
In-Depth Analysis of an Obligate Anaerobe Paraclostridium bifermentans Isolated from Uterus of Bubalus bubalis

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Samples
2.2. Bacterial Isolation and Culture Condition
2.3. Morphological and Biochemical Characterization
2.4. DNA Extraction and Library Preparation
2.5. De Novo Genome Assembly and Annotation
2.6. Identification of Antimicrobial Drug Resistance Genes and Biochemical Pathway
3. Results and Discussion
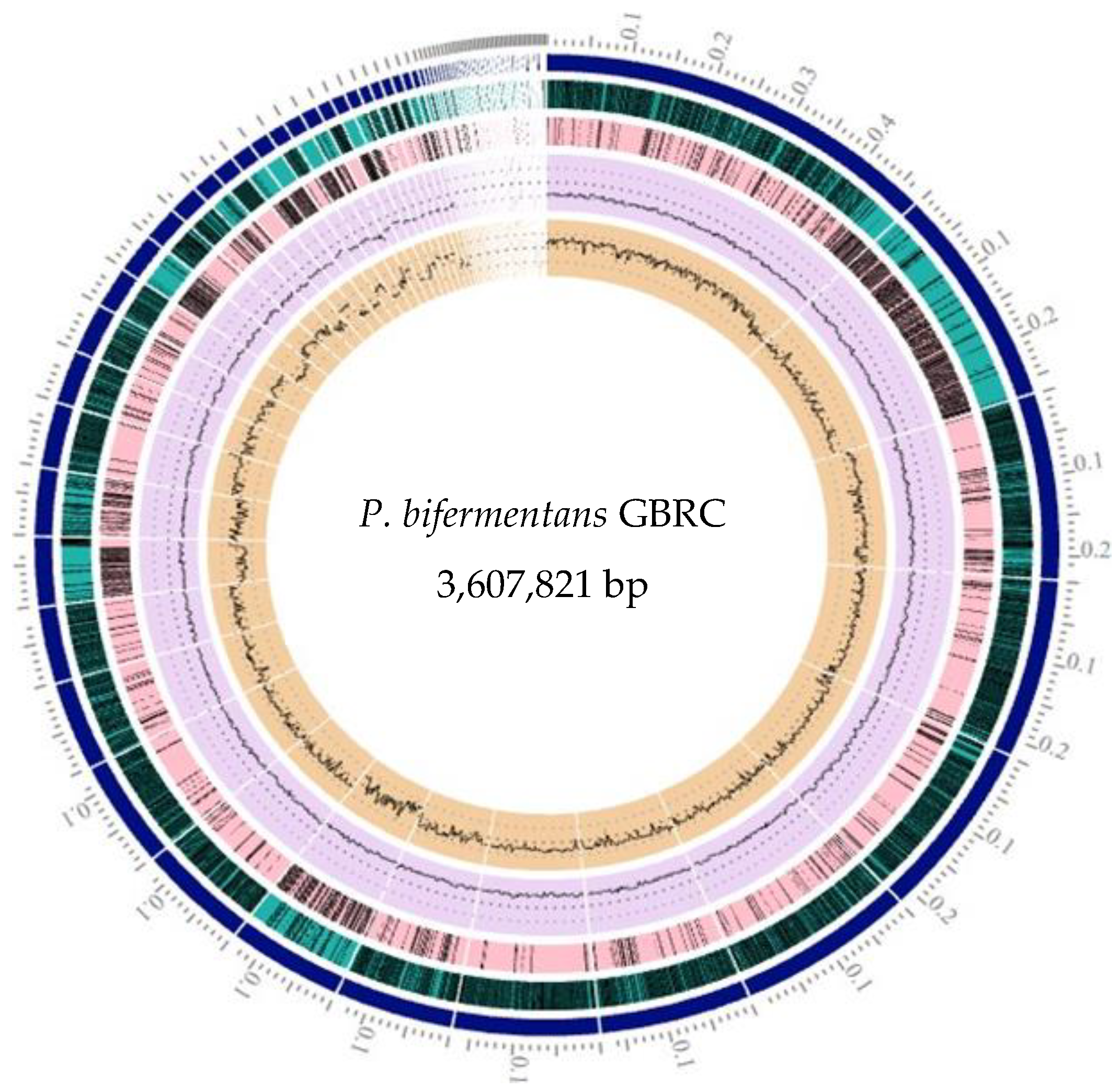
3.1. Morphological Characteristics and Genome Assembly Statistics
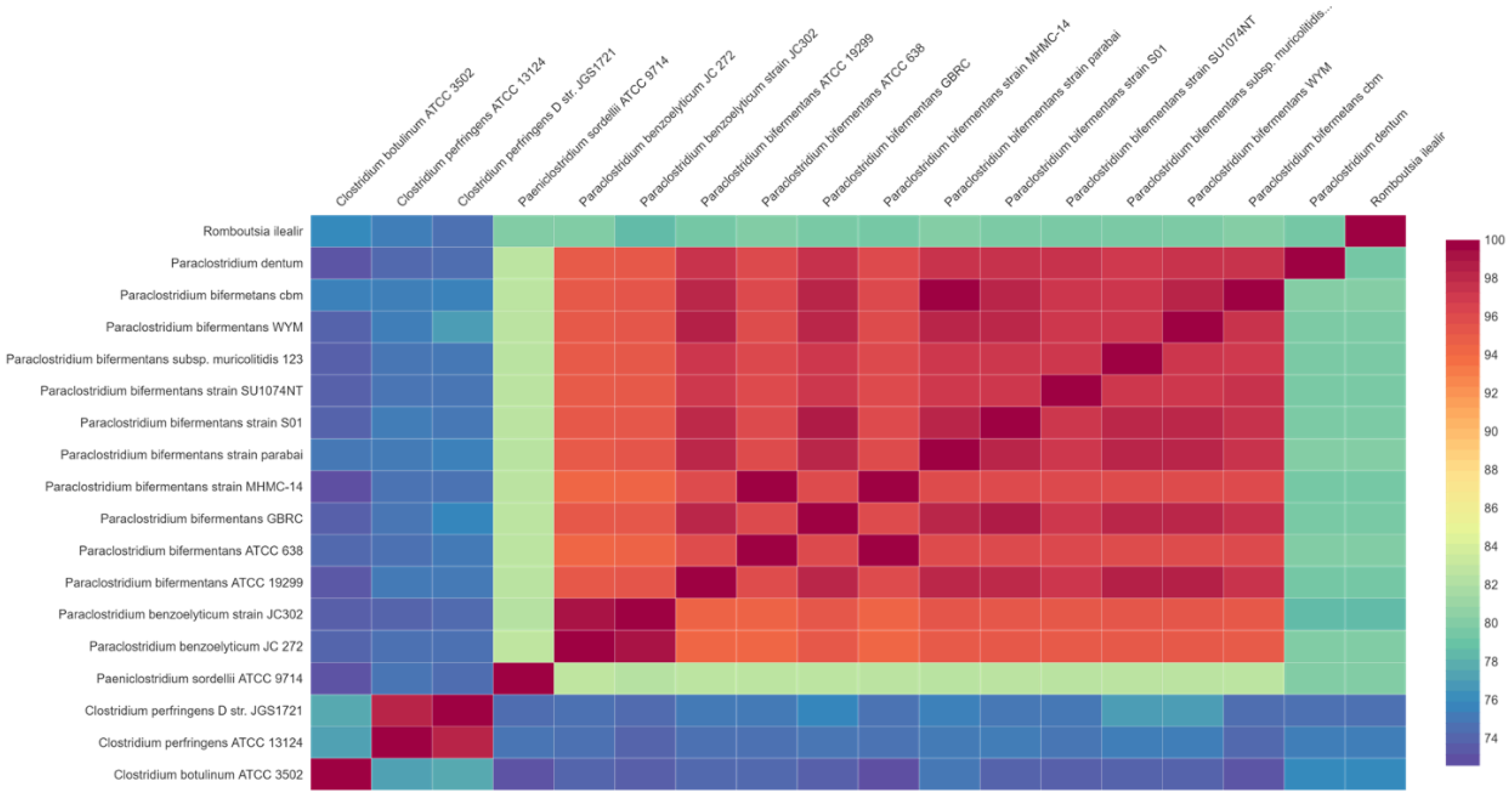
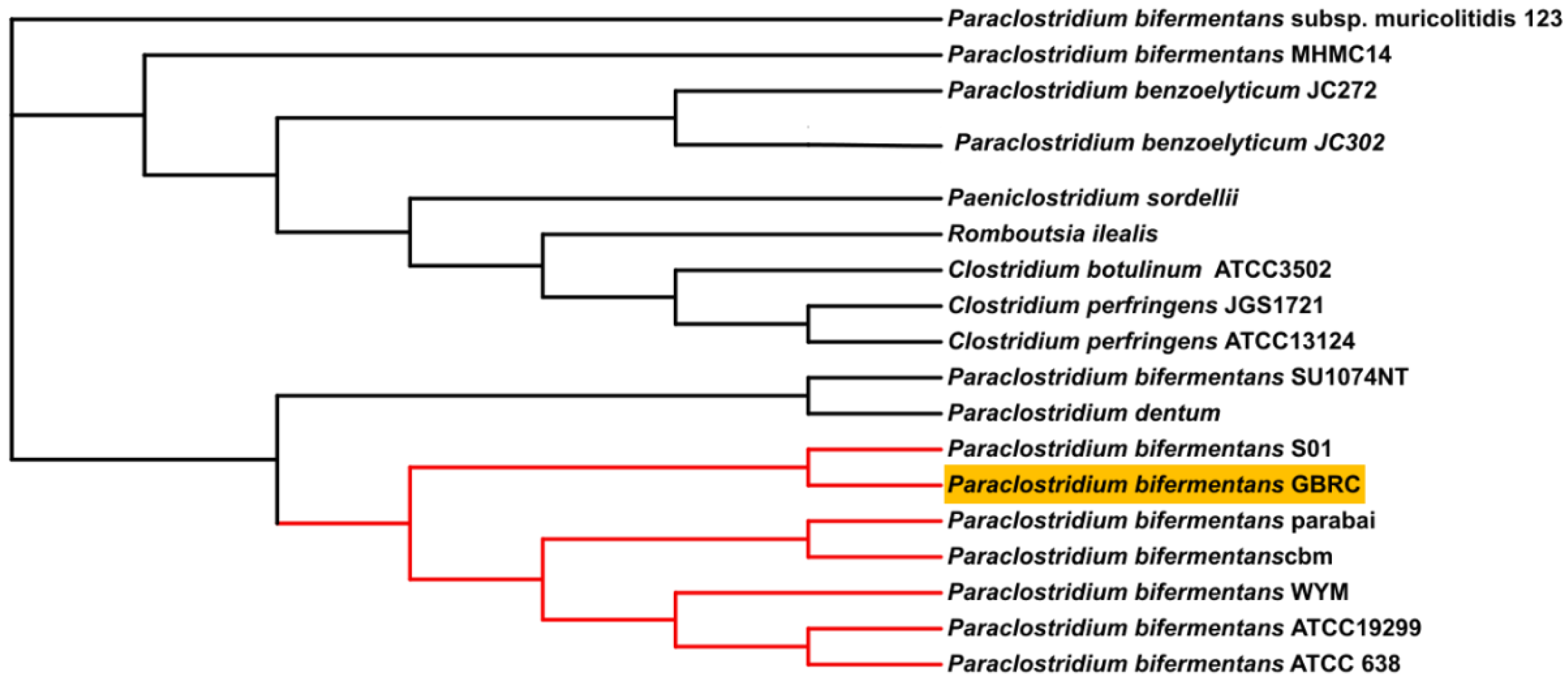
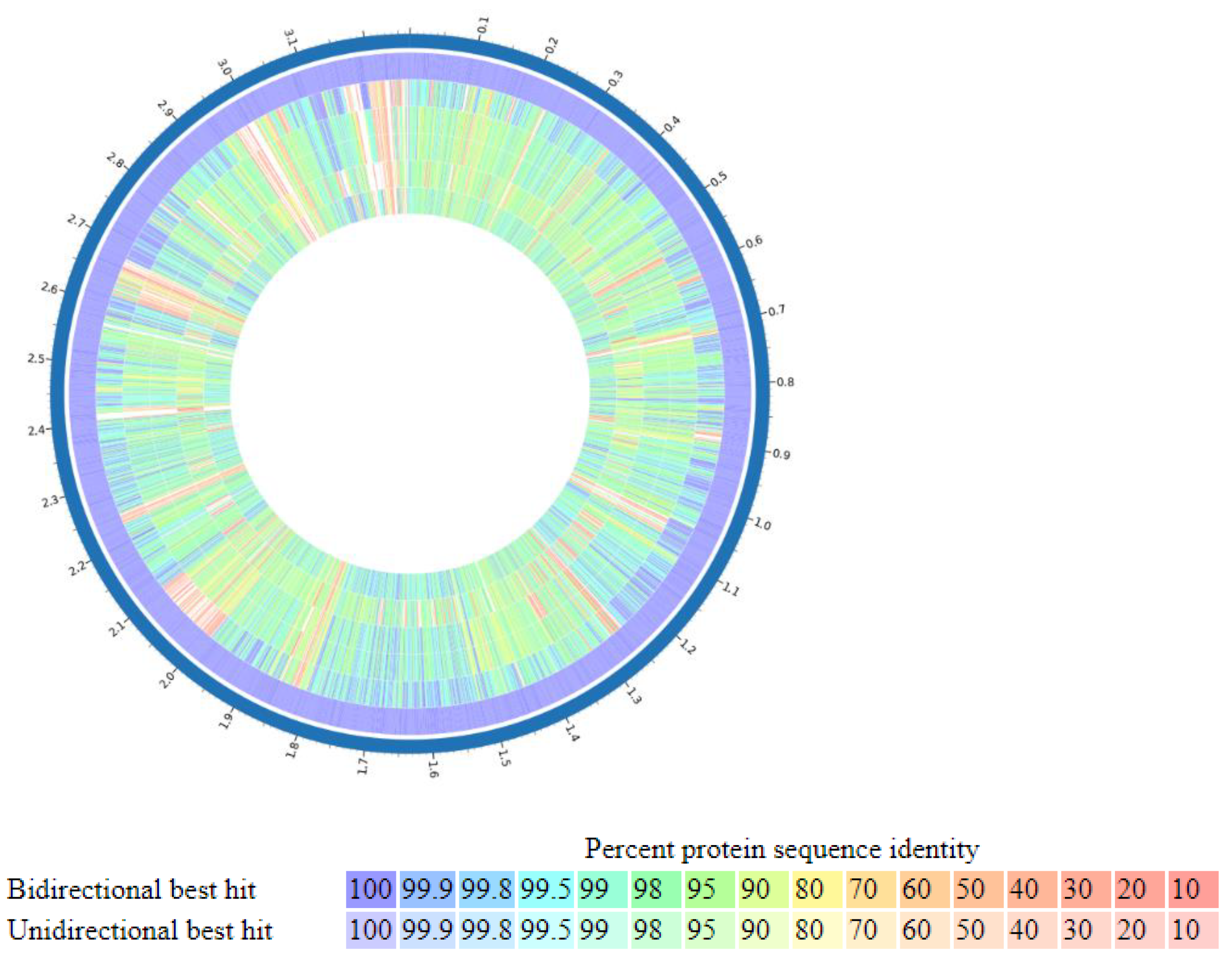
3.2. Comparative Genome Analysis of P. bifermentans GBRC with Closely Related Species
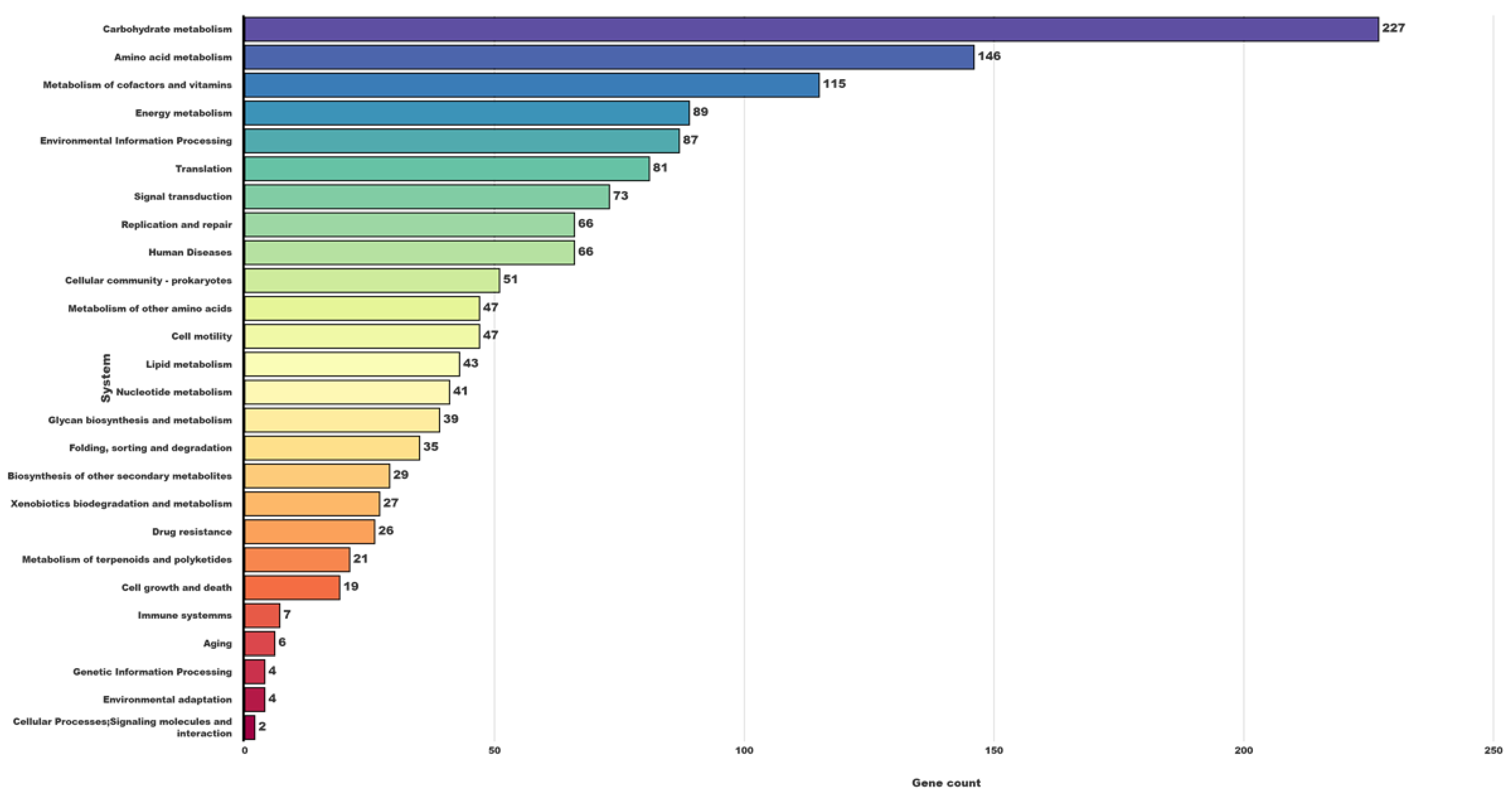
3.3. Gene Predictions and Genome Annotation
3.4. Substrate Utilization and Metabolic Pathway Analysis
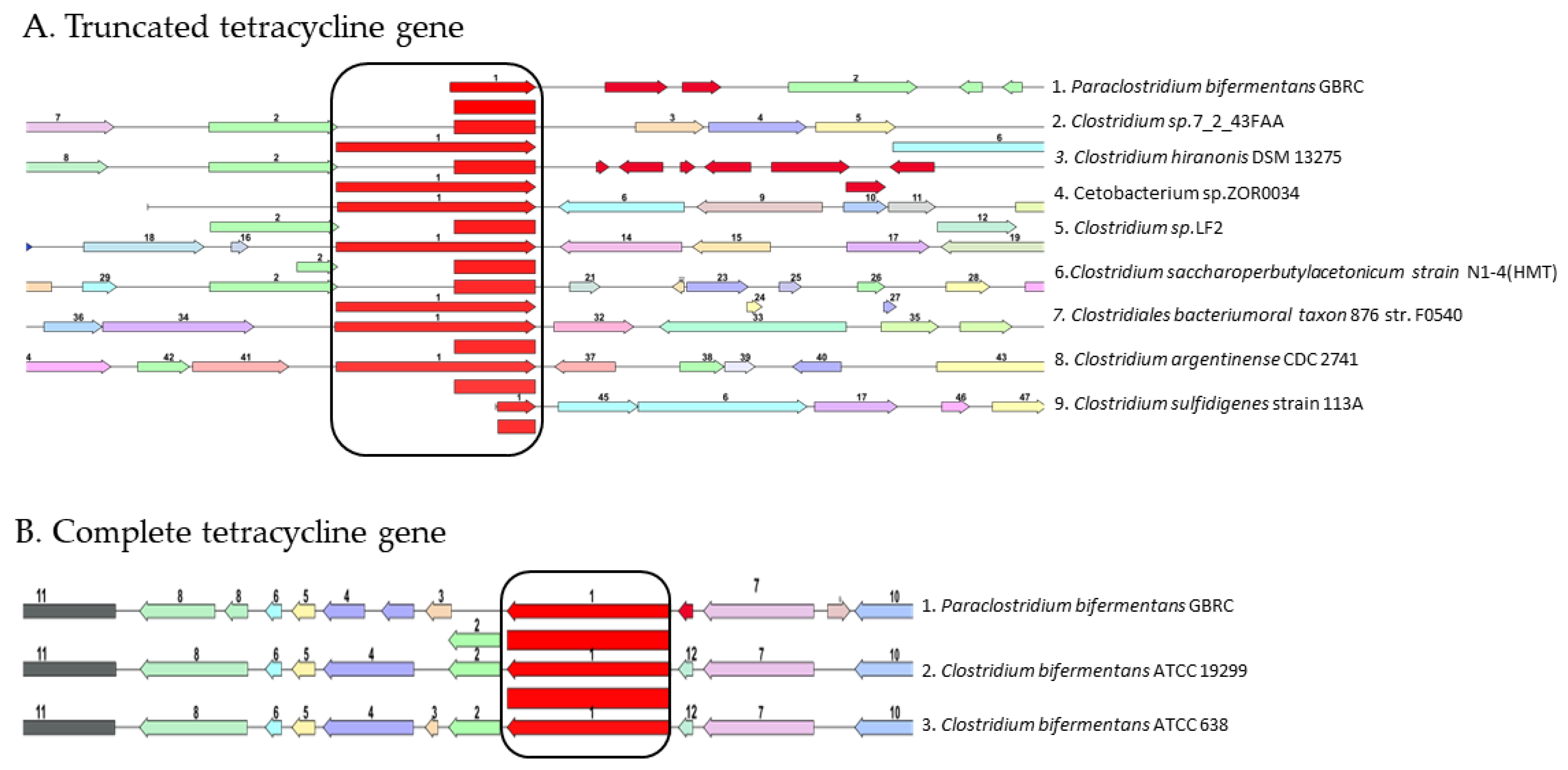
3.5. Antibiotic Sensitivity
4. Conclusions
5. Nucleotide Sequence Accession Numbers
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Philpott, M. The dangers of disease transmission by artificial insemination and embryo transfer. Braz. Vet J. 1993, 149, 339–369. [Google Scholar] [CrossRef]
- Noakes, E.D.; Parkinson, T.J.; England, G.C.W. Post parturient prolapse of the uterus. Arthur’s Vet. Reprod. Obstet. 2001, 8, 333–338. [Google Scholar]
- Sheldon, I.M.; Lewis, G.S.; LeBlanc, S.; Gilbert, O.R. Defining postpartum uterine disease in cattle. Theriogenology 2006, 65, 1516–1530. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, I.M.; Cronin, J.; Goetze, J.; Donofrio, G.; Schuberth, H. Defining postpartum uterine disease and the mechanisms of infection and immunity in the female reproductive tract in Cattle. Biol. Reprod. 2009, 81, 1025–1032. [Google Scholar] [CrossRef]
- Sheldon, I.M.; Owen, S.E. Postpartum uterine infection and endometritis in dairy cattle. In Proceedings of the 33rd Annual Scientific Meeting of the European Embryo Transfer Association (AETE); Anim. Reprod: Bath, UK, 2017; pp. 622–629. [Google Scholar] [CrossRef]
- Azaw, O.I. Uterine infection in buffalo cows; A review. Buffalo Bull 2010, 29, 151–171. Available online: https://ibic.lib.ku.ac.th/e-bulletin/2010-154.pdf (accessed on 3 October 2021).
- Mandhwani, B.A.; Kumar, S.; Shivhare, M.; Aich, R. Insights into bovine endometritis with special reference to phytotherapy. VetWorld 2017, 10, 1529–1532. [Google Scholar] [CrossRef] [Green Version]
- Parmar, K. Endometritis in Bovine: A Review. Agricul. Rev. 2021, 42, 342–347. [Google Scholar] [CrossRef]
- Galvão, K.N.; Bicalho, R.C.; Jeon, S.J. Symposium review: The uterine microbiome associated with the development of uterine disease in dairy cows. J. Dairy Sci. 2019, 102, 11786–11797. [Google Scholar] [CrossRef]
- Hungate, R.E.; Smith, W.; Clarke, R.T. Suitability of butyl rubber stoppers for closing anaerobic roll culture tubes. J. Bacteriol. 1966, 91, 908–909. [Google Scholar] [CrossRef] [Green Version]
- Mah, R.A.; Smith, M.R.; Baresi, L. Studies on an acetate-fermenting strain of Methanosarcina. Appl. Environ. Microbiol. 1978, 35, 1174–1184. [Google Scholar] [CrossRef] [Green Version]
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.A.; Stanley, J.T.; William, S.T. Bergey’s Manual of Determinative Bacteriology. Williams Wilikins Baltim. 1994, 9, 786–788. [Google Scholar]
- Kutsuna, R.; Miyoshi-Akiyama, T.; Mori, K.; Hayashi, M.; Tomida, J.; Morita, Y.; Tanaka, K.; Kawamura, Y. Description of P. bifermentans subsp. muricolitidis subsp. emended description of P. bifermentans and creation of P. bifermentans subsp. Bifermentans subsp. nov. Microbiol. Immunol. 2019, 63, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jyothsna, T.S.; Tushar, L.; Sasikala, C.; Ramana, C.V. Paraclostridium benzoelyticum gen. nov., sp. nov., isolated from marine sediment and reclassification of Clostridium bifermentans as Paraclostridium bifermentans comb. nov. Proposal of a new genus Paeniclostridium gen. nov. to accommodate Clostridium sordellii and Clostridium ghonii. Int. J. Syst. Evol. Microbiol. 2016, 66, 1268–1274. [Google Scholar] [CrossRef]
- Wilson, K. Preparation of genomic DNA from bacteria. Curr. Protoc. Mol. Biol. 2001, 56, 2–4. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. A Quality Control Tool for High throughput Sequence Data. FastQC. 2014. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 6 October 2021).
- Simpson, J.T.; Wong, K.; Jackman, S.D.; Schein, J.E.; Jones, S.J.; Birol, I. ABySS: A parallel assembler for short read sequence data. Genome Res. 2009, 19, 1117–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zerbino, D.R. Using the Velvet de novo assembler for short-read sequencing technologies. Curr. Protoc. Bioinform. 2010, 11, Unit 11.5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, R.; Liu, B.; Xie, Y.; Li, Z.; Huang, W.; Yuan, J.; He, G.; Chen, Y.; Pan, Q.; Liu, Y.; et al. SOAPdenovo2: An empirically improved memory-efficient short-read de novo assembler. GigaScience 2012, 1, 18. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comp. Biol. 2012, 19, 455–2477. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Zhao, L.; Gao, J.; Fei, Z. I Assembler: A package for de novo assembly of Roche-454/Sanger transcriptome sequences. BMC Bioinform. 2011, 12, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Hyatt, D.; Chen, G.L.; LoCascio, P.F.; Miriam, L.; Frank, W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and ranslation initiation site identification. BMC Bio. 2010, 11, 119. [Google Scholar] [CrossRef] [Green Version]
- Kelley, D.R.; Liu, B.; Delcher, A.L.; Pop, M.; Salzberg, S.L. Gene prediction with Glimmer for metagenomics sequences augmented by classification and clustering. Nucleic Acids Res. 2012, 40, e9. [Google Scholar] [CrossRef] [PubMed]
- Besemer, J.; Lomsadze, A.; Borodovsky, M. GeneMarkS: A self-training method for prediction of gene starts in microbial genomes. Implications for finding sequence motifs in regulatory regions. Nucleic Acids Res. 2001, 29, 2607–2618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noguchi, H.; Taniguchi, T.; Itoh, T. MetaGeneAnnotator: Detecting species-specific patterns of ribosomal binding site for precise gene prediction in anonymous prokaryotic and phage genomes. DNA Res. Int. J. Rap. Pub. Repo Genes Genomes 2008, 15, 387–396. [Google Scholar] [CrossRef] [Green Version]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the rapid annotation of microbial genomes using subsystems technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef] [PubMed]
- Tanizawa, Y.; Fujisawa, T.; Nakamura, Y. DFAST: A flexible prokaryotic genome annotation pipeline for faster genome publication. Bioinformatics 2018, 34, 1037–1039. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef]
- Rodriguez-R, L.M.; Konstantinidis, K.T. The Enveomics Collection: A Toolbox for Specialized Analyses of Microbial Genomes and Metagenomes (No. e1900v1). PeerJ Prepr. 2016, 4, e1900v1. [Google Scholar]
- Letunic, I.; Bork, P. Nucleic Acids Res Interactive Tree Of Life v2: Online annotation and display of phylogenetic trees made easy. Nucleic Acids Res. 2011, 39, W475–W478. [Google Scholar] [CrossRef]
- Knudsen, S. Promoter2.0: For the recognition of PolII promoter sequences. Bioinformatics 1999, 15, 356–361. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bray, J.E.; Maiden, M. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Fujikawa, T.; Sawada, H. Genome analysis of the kiwifruit canker pathogen Pseudomonas syringae pv actinidiae biovar 5. Sci. Rep. 2016, 6, 21399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef] [PubMed]
- Simonsen, M.; Mailund, T.; Pedersen, C.N. Rapid neighbour-joining. In International Workshop on Algorithms in Bioinformatics; Springer: Berlin/Heidelberg, Germany, 2008; pp. 113–122. [Google Scholar]
- Minder, A.C.; Fischer, H.M.; Hennecke, H.; Narberhaus, F. Role of HrcA and CIRCE in the heat shock regulatory network of Bradyrhizobium japonicum. J. Bacteriol. 2000, 182, 14–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating viruses and cellular organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef]
- Martens, E.C.; Neumann, M.; Desai, M.S. Interactions of commensal and pathogenic microorganisms with the intestinal mucosal barrier. Nat. Rev. Microbiol. 2018, 16, 457–470. [Google Scholar] [CrossRef]
- O’Toole, G.A.; Kolter, R. Flagellar and twitching motility are necessary for Pseudomonas aeruginosa biofilm development. Mol. Microbiol. 1998, 30, 295–304. [Google Scholar] [CrossRef]
- Pratt, L.A.; Kolter, R. Genetic analysis of Escherichia coli biofilm formation: Roles of flagella, motility, chemotaxis and type I. pili. Mol. Microbiol. 1998, 30, 285–293. [Google Scholar] [CrossRef]
- Prigent-Combaret, C.; Vidal, O.; Dorel, C.; Lejeune, P. Abiotic surface sensing and biofilm-dependent regulation of gene expression in Escherichia coli. J. Bacteriol. 1999, 181, 5993–6002. [Google Scholar] [CrossRef] [Green Version]
- Watnick, P.I.; Kolter, R. Steps in the development of a Vibrio cholerae El Tor biofilm. Mol. Microbiol. 1999, 34, 586–595. [Google Scholar] [CrossRef] [Green Version]
- Haiko, J.; Westerlund-Wikström, B. The role of the bacterial flagellum in adhesion and virulence. Biology 2013, 2, 1242–1267. [Google Scholar] [CrossRef] [Green Version]
- Raheel, I.A.E.R.; Hassan, W.H.; Salem, S.S.R.; Salam, H.S.H. Biofilm forming potentiality of Escherichia coli isolated from bovine endometritis and their antibiotic resistance profiles. J. Adv. Vet. Anim. Res. 2020, 7, 442. [Google Scholar] [CrossRef] [PubMed]
- Gurunathan, S.; Choi, Y.J.; Kim, J.H. Antibacterial efficacy of silver nanoparticles on endometritis caused by Prevotella melaninogenica and Arcanobacterum pyogenes in dairy cattle. Int. J. Mol. Sci. 2018, 19, 1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braoudaki, M.; Hilton, A.C. Mechanisms of resistance in Salmonella enterica adapted to erythromycin, benzalkonium chloride and triclosan. Int. J. Antimicrob. Agents 2005, 25, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Ravanel, N.; Gestin, B.; Maurin, M. In vitro selection of fluoroquinolone resistance in Brucella melitensis. Int. J. Antimicrob. Agents 2009, 34, 76–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, M.B.; Martínez, J.L. Overexpression of the efflux pumps SmeVWX and SmeDEF is a major cause of resistance to co-trimoxazole in Stenotrophomonas maltophilia. Antimicrob. Agents Chemother. 2018, 62, e00301-18. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, M.; Hirt, H.; Zurek, L. Horizontal transfer of the tetracycline resistance gene tetM mediated by pCF10 among Enterococcus faecalis in the house fly (Musca domestica L.) alimentary canal. Microbiol. Ecol. 2009, 58, 509–518. [Google Scholar] [CrossRef]
- Von Wintersdorff, C.J.; Penders, J.; Van Niekerk, J.M.; Mills, N.D.; Majumder, S.; Van Alphen, L.B.; Wolffs, P.F. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front. Microbiol 2016, 7, 173. [Google Scholar] [CrossRef] [Green Version]
- Farca, A.M.; Nebbia, P.; Robino, P.; Re, G. Effects of the combination antibiotic—edta–tris in the treatment of chronic bovine endometritis caused by antimicrobial-resistant bacteria. Pharm. Res. 1997, 36, 3539. [Google Scholar] [CrossRef]
- Bicalho, M.L.; Machado, V.S.; Oikonomou, G.; Gilbert, R.O.; Bicalho, R.C. Association between virulence factors of Escherichia coli, Fusobacterium necrophorum, and Arcanobacterium pyogenes and uterine diseases of dairy cows. Vet. Microbiol. 2012, 157, 125–131. [Google Scholar] [CrossRef]
- Contreras, E.; Masuyer, G.; Qureshi, N.; Chawla, S.; Dhillon, H.S.; Lee, H.L.; Chen, J.; Stenmark, P.; Gill, S.S. A neurotoxin that specifically targets Anopheles mosquitoes. Nat. Commun. 2019, 10, 2869. [Google Scholar] [CrossRef]
- Mackeen, A.D.; Packard, R.E.; Ota, E.; Speer, L. Antibiotic regimens for postpartum endometritis. Cochrane Database Syst. Rev. 2015. [Google Scholar] [CrossRef]
- LeBlanc, S.J. Postpartum uterine disease and dairy herd reproductive performance: A review. Vet. J. 2008, 176, 102–114. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assembler Platform | Total Number of Scaffolds | N50 Size (bp) | Longest Contig’s Length |
|---|---|---|---|
| ABySS | 73,275 | 240 | 1237 |
| Velvet | 4451 | 502 | 3135 |
| SOAPdenovo2 | 3968 | 664 | 4829 |
| SPAdes | 178 | 165,665 | 517,124 |
| Newbler 2.7 | 78 | 177,236 | 477,527 |
| Feature | Paraclostridium bifermentans GBRC |
|---|---|
| Domain | Bacteria |
| Taxonomy | Firmicutes; Clostridia; Eubacteriales; Peptostreptococcaceae; Paraclostridium; |
| Paraclostridium bifermentans | |
| Genome size | 3,607,821 bp |
| N50 size(bp) | 177,236 bp |
| Total Num. of contigs | 78 |
| G + C content | 28.10% |
| Contamination | 0% |
| Completeness | 0.99 |
| Genes | 3511 |
| CDSs (Total) | 3441 |
| Genes (Coding) | 3.395 |
| Genes(RNA) | 70 |
| rRNAs | 3, 3, 5 (5 S, 16 S, 23 S) |
| Complete rRNAs | 3 (5 S) |
| Partial rRNAs | 3, 5 (16 S, 23 S) |
| tRNAs | 55 |
| ncRNAs | 4 |
| Virulence Class | Virulence Factors | Related Genes | Predicated Gene ID |
|---|---|---|---|
| Adherence | Fibronectin-binding protein | fbpA/fbp68 | gene_12_113 |
| GroEL | groEL | gene_11_64 | |
| Flagella (Pseudomonas) | flip | gene_3_40 | |
| LPS O-antigen (P. aeruginosa) | gene_1_129, gene_1_130 | ||
| Listeria adhesion protein | Lap | gene_7_23 | |
| Regulation | CheA/CheY (Listeria) | cheY | gene_3_167 |
| LisR/LisK (Listeria) | lisR | gene_19_8, gene_8_135 | |
| Sigma A (Mycobacterium) | sigA/rpoV | gene_9_93 | |
| Toxin | Alpha-toxin | Plc | gene_18_29 |
| Hemolysin | gene_11_116 | ||
| gene_8_36 | |||
| Kappa-toxin (collagenase) | colA | gene_11_57 | |
| Perfrigolysin O (theta-toxin/PFO) /botulinolysin | pfoA | gene_6_144 | |
| Cytolysin | cylR2 | gene_10_57 | |
| Antiphagocytosis | Capsular polysaccharide | rmlB | gene_12_15 |
| Capsular polysaccharide | wcaJ | gene_8_159 | |
| Capsule | gene_3_150 | ||
| Cell surface Components | Trehalose-recycling ABC transporter | sugC | gene_13_45 |
| Immune evasion | Capsule | cps4I | gene_26_21 |
| gene_8_154 | |||
| LPS | acpXL | gene_26_5 | |
| Polysaccharide capsule | gene_8_145 | ||
| gene_8_146 | |||
| gene_8_155 | |||
| gene_8_161 | |||
| Polysaccharide capsule | epsE | gene_12_3 | |
| manA | gene_12_22 | ||
| Iron uptake | Heme biosynthesis | hemB | gene_25_23 |
| Periplasmic binding protein | vctC | gene_13_103 | |
| Nutritional virulence | Pyrimidine biosynthesis | gene_11_52 | |
| Secretoin system | T6SS-II | gene_29_17 | |
| Serum resistance and immune evastion | Capsule | gene_12_130 | |
| LPS | wbtE | gene_8_165 | |
| LPS | wbtF | gene_8_166 | |
| LPS | wbtI | gene_3_25 |
| Characteristic | 1 | 2 | 3 | 4 |
|---|---|---|---|---|
| Gram-stain | Gram-Positive | Gram-Positive | Gram-Positive | Gram-Positive |
| Shape | Rod Shaped | Rod Shaped | Rod Shaped | Rod Shaped |
| Sporulation | Central to Terminal | Terminal | Terminal to Sub terminal | Terminal |
| Motility | Motile | Motile | Motile | Motile |
| Obligate Anaerobes | Obligate Anaerobe | Obligate Anaerobe | Obligate Anaerobe | Obligate Anaerobe |
| Catalase | − | − | − | − |
| Oxidase | − | − | − | − |
| Indole | + | − | + | + |
| H2S production | +/− | + | − | + |
| Starch hydrolysis | − | + | − | + |
| Gelatin hydrolysis | + | + | + | + |
| Nitrate reduction | − | − | − | − |
| Lysine Decarboxylase | + | + | + | − |
| Casein | + | + | + | + |
| Classification | Substrate | Predicted KEGG Pathway ID | |
|---|---|---|---|
| System | Subsystem | ||
| Carbohydrates | Ketose | d-fructose | map00051, map00052, & map00500 |
| Monosaccharide | α–d-Glucose | map00010, map00052, & map00520 | |
| d-Mannose | map00051 | ||
| α–d-Glucose 6 phosphate | map00010, map00030, & map00052 | ||
| Galactose | map00052 | ||
| Hexosamine | N-Acetyl-d-Galactosamine | map00052 | |
| Sugar Alcohol | Sorbitol | map00051 | |
| Nucleoside | Inosine | map00230 | |
| Amino acids | l-Asparagine | map00250, & map00460 | |
| l-Glutamine | map00250 | ||
| l-Threonine | map00260 | ||
| l-Methionine | map00270 | ||
| l-Serine | map00260, & map00270 | ||
| l-Proline | map00330, & map00470 | ||
| l-Phenylalanine | map00360 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gohil, P.; Patel, K.; Patel, S.; Pandit, R.; Suthar, V.; Duggirala, S.; Joshi, M.; Patil, D.; Joshi, C. In-Depth Analysis of an Obligate Anaerobe Paraclostridium bifermentans Isolated from Uterus of Bubalus bubalis. Animals 2022, 12, 1765. https://doi.org/10.3390/ani12141765
Gohil P, Patel K, Patel S, Pandit R, Suthar V, Duggirala S, Joshi M, Patil D, Joshi C. In-Depth Analysis of an Obligate Anaerobe Paraclostridium bifermentans Isolated from Uterus of Bubalus bubalis. Animals. 2022; 12(14):1765. https://doi.org/10.3390/ani12141765
Chicago/Turabian StyleGohil, Purva, Kajal Patel, Srushti Patel, Ramesh Pandit, Vishal Suthar, Srinivas Duggirala, Madhvi Joshi, Deepak Patil, and Chaitanya Joshi. 2022. "In-Depth Analysis of an Obligate Anaerobe Paraclostridium bifermentans Isolated from Uterus of Bubalus bubalis" Animals 12, no. 14: 1765. https://doi.org/10.3390/ani12141765