
Physicochemical Analysis of Mixed Venous and Arterial Blood Acid-Base State in Horses at Core Temperature during and after Moderate-Intensity Exercise


Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods
2.1. Ethical Approval
2.2. Animals
2.3. Exercise Protocol
2.4. Analysis
2.5. Calculations
2.6. Statistics
3. Results
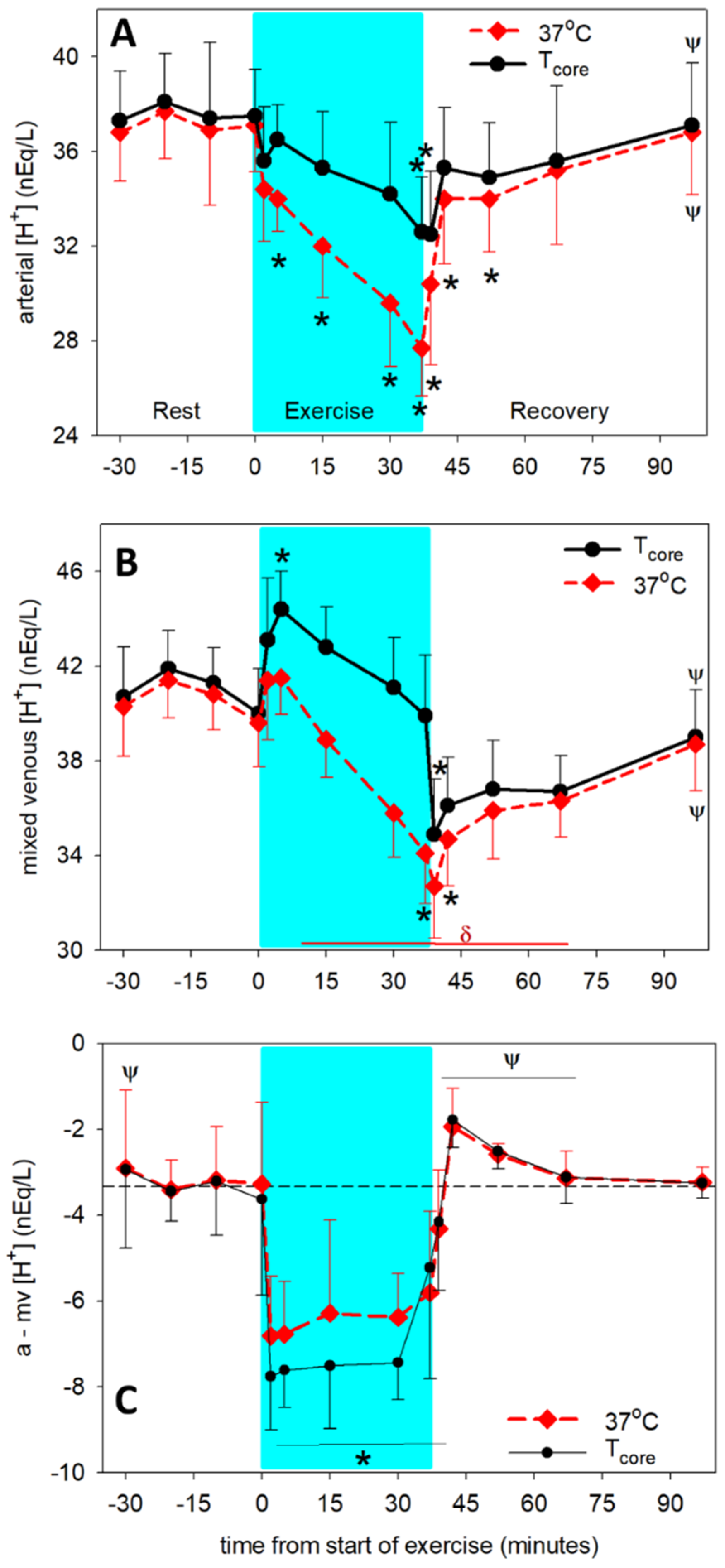
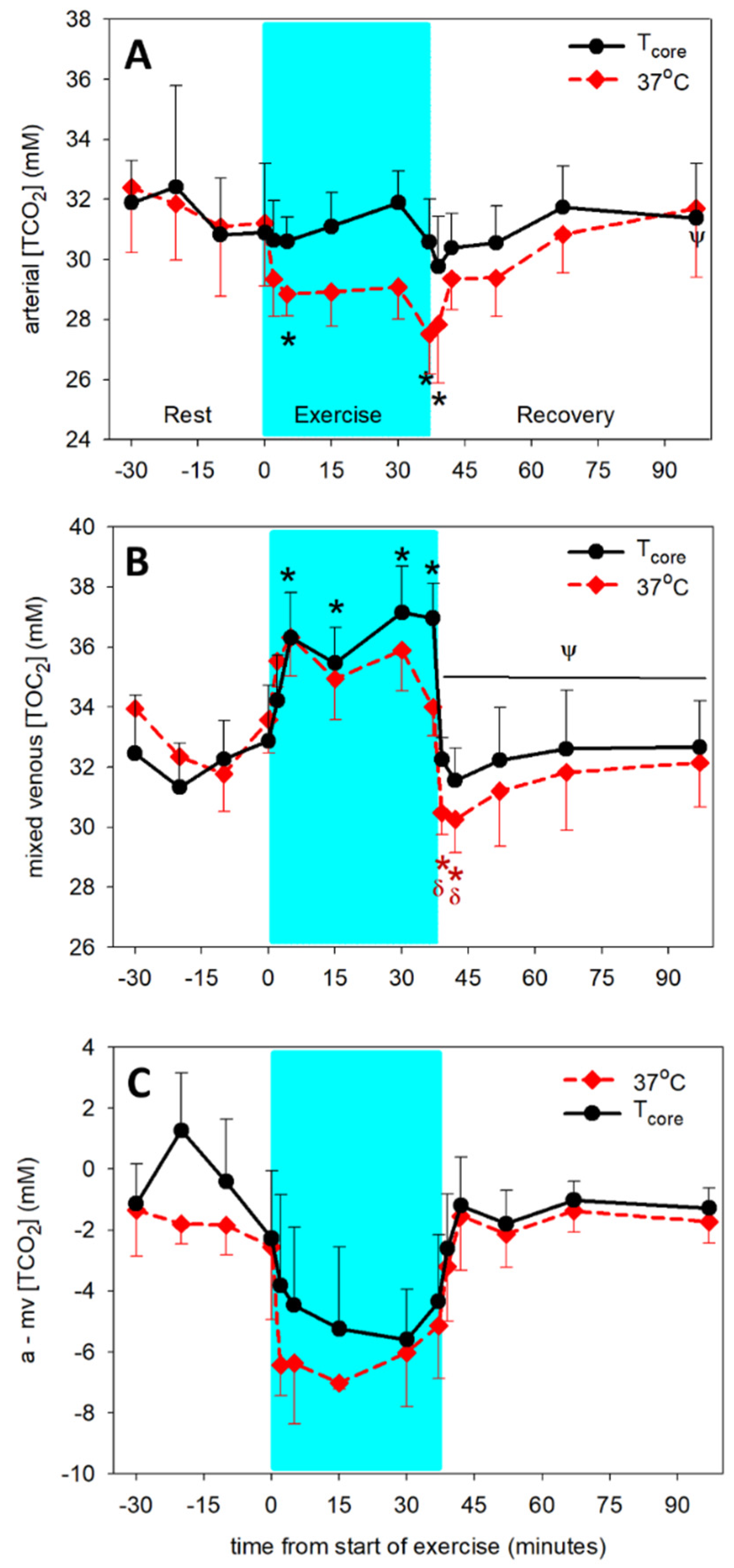
3.1. Temperature-Dependent Acid-Base Variables
3.2. Temperature-Independent Acid-Base Variables
3.3. Determining the Origins of the Changes in Dependent Acid-Base Variables
4. Discussion
4.1. Temperature Adjustment
4.2. Gas Exchange
4.3. Factors Contributing to the Acid-Base Changes of Moderate-Intensity Exercise
5. Limitations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arfuso, F.; Assenza, A.; Fazio, F.; Rizzo, M.; Giannetto, C.; Piccione, G. Dynamic Change of Serum Levels of Some Branched-Chain Amino Acids and Tryptophan in Athletic Horses After Different Physical Exercises. J. Equine Vet. Sci. 2019, 77, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Arfuso, F.; Giudice, E.; Panzera, M.; Rizzo, M.; Fazio, F.; Piccione, G.; Giannetto, C. Interleukin-1Ra (Il-1Ra) and Serum Cortisol Level Relationship in Horse as Dynamic Adaptive Response during Physical Exercise. Vet. Immunol. Immunopathol. 2022, 243, 110368. [Google Scholar] [CrossRef] [PubMed]
- Westerblad, H.; Allen, D.G.; Lännergren, J. Muscle Fatigue: Lactic Acid or Inorganic Phosphate the Major Cause? News Physiol. Sci. 2002, 17, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Meyer, N.D.; Bayly, W.M.; Sides, R.H.; Wardrop, K.J.; Slinker, B.K. Changes in Arterial, Mixed Venous and Intraerythrocytic Ion Concentrations during Prolonged Exercise. Equine Vet. J. 2010, 42, 185–190. [Google Scholar] [CrossRef]
- Kupczyński, R.; Śpitalniak, K. Analysis of Acid-Base Balance as Well as Hematological and Biochemical Parameters in Horses of Combined Driving Discipline. Arch. Anim. Breed. 2015, 58, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Arfuso, F.; Giannetto, C.; Giudice, E.; Fazio, F.; Panzera, F.; Piccione, G. Venous Blood Acid-Base Status in Show Jumper Horses Subjected to Different Physical Exercises. J. Equine Vet. Sci. 2020, 94, 103251. [Google Scholar] [CrossRef] [PubMed]
- Tavares Miranda, A.C.; Padilha, F.G.F.; Ramos, M.T.; Dimache, L.A.G.; Nascimento Godoi, F.; Galina Filho, A.; de Almeida, F.Q. Hemogasometry of Eventing Horses before and after Intense Exercise on a High Speed Treadmill. Livest. Sci. 2016, 186, 85–87. [Google Scholar] [CrossRef]
- Kingston, J.K.; Bayly, W.M. Effect of Exercise on Acid-Base Status of Horses. Vet. Clin. N. Am. Equine Pract. 1998, 14, 61–73. [Google Scholar] [CrossRef]
- Vengust, M.; Kerr, C.; Staempfli, H.R.; Pringle, J.; Heigenhauser, G.J.; Viel, L. Effect of Frusemide on Transvascular Fluid Fluxes across the Lung in Exercising Horses. Equine Vet. J. 2011, 43, 451–459. [Google Scholar] [CrossRef]
- Vengust, M.; Stämpfli, H.; de Moraes, A.N.; Teixeiro-Neto, F.; Viel, L.; Heigenhauser, G. Effects of Chronic Acetazolamide Administration on Gas Exchange and Acid-Base Control in Pulmonary Circulation in Exercising Horses. Equine Vet. J. 2010, 42, 40–50. [Google Scholar] [CrossRef]
- Kronfeld, D.S.; Ferrante, P.L.; Taylor, L.E.; Tiegs, W. Partition of Plasma Hydrogen Ion Concentration Changes during Repeated Sprints. Equine Vet. J. Suppl. 1999, 30, 380–383. [Google Scholar] [CrossRef] [PubMed]
- Viu, J.; Jose-Cunilleras, E.; Armengou, L.; Cesarini, C.; Tarancón, I.; Rios, J.; Monreal, L. Acid-Base Imbalances during a 120 Km Endurance Race Compared by Traditional and Simplified Strong Ion Difference Methods. Equine Vet. J. 2010, 42, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Kirsch, K.; Sandersen, C. Traditional and Quantitative Analysis of Acid-Base and Electrolyte Imbalances in Horses Competing in Cross-Country Competitions at 2-Star to 5-Star Level. J. Vet. Intern. Med. 2020, 34, 909–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, G.; Fuller, A.; Maloney, S.K.; Rump, N.; Mitchell, D. Guttural Pouches, Brain Temperature and Exercise in Horses. Biol. Lett. 2006, 2, 475–477. [Google Scholar] [CrossRef]
- Gunn, H.M.M. Muscle, Bone and Fat Proportions and Muscle Distribution of Thoroughbreds and Other Horses. In Equine Exercise Physiology; ICEEP Publications: Davis, CA, USA, 1987; Vol. n/a, pp. 253–264. [Google Scholar]
- Sheel, A.W.; Boushel, R.; Dempsey, J.A. Competition for Blood Flow Distribution between Respiratory and Locomotor Muscles: Implications for Muscle Fatigue. J. Appl. Physiol. 2018, 125, 820–831. [Google Scholar] [CrossRef]
- Laughlin, H.M.; Davis, M.J.; Secher, N.H.; van Lieshout, J.J.; Arce-Esquivel, A.A.; Simmons, G.H.; Bender, S.B.; Padilla, J.; Bache, R.J.; Merkus, D.; et al. Peripheral Circulation. Compr. Physiol. 2012, 2, 321–447. [Google Scholar] [CrossRef]
- Jones, J.H.; Taylor, C.R.; Lindholm, A.; Straub, R.; Longworth, K.E.; Karas, R.H. Blood Gas Measurements during Exercise: Errors Due to Temperature Correction. J. Appl. Physiol. 1989, 67, 879–884. [Google Scholar] [CrossRef]
- Constable, P.D. Clinical Assessment of Acid-Base Status. Strong Ion Difference Theory. Vet. Clin. N. Am. Food Anim. Pract. 1999, 15, 447–471. [Google Scholar] [CrossRef]
- Waller, A.; Heigenhauser, G.J.; Lindinger, M.I. Electrolyte Supplementation after Prolonged Moderate-Intensity Exercise Results in Decreased Plasma [TCO2] in Standardbreds. Equine Comp. Exerc. Physiol. 2007, 4, 149–158. [Google Scholar] [CrossRef]
- Henderson, L.J. Acidosis: I. Science 1917, 46, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Singer, R.B.; Hastings, A.B. An Improved Clinical Method for the Estimation of Disturbances of the Acid-Base Balance of Human Blood. Medicine 1948, 27, 223–242. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.A. Independent and Dependent Variables of Acid-Base Control. Respir. Physiol. 1978, 33, 9–26. [Google Scholar] [CrossRef]
- Stewart, P.A. Modern Quantitative Acid-Base Chemistry. Can. J. Physiol. Pharmacol. 1983, 61, 1444–1461. [Google Scholar] [CrossRef] [PubMed]
- Waller, A.; Lindinger, M.I. The Effect of Oral Sodium Acetate Administration on Plasma Acetate Concentration and Acid-Base State in Horses. Acta Vet. Scand. 2007, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constable, P.D. The Physicochemical Approach for Evaluating Acid-Base Balance in Exercising Horses. Equine Vet. J. 1999, 31, 636–638. [Google Scholar] [CrossRef]
- Stickland, M.K.; Lindinger, M.I.; Olfert, I.M.; Heigenhauser, G.J.F.; Hopkins, S.R. Pulmonary Gas Exchange and Acid-Base Balance during Exercise. Compr. Physiol. 2013, 3, 693–739. [Google Scholar] [CrossRef]
- Lindinger, M.I.; Heigenhauser, G.J.F. Effects of Gas Exchange on Acid-Base Balance. Compr. Physiol. 2012, 2, 2203–2254. [Google Scholar] [CrossRef]
- Malan, A. Blood Acid-Base State at a Variable Temperature. a Graphical Representation. Respir. Physiol. 1977, 31, 259–275. [Google Scholar] [CrossRef]
- Reeves, R.B. Temperature Induced Changes in Blood Acid Base Status: PH and Pco2 in a Binary Buffer. J. Appl. Physiol. 1976, 40, 752–761. [Google Scholar] [CrossRef]
- Ashwood, E.R.; Kost, G.; Kenny, M. Temperature Correction of Blood-Gas and PH Measurements. Clin. Chem. 1983, 29, 1877–1885. [Google Scholar] [CrossRef]
- Malan, A. The Evolution of Mammalian Hibernation: Lessons from Comparative Acid-Base Physiology. Integr. Comp. Biol. 2014, 54, 484–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, L.E.; Kronfeld, D.S.; Ferrante, P.L.; Wilson, J.A.; Tiegs, W. Blood-Gas Measurements Adjusted for Temperature at Three Sites during Incremental Exercise in the Horse. J. Appl. Physiol. 1998, 85, 1030–1036. [Google Scholar] [CrossRef] [PubMed]
- Forster, H.V.; Murphy, C.L.; Brice, A.G.; Pan, L.G.; Lowry, T.F. In Vivo Regulation of Plasma [H+] in Ponies during Acute Changes in PCO2. J. Appl. Physiol. 1990, 68, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Lindinger, M.I.; Geor, R.J.; Ecker, G.L.; McCutcheon, L.J. Plasma Volume and Ions during Exercise in Cool, Dry; Hot, Dry; and Hot, Humid Conditions. Equine Vet. J. 1995, 27, 133–139. [Google Scholar] [CrossRef]
- McCutcheon, L.J.; Geor, R.J.; Hare, M.J.; Ecker, G.L.; Lindinger, M.I. Sweating Rate and Sweat Composition during Exercise and Recovery in Ambient Heat and Humidity. Equine Vet. J. 1995, 27, 153–157. [Google Scholar] [CrossRef]
- Geor, R.J.; McCutcheon, L.J.; Ecker, G.L.; Lindinger, M.I. Thermal and Cardiorespiratory Responses of Horses to Submaximal Exercise under Hot and Humid Conditions. Equine Vet. J. 1995, 27, 125–132. [Google Scholar] [CrossRef]
- Lindinger, M.I.; Heigenhauser, G.J.F.; McKelvie, R.S.; Jones, N.L. Blood Ion Regulation during Repeated Maximal Exercise and Recovery in Humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1992, 262. [Google Scholar] [CrossRef]
- Waller, A.; Lindinger, M.I. Physicochemical Analysis of Acid–Base Status during Recovery from High-Intensity Exercise in Standardbred Racehorses. Equine Comp. Exerc. Physiol. 2005, 2, 119–127. [Google Scholar] [CrossRef]
- Siggaard-Andersen, O.; Wimberley, P.D.; Gøthgen, I.H.; Fogh-Andersen, N.; Rasmussen, J.P. Variability of the Temperature Coefficients for Ph, PCO2, and Po2 in Blood. Scand. J. Clin. Lab. Investig. 1988, 48, 85–88. [Google Scholar] [CrossRef]
- Austin, W.H.; Lacombe, E.; Rand, P.W.; Chaterjee, M. Solubility of Carbon Dioxide in Serum from 15 to 38 C. J. Appl. Physiol. 1963, 18, 301–304. [Google Scholar] [CrossRef]
- Rispens, P.; Dellebarre, C.W.; Eleveld, D.; Helder, W.; Zijlstra, W.G. The Apparent First Dissociation Constant of Carbonic Acid in Plasma between 16 and 42.5°. Clin. Chim. Acta 1968, 22, 627–637. [Google Scholar] [CrossRef]
- Burnett, R.W.; Noonan, D.C. Calculations and Correction Factors Used in Determination of Blood PH and Blood Gases. Clin. Chem. 1974, 20, 1499–1506. [Google Scholar] [CrossRef] [PubMed]
- Beetham, R. A Review of Blood PH and Blood-Gas Analysis. Ann. Clin. Biochem. 1982, 19, 198–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constable, P.D. A Simplified Strong Ion Model for Acid-Base Equilibria: Application to Horse Plasma. J. Appl. Physiol. 1997, 83, 297–311. [Google Scholar] [CrossRef] [Green Version]
- Staempfli, H.R.; Constable, P.D. Experimental Determination of Net Protein Charge and Atot and Ka of Nonvolatile Buffers in Human Plasma. J. Appl. Physiol. 2003, 95, 620–630. [Google Scholar] [CrossRef] [Green Version]
- Forster, H.V.; Murphy, C.L.; Brice, A.G.; Pan, L.G.; Lowry, T.F. Plasma [H+] Regulation and Whole Blood [CO2] in Exercising Ponies. J. Appl. Physiol. 1990, 68, 309–315. [Google Scholar] [CrossRef]
- Foreman, J.H.; Grubb, T.L.; Benson, G.J.; Frey, L.P.; Foglia, R.A.; Griffin, R.L. Acid-Base and Electrolyte Effects of Shortening Steeplechase in a Three-Day-Event. Equine Vet. J. Suppl. 1996, 85–90. [Google Scholar] [CrossRef]
- Marlin, D.J.; Scott, C.M.; Schroter, R.C.; Harris, R.C.; Harris, P.A.; Roberts, C.A.; Mills, P.C. Physiological Responses of Horses to a Treadmill Simulated Speed and Endurance Test in High Heat and Humidity before and after Humid Heat Acclimation. Equine Vet. J. 1999, 31, 31–42. [Google Scholar] [CrossRef]
- Geor, R.J.; McCutcheon, L.J.; Lindinger, M.I. Adaptations to Daily Exercise in Hot and Humid Ambient Conditions in Trained Thoroughbred Horses. Equine Vet. J. Suppl. 1996, 63–68. [Google Scholar] [CrossRef]
- Geor, R.J.; McCutcheon, L.J.; Ecker, G.L.; Lindinger, M.I. Heat Storage in Horses during Submaximal Exercise before and after Humid Heat Acclimation. J. Appl. Physiol. 2000, 89, 2283–2293. [Google Scholar] [CrossRef] [Green Version]
- Burnett, R.W.; Covington, A.K.; Fogh-Andersen, N.; Külpmann, W.R.; Maas, A.H.; Müller-Plathe, O.; Siggaard-Andersen, O.; van Kessel, A.L.; Wimberley, P.D.; Zijlstra, W.G. International Federation of Clinical Chemistry (IFCC). Scientific Division. Committee on PH, Blood Gases and Electrolytes. Approved IFCC Recommendations on Whole Blood Sampling, Transport and Storage for Simultaneous Determination of PH, Blood Gases and e. Eur. J. Clin. Chem Clin. Biochem. 1995, 33, 247–253. [Google Scholar] [PubMed]
- Kruse, J.A. Calculation of Plasma Bicarbonate Concentration versus Measurement of Serum CO2 Content. PK´ Revisited. Clin. Intensive Care 1995, 6, 15–20. [Google Scholar] [CrossRef]
- Forster, H.V.; Pan, L.G.; Bisgard, G.E. Hyperpnea of Exercise at Various PIO2 in Normal and Carotid Body-Denervated Ponies. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1983, 54, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Forster, H.; Pan, L.; Bisgard, G.; Dorsey, S.; Britton, M. Temporal Pattern of Pulmonary Gas Exchange during Exercise in Ponies. J. Appl. Physiol. 1984, 57, 760–767. [Google Scholar] [CrossRef]
- Pan, L.; Forster, H.; Bisgard, G.; Murphys, C.; Lowry, T. Independence of Exercise Hyperpnea and Acidosis during High-Intensity Exercise in Ponies. J. Appl. Physiol. 1986, 60, 1016–1024. [Google Scholar] [CrossRef]
- Pan, L.; Forster, H.; Bisgard, G.; Dorsey, S.; Busch, M. Cardiodynamic Variables and Ventilation during Treadmill Exercise in Ponies. J. Appl. Physiol. 1984, 57, 753–759. [Google Scholar] [CrossRef]
- Forster, H.V.; Haouzi, P.; Dempsey, J.A. Control of Breathing during Exercise. Compr. Physiol. 2012, 2, 743–777. [Google Scholar] [CrossRef]
- Forster, H.V.; Pan, L.G. Contribution of Acid-Base Changes to Control of Breathing during Exercise. Can. J. Appl. Physiol. 1995, 20, 380–394. [Google Scholar] [CrossRef]
- Henderson, L.J. The Regulation of Neutrality in the Animal Body. Science 1913, 37, 389–395. [Google Scholar] [CrossRef] [Green Version]
- Waller, A.; Lindinger, M.I. Time Course and Magnitude of Fluid and Electrolyte Shifts during Recovery from High-Intensity Exercise in Standardbred Racehorses. Equine Comp. Exerc. Physiol. 2005, 2, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Waller, A.P.; Lindinger, M.I. Pre-Loading Large Volume Oral Electrolytes: Tracing Fluid and Ion Fluxes in Horses during Rest, Exercise and Recovery. J. Physiol. 2021, 599, 3879–3896. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.G.; Forster, H.V.; Bisgard, G.E. O2 Transport in Ponies during Treadmill Exercise. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1984, 57, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Parks, C.M.; Manohar, M. Blood-Gas Tensions and Acid-Base Status in Ponies during Treadmill Exercise. Am. J. Vet. Res. 1984, 45, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Littlejohn, A.; Snow, D.H. Circulatory, Respiratory and Metabolic Responses in Thoroughbred Horses during the First 400 Meters of Exercise. Eur. J. Appl. Physiol. Occup. Physiol. 1988, 58, 307–314. [Google Scholar] [CrossRef]
- Lindinger, M.I. Total Carbon Dioxide in Adult Standardbred and Thoroughbred Horses. J. Equine Vet. Sci. 2021, 106, 103730. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Arterial | pHmeas Mean | SD | pHadj Mean | SD | pHcalc Mean | SD | [HCO3−]m Mean | SD | [HCO3-]adj Mean | SD | [HCO3−]calc Mean | SD | [TP] Mean | SD | HCT (%) Mean | SD |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| −30 | 7.435 | 0.0364 | 7.430 ** | 0.0364 | 7.403 | 0.0225 | 30.96 ** | 2.03 | 28.68 | 1.34 | 27.71 | 0.53 | 6.28 | 0.14 | 37.40 ** | 3.91 |
| −20 | 7.425 | 0.0239 | 7.420 ** | 0.0239 | 7.403 | 0.0609 | 30.47 ** | 1.81 | 29.17 | 2.19 | 28.78 | 1.40 | 6.16 | 0.15 | 34.50 ** | 5.28 |
| −10 | 7.434 | 0.0388 | 7.429 ** | 0.0388 | 7.417 | 0.0500 | 29.73 ** | 2.14 | 27.67 | 1.77 | 27.66 | 1.19 | 6.15 | 0.22 | 34.67 ** | 5.89 |
| 0 | 7.431 | 0.0233 | 7.426 ** | 0.0233 | 7.42 | 0.0379 | 29.83 | 2.02 | 27.74 | 2.17 | 28.12 | 1.90 | 6.28 | 0.16 | 38.67 | 5.09 |
| 2 | 7.464 | 0.0283 | 7.449 | 0.0283 | 7.439 | 0.0428 | 28.13 | 1.22 | 27.49 | 1.25 | 27.47 | 1.91 | 6.73 * | 0.36 | 49.5 * | 5.47 |
| 5 | 7.468 | 0.0182 | 7.438 | 0.0178 | 7.462 | 0.0462 | 27.67 | 0.74 | 27.47 | 0.76 | 29.60 | 3.06 | 6.54 | 0.31 | 51.3 * | 5.57 |
| 15 | 7.496 * | 0.0294 | 7.453 | 0.0289 | 7.462 | 0.0538 | 27.80 | 1.20 | 27.93 | 1.07 | 28.83 | 2.90 | 6.50 | 0.35 | 48.7 * | 3.78 |
| 30 | 7.53 * | 0.0387 | 7.468 * | 0.0377 | 7.46 | 0.0501 | 28.05 | 1.01 | 28.68 | 1.01 | 28.09 | 0.88 | 6.50 | 0.27 | 48.5 * | 3.94 |
| 37 | 7.559 * | 0.0317 | 7.488 * | 0.0309 | 7.486 | 0.0441 | 26.62 * | 1.34 | 27.44 | 1.35 | 27.11 | 1.21 | 6.55 | 0.40 | 47.8 * | 2.59 |
| 39 | 7.521 * | 0.0369 | 7.492 * | 0.0359 | 7.523 * | 0.0729 | 26.817 * | 1.77 | 26.66 | 1.58 | 29.17 | 4.67 | 6.61 | 0.33 | 48.5 * | 2.26 |
| 42 | 7.47 | 0.0318 | 7.453 | 0.0315 | 7.464 | 0.0429 | 27.83 | 1.17 | 27.26 | 1.08 | 28.59 | 2.10 | 6.52 | 0.31 | 46.0 * | 2.37 |
| 52 | 7.469 | 0.0296 | 7.458 | 0.0293 | 7.457 | 0.0359 | 28.17 | 1.20 | 27.41 | 1.16 | 27.98 | 2.12 | 6.30 | 0.26 | 41.50 ** | 2.88 |
| 67 | 7.455 | 0.0388 | 7.45 | 0.0288 | 7.419 | 0.0494 | 29.53 ** | 1.25 | 28.54 | 1.30 | 27.28 | 2.14 | 6.28 | 0.39 | 39.67 ** | 3.14 |
| 97 | 7.435 | 0.0319 | 7.432 ** | 0.0319 | 7.391 ** | 0.0278 | 30.32 ** | 2.14 | 28.19 | 1.72 | 26.43 | 1.85 | 6.34 | 0.19 | 37.83 ** | 3.92 |
| Venous | ||||||||||||||||
| −30 | 7.396 | 0.0348 | 7.391 | 0.0348 | 7.381 | 0.0352 | 32.63 | 1.46 | 29.22 ** | 2.77 | 29.44 | 2.73 | 6.38 | 0.25 | 36.50 ** | 4.04 |
| −20 | 7.383 | 0.0168 | 7.378 | 0.0168 | 7.405 | 0.0262 | 31.17 | 1.15 | 28.15 ** | 2.33 | 30.918 | 2.287 | 6.27 | 0.297 | 34.75 ** | 4.11 |
| −10 | 7.39 | 0.0157 | 7.385 | 0.0157 | 7.398 | 0.0194 | 30.6 | 1.06 | 29.03 | 1.21 | 31.017 | 2.234 | 6.41 | 0.23 | 38.25 ** | 5.19 |
| 0 | 7.403 | 0.0205 | 7.398 | 0.0205 | 7.393 | 0.0368 | 32.32 | 2.06 | 29.60 ** | 1.756 | 30.203 | 2.559 | 6.41 | 0.24 | 38.40 * | 5.08 |
| 2 | 7.383 | 0.0265 | 7.366 ** | 0.0265 | 7.394 | 0.0366 | 34.08 | 1.26 | 30.88 | 3.318 | 33.94 | 4.40 | 6.77 | 0.53 | 50.7 * | 5.79 |
| 5 | 7.382 | 0.0164 | 7.353 * | 0.0161 | 7.367 | 0.0464 | 32.9 | 2.41 | 32.86 | 2.359 | 34.91 | 3.77 | 6.83 | 0.46 | 50.6 * | 4.83 |
| 15 | 7.41 ** | 0.0178 | 7.369 ** | 0.0178 | 7.381 | 0.0533 | 33.7 | 2.52 | 32.06 | 3.015 | 33.76 | 5.01 | 6.77 | 0.49 | 47.8 * | 3.27 |
| 30 | 7.447 * | 0.023 | 7.386 | 0.0226 | 7.394 | 0.0366 | 34.68 | 1.45 | 33.65 * | 2.391 | 34.53 | 2.85 | 6.70 | 0.31 | 48.4 * | 2.70 |
| 37 | 7.469 * | 0.0424 | 7.401 | 0.0414 | 7.379 | 0.0272 | 32.64 | 0.97 | 33.47 * | 1.091 | 32.03 | 2.89 | 6.72 | 0.52 | 49.4 * | 2.51 |
| 39 | 7.487 * | 0.0304 | 7.458 * | 0.0301 | 7.447 | 0.0232 | 29.28 | 0.69 | 29.01 ** | 0.692 | 28.73 | 2.43 | 6.79 | 0.49 | 47.6 * | 2.61 |
| 42 | 7.46 * | 0.0256 | 7.443 * | 0.0256 | 7.447 | 0.0415 | 29.04 | 1.05 | 28.35 ** | 1.025 | 29.22 | 1.56 | 6.74 | 0.45 | 44.8 | 2.59 |
| 52 | 7.445 * | 0.025 | 7.435 * | 0.0245 | 7.417 | 0.0414 | 29.88 | 2.69 | 28.99 ** | 2.60 | 28.47 | 2.06 | 6.64 | 0.41 | 40.8 ** | 3.49 |
| 67 | 7.44 ** | 0.0184 | 7.435 | 0.0184 | 7.417 | 0.0158 | 30.43 | 1.87 | 29.35 ** | 1.854 | 28.91 | 2.03 | 6.31 | 0.52 | 38.8 * | 1.89 |
| 97 | 7.412 ** | 0.0225 | 7.41 | 0.0225 | 7.394 | 0.0204 | 30.63 | 1.48 | 29.41 ** | 1.456 | 29.25 | 0.93 | 6.212 | 0.69 | 38.0 * | 3.92 |
| [H+] | n | R2 | SEE | p |
|---|---|---|---|---|
| Arterial | ||||
| [H+] temp adj = 0.697 + (0.982 × [H+] meas) | 82 | 1.000 | 0.053 | <0.001 |
| [H+] ion stewart = 6.759 + (0.867 × [H+] meas) | 82 | 0.517 | 3.217 | <0.001 |
| [H+] ion stewart = 6.068 + (0.886 × [H+] temp adj) | 82 | 0.520 | 3.208 | <0.001 |
| Mixed venous | ||||
| [H+] temp adj = 0.522 + (0.987 × [H+] meas) | 68 | 1.000 | 0.048 | <0.001 |
| [H+] stewart = 22.108 + (0.469 × [H+] meas) | 68 | 0.208 | 3.269 | <0.001 |
| [H+] stewart = 21.734 + (0.479 × [H+] temp adj) | 68 | 0.211 | 3.263 | <0.001 |
| TCO2 | ||||
| Arterial | ||||
| TCO2 temp adj = 17.746 + (0.377 × TCO2 meas) | 82 | 0.219 | 1.516 | <0.001 |
| TCO2 stewart = 27.334 + (0.0693 × TCO2 meas) | 82 | 0.00418 | 2.272 | 0.564 |
| TCO2 stewart = 22.390 + (0.242 × TCO2 temp adj) | 82 | 0.0331 | 2.239 | 0.102 |
| Mixed venous | ||||
| TCO2 temp adj = 13.549 + (0.582 × TCO2 meas) | 68 | 0.172 | 3.148 | <0.001 |
| TCO2 = 0.00678 + (0.000782 × TCO2 meas) | 68 | 0.271 | 0.003 | <0.001 |
| TCO2 stewart = 0.0153 + (0.000522 × TCO2 temp adj) | 68 | 0.381 | 0.003 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lindinger, M.I.; Waller, A.P. Physicochemical Analysis of Mixed Venous and Arterial Blood Acid-Base State in Horses at Core Temperature during and after Moderate-Intensity Exercise. Animals 2022, 12, 1875. https://doi.org/10.3390/ani12151875
Lindinger MI, Waller AP. Physicochemical Analysis of Mixed Venous and Arterial Blood Acid-Base State in Horses at Core Temperature during and after Moderate-Intensity Exercise. Animals. 2022; 12(15):1875. https://doi.org/10.3390/ani12151875
Chicago/Turabian StyleLindinger, Michael I., and Amanda P. Waller. 2022. "Physicochemical Analysis of Mixed Venous and Arterial Blood Acid-Base State in Horses at Core Temperature during and after Moderate-Intensity Exercise" Animals 12, no. 15: 1875. https://doi.org/10.3390/ani12151875