Pigs. Animals 2021, 11, 437. [Google Scholar] [CrossRef]
- Storbeck, K.H.; Swart, A.C.; Slabbert, J.T.; Swart, P. The identification of two CYP17 alleles in the South African Angora goat. Drug. Metab. Rev. 2007, 39, 467–480. [Google Scholar] [CrossRef]
- Storbeck, K.H.; Swart, A.C.; Snyman, M.A.; Swart, P. Two CYP17 genes in the South African Angora goat (Capra hircus)—The identification of three genotypes that differ in copy number and steroidogenic output. FEBS J. 2008, 275, 3934–3943. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Cheng, L.; Li, X.; Tang, S.; Zhang, X.; Gong, Y. Transcriptional regulation of CYP19A1 expression in chickens: ESR1, ESR2 and NR5A2 form a functional network. GEN. Comp. Endocr. 2022, 315, 113939. [Google Scholar] [CrossRef]
- Chou, C.H.; Chen, M.J. The Effect of Steroid Hormones on Ovarian Follicle Development. Vitam. Horm. 2018, 107, 155–175. [Google Scholar] [CrossRef]
- Strzalka, W.; Ziemienowicz, A. Proliferating cell nuclear antigen (PCNA): A key factor in DNA replication and cell cycle regulation. Ann. Bot. 2011, 107, 1127–1140. [Google Scholar] [CrossRef] [Green Version]
- Pradeep, P.K.; Li, X.; Peegel, H.; Menon, K.M. Dihydrotestosterone inhibits granulosa cell proliferation by decreasing the cyclin D2 mRNA expression and cell cycle arrest at G1 phase. Endocrinology 2002, 143, 2930–2935. [Google Scholar] [CrossRef]
- Shimizu, K.; Nakamura, T.; Bayasula; Nakanishi, N.; Kasahara, Y.; Nagai, T.; Murase, T.; Osuka, S.; Goto, M.; Iwase, A.; et al. Molecular mechanism of FSHR expression induced by BMP15 in human granulosa cells. J. Assist. Reprod. Genet. 2019, 36, 1185–1194. [Google Scholar] [CrossRef]
- Zi, X.D.; Xu, H.W.; Wang, Y. Variation in sequences and mRNA expression levels of inhibin subunits alpha (INHA) and betaA (INHBA) genes between prolific and nonprolific goat breeds. Mol. Reprod. Dev. 2012, 79, 238. [Google Scholar] [CrossRef]
- Chu, M.X.; Guo, X.H.; Feng, C.J.; Li, Y.; Huang, D.W.; Feng, T.; Cao, G.L.; Fang, L.; Di, R.; Tang, Q.Q.; et al. Polymorphism of 5′ regulatory region of ovine FSHR gene and its association with litter size in Small Tail Han sheep. Mol. Biol. Rep. 2012, 39, 3721–3725. [Google Scholar] [CrossRef]
- Xia, Y.; Wang, Q.; He, X.D.; Chen, Y.; JiGe, M.T.; Zi, X.D. Cloning and expression analysis of the follicle-stimulating hormone receptor (FSHR) gene in the reproductive axis of female yaks (Bos grunniens). Domest. Anim. Endocrinol. 2020, 70, 106383. [Google Scholar] [CrossRef]
- Saint-Dizier, M.; Malandain, E.; Thoumire, S.; Remy, B.; Chastant-Maillard, S. Expression of follicle stimulating hormone and luteinizing hormone receptors during follicular growth in the domestic cat ovary. Mol. Reprod. Dev. 2007, 74, 989–996. [Google Scholar] [CrossRef]
- Silva, J.R.; van den Hurk, R.; van Tol, H.T.; Roelen, B.A.; Figueiredo, J.R. Gene expression and protein localisation for activin-A, follistatin and activin receptors in goat ovaries. J. Endocrinol. 2004, 183, 405–415. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Wang, J.; Zhao, Y.; Ma, D.; Zhao, M.; Li, N.; Men, Y.; Zhang, Y.; Chu, H.; Lei, C.; et al. scRNA-seq of ovarian follicle granulosa cells from different fertility goats reveals distinct expression patterns. Reprod. Domest. Anim. 2021, 56, 801–811. [Google Scholar] [CrossRef]
- Klymiuk, M.C.; Neunzig, J.; Bernhardt, R.; Sanchez-Guijo, A.; Hartmann, M.F.; Wudy, S.A.; Schuler, G. Efficiency of the sulfate pathway in comparison to the Delta4- and Delta5-pathway of steroidogenesis in the porcine testis. J. Steroid. Biochem. Mol. Biol. 2018, 179, 64–72. [Google Scholar] [CrossRef]
- Li, Q.; Du, X.; Liu, L.; Liu, H.; Pan, Z.; Li, Q. Upregulation of miR-146b promotes porcine ovarian granulosa cell apoptosis by attenuating CYP19A1. Domest. Anim. Endocrinol. 2021, 74, 106509. [Google Scholar] [CrossRef]
- Miller, W.L. Androgen biosynthesis from cholesterol to DHEA. Mol. Cell. Endocrinol. 2002, 198, 7–14. [Google Scholar] [CrossRef]
- Miller, W.L. Disorders in the initial steps of steroid hormone synthesis. J. Steroid. Biochem. Mol. Biol. 2017, 165, 18–37. [Google Scholar] [CrossRef]
- Feuerstein, P.; Cadoret, V.; Dalbies-Tran, R.; Guerif, F.; Bidault, R.; Royere, D. Gene expression in human cumulus cells: One approach to oocyte competence. Hum. Reprod. 2007, 22, 3069–3077. [Google Scholar] [CrossRef] [Green Version]
- Hamel, M.; Dufort, I.; Robert, C.; Gravel, C.; Leveille, M.-C.; Leader, A.; Sirard, M.-A. Identification of differentially expressed markers in human follicular cells associated with competent oocytes. Hum. Reprod. 2008, 23, 1118–1127. [Google Scholar] [CrossRef] [Green Version]
- Wathlet, S.; Adriaenssens, T.; Segers, I.; Verheyen, G.; Janssens, R.; Coucke, W.; Devroey, P.; Smitz, J. New candidate genes to predict pregnancy outcome in single embryo transfer cycles when using cumulus cell gene expression. Fertil. Steril. 2012, 98, 432–439.e4. [Google Scholar] [CrossRef]
- Cui, J.; Shen, Y.; Li, R. Estrogen synthesis and signaling pathways during aging: From periphery to brain. Trends Mol. Med. 2013, 19, 197–209. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequences (5′-3′) |
|---|---|
| sh1-CYP19A1 | F: CACCGCGGTCACCAACATAATCAGCTTCAAGAGAGCTGATTATGTTGGTGACCGCTTTTTTG R: GATCCAAAAAAGCGGTCACCAACATAATCAGCTCTCTTGAAGCTGATTATGTTGGTGACCGC |
| sh2-CYP19A1 | F: CACCGACCAGAATATAGGTTTCAATTTCAAGAGAATTGAAACCTATATTCTGGTCTTTTTTG R: GATCCAAAAAAGACCAGAATATAGGTTTCAATTCTCTTGAAATTGAAACCTATATTCTGGTC |
| sh3-CYP19A1 | F: CACCGAGGCAATGATGAGGGAAATCTTCAAGAGAGATTTCCCTCATCATTGCCTCTTTTTTG R: GATCCAAAAAAGAGGCAATGATGAGGGAAATCTCTCTTGAAGATTTCCCTCATCATTGCCTC |
| sh4-CYP19A | F: CACCGCTGTGCAGAAAGTATGAAAATTCAAGAGATTTTCATACTTTCTGCACAGCTTTTTTG R: GATCCAAAAAAGCTGTGCAGAAAGTATGAAAATCTCTTGAATTTTCATACTTTCTGCACAGC |
| sh-NC | F: CACCGTTCTCCGAACGTGTCACGTTTCAAGAGAACGTGACACGTTCGGAGAATTTTTTG R: GATCCAAAAAATTCTCCGAACGTGTCACGTTCTCTTGAAACGTGACACGTTCGGAGAAC |
| pEGFP-N3-CYP19A1 | (BglII) F: CGTCAGATCCGCTAGCGCTACCGGACTCAGATCTATGCTTTTGGAAGTGCTGAAC (SacIIR): CACCATGGTGGCGATGGATCCCGGGCCCGCGGCGCACTCGAGGCACTTGTCTGAATTTCTC |
| Gene Name | Primer Sequences (5′-3′) | Login ID | Fragment Size (bp) |
|---|---|---|---|
| BMPR-1B | F: GCTCTTGGTCCTCATCATTTTATTC R: ATGTAAGTTTCGTCCTGTTCTAACC | NC_022298.2 | 143 |
| FSHR | F: TGTTATGTCCCTCCTTGTGCTC R: CGCTTGGCTATCTTGGTGTCA | NM_001285644.1 | 122 |
| PCNA | F: GTAGCCGTGTCATTGCGACTCC R: GCTCTGTAGGTTCACGCCACTTG | XM_005688167.3 | 145 |
| INHBA | F: AAGGTGGTGGATGCTCGAAA R: GTCTCCTGACACTGCTCACA | NM_001285581.1 | 125 |
| CYP19A1 | F: CCCAAGGCATTACAATGT R: TAAGGGTTTCCTCTCCAC | NM_001314145.1 | 169 |
| STAR | F: GGTCCCCGAGACTTTGTGAG R: AATCCACTTGGGTCTGCGAG | XM_013975437.2 | 262 |
| 3βHSD | F: AGACCAGAAGTTCGGGAGGAA R: TCTCCCTGTAGGAGTTGGGC | NM_001285716.1 | 292 |
| CYP11A1 | F: CTCCAGAGGCAATAAAGAA R: TCAAAGGCAAAGTGAAACA | NM_001287574.1 | 145 |
| Cyclin E | F: GATGTCGGCTGCTTAGAAT R: CACCACTGATACCCTGAAAC | XM_018062248.1 | 104 |
| β-actin | F: AGATGTGGATCAGCAAGCAG R: CCAATCTCATCTCGTTTTCTG | NM_001297986.1 | 139 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Chen, X.; Zhou, Z.; Tian, X.; Yang, P.; Fu, K. CYP19A1 May Influence Lambing Traits in Goats by Regulating the Biological Function of Granulosa Cells. Animals 2022, 12, 1911. https://doi.org/10.3390/ani12151911
Zhang Y, Chen X, Zhou Z, Tian X, Yang P, Fu K. CYP19A1 May Influence Lambing Traits in Goats by Regulating the Biological Function of Granulosa Cells. Animals. 2022; 12(15):1911. https://doi.org/10.3390/ani12151911
Chicago/Turabian StyleZhang, Yan, Xiang Chen, Zhinan Zhou, Xingzhou Tian, Peifang Yang, and Kaibing Fu. 2022. "CYP19A1 May Influence Lambing Traits in Goats by Regulating the Biological Function of Granulosa Cells" Animals 12, no. 15: 1911. https://doi.org/10.3390/ani12151911