
Dietary Inclusion of Dried Chicory Root Affects Cecal Mucosa Proteome of Nursery Pigs

 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Study Design and Sample Collection
2.2. Two-Dimensional Electrophoresis (2-DE)
2.3. Image Acquisition and Data Analysis
2.4. Mass Spectrometry and Bioinformatic Data Analysis
2.5. Statistical Analysis
3. Results
3.1. Production Traits
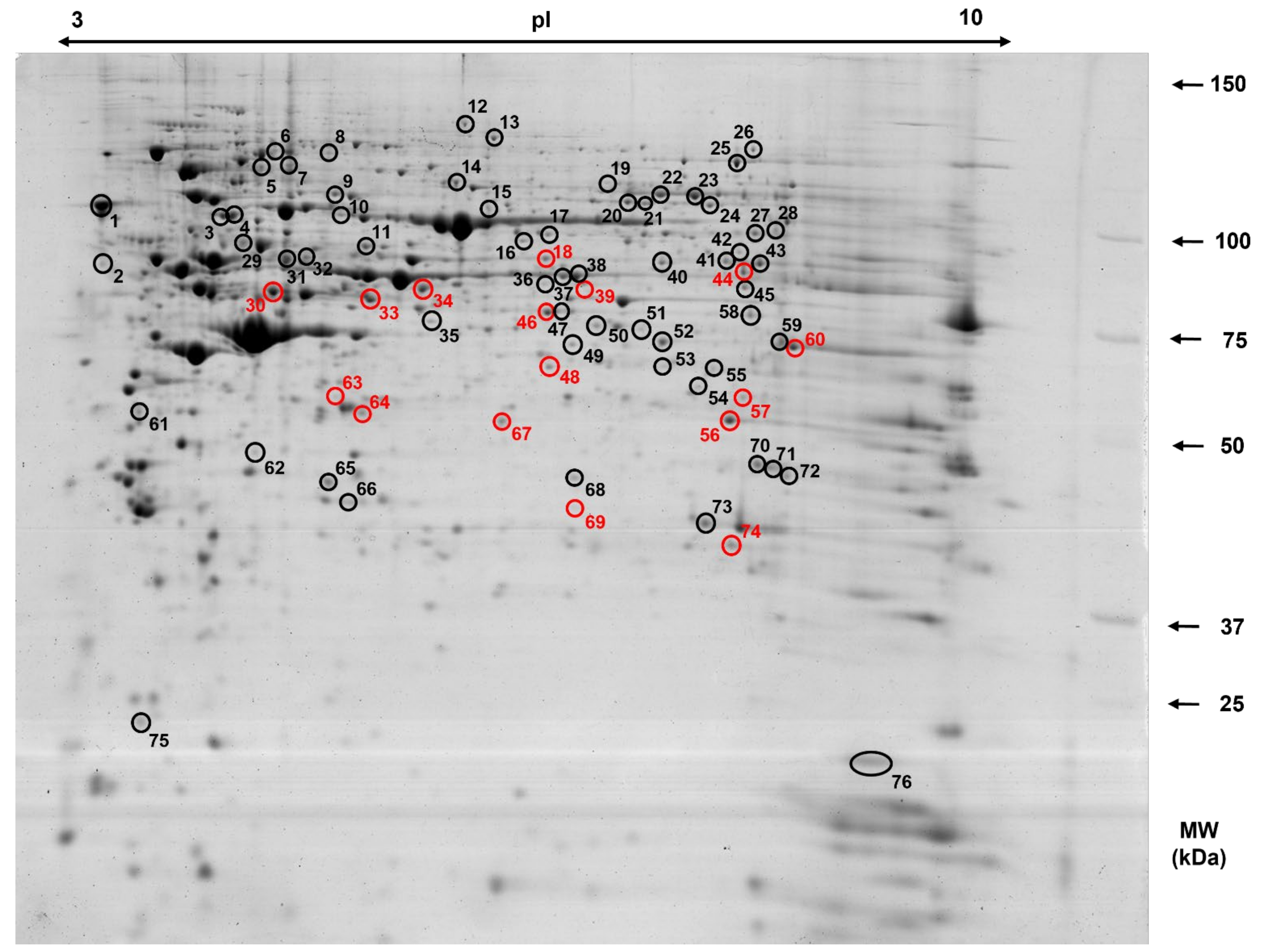
3.2. Two-Dimensional (2-D) Gel-Based Cecal Mucosa Protein Profile of 50-Day-Old Piglets
3.3. Differentially Expressed Proteins in the Cecal Mucosa in Response to CR Diet
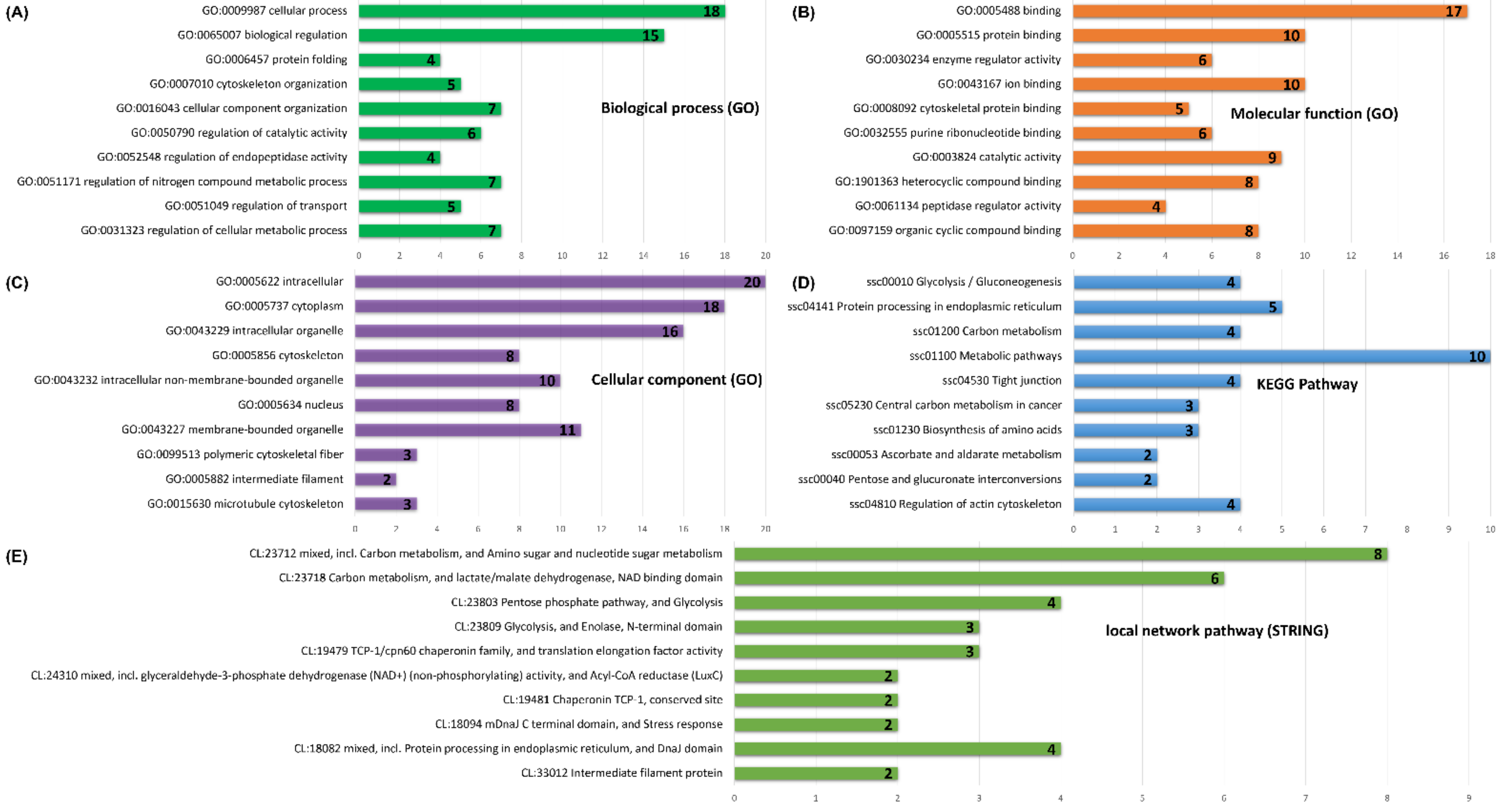
3.4. Functional Annotations and Pathway Enrichment Analysis of Porcine Cecal Mucosa Proteins
3.5. STRING Analysis of Protein Networks
4. Discussion
4.1. 2-D Gel-Based Cecal Mucosa Protein Profile of 50-Day-Old Piglets
4.2. Differentially Expressed Proteins in the Cecal Mucosa in Response to CR Diet
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saffrey, M.J. Aging of the mammalian gastrointestinal tract: A complex organ system. Age 2014, 36, 9603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, P.; Liu, Y.; Le, B.; Qin, B.; Liu, M.; Zhao, Y.; Guo, X.; Cao, G.; Liu, J.; Li, B.; et al. A comparison of dynamic distributions of intestinal microbiota between Large White and Chinese Shanxi Black pigs. Arch. Microbiol. 2019, 201, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Wang, Y.; Liu, S.; Huang, J.; Zhai, Z.; He, C.; Ding, J.; Wang, J.; Wang, H.; Fan, W.; et al. The dynamic distribution of porcine microbiota across different ages and gastrointestinal tract segments. PLoS ONE 2015, 10, e0117441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celi, P.; Verlhac, V.; Pérez, C.E.; Schmeisser, J.; Kluenter, A.M. Biomarkers of gastrointestinal functionality in animal nutrition and health. Anim. Feed Sci. Technol. 2019, 250, 9–31. [Google Scholar] [CrossRef]
- Nwafor, I.C.; Shale, K.; Achilonu, M.C. Chemical composition and nutritive benefits of chicory (Cichorium intybus) as an ideal complementary and/or alternative livestock feed supplement. Sci. World J. 2017, 2017, 7343928. [Google Scholar] [CrossRef] [Green Version]
- Mensink, M.A.; Frijlink, H.W.; van der Voort, M.; Hinrichs, W.L. Inulin, a flexible oligosaccharide I: Review of its physicochemical characteristics. Carbohydr. Polym. 2015, 130, 405–419. [Google Scholar] [CrossRef] [Green Version]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Lepczyński, A.; Herosimczyk, A.; Ożgo, M.; Barszcz, M.; Taciak, M.; Skomiał, J. Modification of ileal proteome in growing pigs by dietary supplementation with inulin or dried chicory root. J. Anim. Feed Sci. 2019, 28, 177–186. [Google Scholar] [CrossRef]
- Lepczyński, A.; Herosimczyk, A.; Ożgo, M.; Marynowska, M.; Pawlikowska, M.; Barszcz, M.; Taciak, M.; Skomiał, J. Dietary chicory root and chicory inulin trigger changes in energetic metabolism, stress prevention and cytoskeletal proteins in the liver of growing pigs—A proteomic study. J. Anim. Physiol. Anim. Nutr. 2017, 101, e225–e236. [Google Scholar] [CrossRef]
- Lepczynski, A.; Herosimczyk, A.; Ozgo, M.; Skomial, J.; Taciak, M.; Barszcz, M.; Berezecka, N. Dietary supplementation with dried chicory root triggers changes in the blood serum proteins engaged in the clotting process and the innate immune response in growing pigs. J. Physiol. Pharmacol. 2015, 66, 47–55. [Google Scholar]
- Yasuda, K.; Maiorano, R.; Welch, R.M.; Miller, D.D.; Lei, X.G. Cecum is the major degradation site of ingested inulin in young pigs. J. Nutr. 2007, 137, 2399–2404. [Google Scholar] [CrossRef] [Green Version]
- Patterson, J.K.; Yasuda, K.; Welch, R.M.; Miller, D.D.; Lei, X.G. Supplemental dietary inulin of variable chain lengths alters intestinal bacterial populations in young pigs. J. Nutr. 2010, 140, 2158–2161. [Google Scholar] [CrossRef]
- Barszcz, M.; Taciak, M.; Skomiał, J. Influence of different inclusion levels and chain length of inulin on microbial ecology and the state of mucosal protective barrier in the large intestine of young pigs. Anim. Prod. Sci. 2018, 58, 1109–1118. [Google Scholar] [CrossRef]
- Paßlack, N.; Al-samman, M.; Vahjen, W.; Männer, K.; Zentek, J. Chain length of inulin affects its degradation and the microbiota in the gastrointestinal tract of weaned piglets after a short-term dietary application. Livest. Sci. 2012, 149, 128–136. [Google Scholar] [CrossRef]
- Loh, G.; Eberhard, M.; Brunner, R.M.; Hennig, U.; Kuhla, S.; Kleessen, B.; Metges, C.C. Inulin alters the intestinal microbiota and short-chain fatty acid concentrations in growing pigs regardless of their basal diet. J. Nutr. 2006, 136, 1198–1202. [Google Scholar] [CrossRef]
- Pouille, C.L.; Ouaza, S.; Roels, E.; Behra, J.; Tourret, M.; Molinié, R.; Fontaine, J.-X.; Mathiron, D.; Gagneul, D.; Taminiau, B.; et al. Chicory: Understanding the effects and effectors of this functional food. Nutrients 2022, 14, 957. [Google Scholar] [CrossRef]
- Lepczyński, A.; Herosimczyk, A.; Barszcz, M.; Ożgo, M.; Michałek, K.; Grabowska, M.; Tuśnio, A.; Szczerbińska, D.; Skomiał, J. Diet supplemented either with dried chicory root or chicory inulin significantly influence kidney and liver mineral content and antioxidative capacity in growing pigs. Animal 2021, 15, 100129. [Google Scholar] [CrossRef]
- Fouhse, J.M.; Zijlstra, R.T.; Willing, B.P. The role of gut microbiota in the health and disease of pigs. Anim. Front. 2016, 6, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Whitley, C.S.; Haribabu, B.; Jala, V.R. Regulation of intestinal barrier function by microbial metabolites. Cell Mol. Gastroenterol. Hepatol. 2021, 11, 1463–1482. [Google Scholar] [CrossRef]
- Herosimczyk, A.; Lepczyński, A.; Ożgo, M.; Tuśnio, A.; Taciak, M.; Barszcz, M. Effect of dietary inclusion of 1% or 3% of native chicory inulin on the large intestinal mucosa proteome of growing pigs. Animal 2020, 14, 1647–1658. [Google Scholar] [CrossRef]
- Barmpatsalou, V.; Dubbelboer, I.R.; Rodler, A.; Jacobson, M.; Karlsson, E.; Pedersen, B.L.; Bergström, C.A.S. Physiological properties, composition and structural profiling of porcine gastrointestinal mucus. Eur. J. Pharm. Biopharm. 2021, 169, 156–167. [Google Scholar] [CrossRef] [PubMed]
- AOAC International. Official Methods of Analysis of AOAC International, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2011. [Google Scholar]
- Pink, M.; Verma, N.; Rettenmeier, A.W.; Schmitz-Spanke, S. CBB staining protocol with higher sensitivity and mass spectrometric compatibility. Electrophoresis 2010, 31, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Ozgo, M.; Lepczynski, A.; Robak, P.; Herosimczyk, A.; Marynowska, M. The current proteomic landscape of the porcine liver. J. Physiol. Pharmacol. 2019, 70, 369–387. [Google Scholar]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tröscher-Mußotter, J.; Tilocca, B.; Stefanski, V.; Seifert, J. Analysis of the bacterial and host proteins along and across the porcine gastrointestinal tract. Proteomes 2019, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Kiseleva, O.; Ponomarenko, E.; Poverennaya, E. Empowering shotgun mass spectrometry with 2DE: A HepG2 study. Int. J. Mol. Sci. 2020, 21, 3813. [Google Scholar] [CrossRef]
- Suzuki, T. Regulation of the intestinal barrier by nutrients: The role of tight junctions. Anim. Sci. J. 2020, 91, e13357. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Yu, M. Role of goblet cells in intestinal barrier and mucosal immunity. J. Inflamm. Res. 2021, 14, 3171–3183. [Google Scholar] [CrossRef]
- Johansson, M.E.; Thomsson, K.A.; Hansson, G.C. Proteomic analyses of the two mucus layers of the colon barrier reveal that their main component, the Muc2 mucin, is strongly bound to the Fcgbp protein. J. Proteome Res. 2009, 8, 3549–3557. [Google Scholar] [CrossRef]
- Wang, H.; Xu, R.; Zhang, H.; Su, Y.; Zhu, W. Swine gut microbiota and its interaction with host nutrient metabolism. Anim. Nutr. 2020, 6, 410–420. [Google Scholar] [CrossRef]
- Van der Wielen, N.; Moughan, P.J.; Mensink, M. Amino acid absorption in the large intestine of humans and porcine models. J. Nutr. 2017, 147, 1493–1498. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, L.S.; Fanning, A.S. Regulation of epithelial permeability by the actin cytoskeleton. Cytoskeleton 2011, 68, 653–660. [Google Scholar] [CrossRef] [Green Version]
- Diao, H.; Jiao, A.R.; Yu, B.; Mao, X.B.; Chen, D.W. Gastric infusion of short-chain fatty acids can improve intestinal barrier function in weaned piglets. Genes Nutr. 2019, 14, 4. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.; He, T.; Becker, S.; Zhang, G.; Li, D.; Ma, X. Butyrate: A double-edged sword for health? Adv. Nutr. 2018, 9, 21–29. [Google Scholar] [CrossRef] [Green Version]
- Darcy-Vrillon, B.; Cherbuy, C.; Morel, M.T.; Durand, M.; Duée, P.H. Short chain fatty acid and glucose metabolism in isolated pig colonocytes: Modulation by NH4+. Mol. Cell. Biochem. 1996, 156, 145–151. [Google Scholar] [CrossRef]
- Liu, H.; Ivarsson, E.; Dicksved, J.; Lundh, T.; Lindberg, J.E. Inclusion of chicory (Cichorium intybus L.) in pigs' diets affects the intestinal microenvironment and the gut microbiota. Appl. Environ. Microbiol. 2012, 78, 4102–4109. [Google Scholar] [CrossRef] [Green Version]
- Polari, L.; Alam, C.M.; Nyström, J.H.; Heikkilä, T.; Tayyab, M.; Baghestani, S.; Toivola, D.M. Keratin intermediate filaments in the colon: Guardians of epithelial homeostasis. Int. J. Biochem. Cell Biol. 2020, 129, 105878. [Google Scholar] [CrossRef]
- Zhou, Q.; Cadrin, M.; Herrmann, H.; Chen, C.H.; Chalkley, R.J.; Burlingame, A.L.; Omary, M.B. Keratin 20 serine 13 phosphorylation is a stress and intestinal goblet cell marker. J. Biol. Chem. 2006, 281, 16453–16461. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.W.; Wong, N.A.; Liu, Y.; Bicknell, D.; Turley, H.; Hollins, L.; Miller, C.J.; Wilding, J.L.; Bodmer, W.F. Gastrointestinal differentiation marker cytokeratin 20 is regulated by homeobox gene CDX1. Proc. Natl. Acad. Sci. USA 2009, 106, 1936–1941. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zhou, Y.; Rychahou, P.; Fan, T.W.; Lane, A.N.; Weiss, H.L.; Evers, B.M. Ketogenesis contributes to intestinal cell differentiation. Cell Death Differ. 2017, 24, 458–468. [Google Scholar] [CrossRef] [Green Version]
- Kien, C.L.; Schmitz-Brown, M.; Solley, T.; Sun, D.; Frankel, W.L. Increased colonic luminal synthesis of butyric acid is associated with lowered colonic cell proliferation in piglets. J. Nutr. 2006, 136, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Herosimczyk, A.; Lepczyński, A.; Ożgo, M.; Barszcz, M.; Marynowska, M.; Tuśnio, A.; Taciak, M.; Markulen, A.; Skomiał, J. Proteome changes in ileal mucosa of young pigs resulting from different levels of native chicory inulin in the diet. J. Anim. Feed Sci. 2018, 27, 229–237. [Google Scholar] [CrossRef]
- Uchiyama, K.; Naito, Y.; Takagi, T.; Mizushima, K.; Hirai, Y.; Hayashi, N.; Harusato, A.; Inoue, K.; Fukumoto, K.; Yamada, S.; et al. Serpin B1 protects colonic epithelial cell via blockage of neutrophil elastase activity and its expression is enhanced in patients with ulcerative colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G1163–G1170. [Google Scholar] [CrossRef]
- Duncan, R.; Carpenter, B.; Main, L.C.; Telfer, C.; Murray, G.I. Characterisation and protein expression profiling of annexins in colorectal cancer. Br. J. Cancer 2008, 98, 426–433. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.Y.; Wei, P.L.; Chen, W.Y.; Chang, W.C.; Chang, Y.J. Silencing heat shock protein 27 inhibits the progression and metastasis of colorectal cancer (CRC) by maintaining the stability of stromal interaction molecule 1 (STIM1) proteins. Cells 2018, 7, 262. [Google Scholar] [CrossRef] [Green Version]
- Janda, K.; Gutowska, I.; Geszke-Moritz, M.; Jakubczyk, K. The common chicory (Cichorium intybus L.) as a source of extracts with health-promoting properties—A review. Molecules 2021, 26, 1814. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Diet | C Group | CR Group |
|---|---|---|
| Composition (g/kg) | ||
| Wheat | 468.40 | 458.40 |
| Barley | 200.00 | 200.00 |
| Corn starch | 30.00 | 0.00 |
| Full-fat soy bean | 59.00 | 59.00 |
| Whey | 97.00 | 97.00 |
| Fish meal | 40.00 | 40.00 |
| Spray-dried blood plasma | 40.00 | 40.00 |
| Soybean oil | 34.00 | 34.00 |
| Calcium formate | 3.00 | 3.00 |
| Calcium carbonate | 5.00 | 5.00 |
| Calcium monophosphate | 6.00 | 6.00 |
| Sodium chloride | 0.70 | 0.70 |
| L-Lysine HCL (78.5%) | 6.10 | 6.10 |
| DL-Methionine (99%) | 2.30 | 2.30 |
| L-Threonine (98%) | 2.60 | 2.60 |
| L-Tryptophan (98%) | 0.90 | 0.90 |
| Mineral–vitamin mix 1 | 4.00 | 4.00 |
| Aroma | 1.00 | 1.00 |
| Dried chicory roots | 0.00 | 40.00 |
| Dry mater (%) and nutrient (% dry matter) 2 | ||
| Dry matter | 90.03 | 90.15 |
| Crude ash | 4.54 | 4.55 |
| Crude protein | 20.05 | 20.05 |
| Ether extract | 6.04 | 6.04 |
| Crude fibre | 1.52 | 1.53 |
| Fructans | 1.00 | 2.74 |
| ME (MJ/kg) 3 | 14.30 | 14.20 |
| Item | CR 1 |
|---|---|
| Dry matter | 92.67 |
| Crude protein | 5.63 |
| Crude ash | 3.61 |
| Ether extract | 0.34 |
| Crude fiber | 5.56 |
| Fructans | 51.56 |
| Parameter | C | CR 1 | SEM | p-Value |
|---|---|---|---|---|
| Initial body weight (at weaning), kg | 8.00 | 8.60 | 0.26 | 0.256 |
| Average daily feed intake, kg/day | 0.53 | 0.78 | 0.075 | - |
| Final body weight, kg | 17.9 | 19.1 | 0.72 | 0.427 |
| Total body weight gain, kg | 9.9 | 10.5 | 0.58 | 0.628 |
| Average daily body weight gain, kg/day | 0.47 | 0.50 | 0.028 | 0.624 |
| Spot No. | Gene Name 3 | MVM 4/ MS 5 | SC 6 | Theoretical pI/Mw (pH/kDa) 7 | Experimental pI/Mw (pH/kDa) 8 | Taxonomy |
|---|---|---|---|---|---|---|
| 1 | CALR 1 | 8/92 | 34 | 4.32/48.43 | 3.70/107.60 | Sus scrofa |
| 2 | CALR 1 | 8/84 | 30 | 4.32/48.43 | 3.70/91.90 | Sus scrofa |
| 3 | GSN 1 | 6/67 | 15 | 5.93/85.06 | 4.60/105.00 | Sus scrofa |
| 4 | GSN 1 | 6/67 | 15 | 5.93/85.06 | 4.70/105.40 | Sus scrofa |
| 5 | LCP1 1 | 10/81 | 24 | 5.29/70.65 | 4.80/119.80 | Sus scrofa |
| 6 | MX1 1 | 6/89 | 15 | 5.45/76.27 | 4.90/124.10 | Sus scrofa |
| 7 | ARHGDIA 1 | 8/97 | 40 | 5.12/23.44 | 5.10/119.50 | Sus scrofa |
| 8 | MVP 1 | 17/137 | 31 | 5.53/99.98 | 5.30/124.40 | Sus scrofa |
| 9 | HSPCA 1 | 11/105 | 23 | 4.93/85.12 | 5.40/111.30 | Sus scrofa |
| 10 | HSP70.2 1 | 10/106 | 26 | 5.60/70.43 | 5.50/105.30 | Sus scrofa |
| 11 | CCT5 1 | 7/88 | 29 | 5.22/53.64 | 5.70/96.50 | Pan troglodytes |
| 12 | GSN 1 | 14/159 | 31 | 5.93/85.06 | 6.30/134.80 | Sus scrofa |
| 13 | EEF1G 1 | 7/90 | 31 | 6.16/49.93 | 6.60/129.20 | Sus scrofa |
| 14 | YWHAQ 1 | 10/112 | 65 | 4.37/18.36 | 6.30/114.80 | Sus scrofa |
| 15 | HSP8 1 | 8/80 | 24 | 5.38/71.10 | 6.60/107.30 | Chrysochloris asiatica |
| 16 | SARS1 1 | 7/65 | 22 | 5.86/59.06 | 6.80/98.00 | Rattus norvegicus |
| 17 | DPYSL2 1 | 9/78 | 27 | 5.95/62.64 | 6.90/100.30 | Bos taurus |
| 18 | PRPF19 1 | 8/82 | 26 | 6.14/55.60 | 7.00/93.30 | Sus scrofa |
| 19 | GFPT1 1 | 8/69 | 16 | 6.39/79.29 | 7.40/114.80 | Mus musculus |
| 20 | MSN 1 | 34/259 | 44 | 6.18/68.02 | 7.50/108.90 | Sus scrofa |
| 21 | MSN 1 | 15/131 | 28 | 6.18/68.02 | 7.60/108.60 | Sus scrofa |
| 22 | TF 1 | 9/80 | 20 | 6.73/78.95 | 7.70/111.60 | Sus scrofa |
| 23 | TF 1 | 10/70 | 20 | 6.93/78.97 | 8.20/111.30 | Sus scrofa |
| 24 | LMNA 1 | 28/241 | 49 | 6.40/65.19 | 8.20/107.50 | Sus scrofa |
| 25 | EEF2 1 | 43/396 | 56 | 6.41/96.26 | 8.30/120.70 | Sus scrofa |
| 26 | SND1 1 | 23/201 | 28 | 6.72/102.52 | 8.40/125.30 | Sus scrofa |
| 27 | HNRNPLL 1 | 15/135 | 26 | 8.46/64,57 | 8.50/100.10 | Sus scrofa |
| 28 | C3 1 | 31/208 | 24 | 5.99/193.05 | 8.60/101.50 | Sus scrofa |
| 29 | HNRNPK 1 | 9/76 | 28 | 5.39/51.23 | 4.70/97.00 | Sus scrofa |
| 30 | KRT20 1 | 16/176 | 52 | 5.38/49.09 | 4.90/85.20 | Sus scrofa |
| 31 | TUBA1B 1 | 9/93 | 35 | 4.94/50.80 | 5.00/92.50 | Sus scrofa |
| 32 | TUBA1B 1 | 10/85 | 33 | 4.94/50.87 | 5.20/92.20 | Rousettus aegyptiacus |
| 33 | ACTR3 1 | 19/130 | 57 | 5.61/47.80 | 5.60/83.00 | Bos taurus |
| 34 | ALDH9A1 1 | 12/62 | 29 | 5.84/54.82 | 5.70/83.00 | Sus scrofa |
| 35 | VCP 1 | 14/127 | 31 | 5.13/89.92 | 6.10/78.10 | Sus scrofa |
| 36 | ENO1 1 | 11/80 | 28 | 6.44/47.60 | 7.00/86.80 | Sus scrofa |
| 37 | EEF1G 1 | 10/130 | 38 | 6.16/49.93 | 7.20/88.50 | Sus scrofa |
| 38 | SELENBP1 1 | 18/161 | 41 | 5.93/52.93 | 7.20/88.50 | Homo sapiens |
| 39 | G6PD 1 | 8/93 | 29 | 6.27/59.71 | 7.30/86.30 | Sus scrofa |
| 40 | CAP1 1 | 8/62 | 23 | 7.64/51.88 | 7.70/92.20 | Macaca fascicularis |
| 41 | CCT7 1 | 11/64 | 26 | 6.78/59.92 | 8.30/92.20 | Bos taurus |
| 42 | UDPGDH1 1 | 16/124 | 42 | 6.73/55.72 | 8.30/95.00 | Sus scrofa |
| 43 | UDPGDH1 1 | 14/109 | 39 | 6.73/55.72 | 8.50/92.00 | Sus scrofa |
| 44 | CAP1 1 | 11/109 | 27 | 6.84/51.56 | 8.30/89.30 | Sus scrofa |
| 45 | CAP1 1 | 8/95 | 24 | 7.62/51.77 | 8.30/86.00 | Sus scrofa |
| 46 | EEF1G 1 | 17/178 | 52 | 6.16/49.93 | 7.00/80.40 | Sus scrofa |
| 47 | EEF1G 1 | 17/145 | 56 | 6.15/50.38 | 7.10/74.50 | Sus scrofa |
| 48 | SERPINB1 1 | 14/142 | 43 | 6.13/42.66 | 7.00/66.20 | Sus scrofa |
| 49 | ACTR1A 1 | 5/64 | 30 | 6.19/42.70 | 7.30/77.10 | Canis lupus familiaris |
| 50 | SEPTIN2 1 | 7/64 | 34 | 6.15/41.69 | 7.30/77.10 | Homo sapiens |
| 51 | ZFP42 1 | 8/75 | 31 | 9.12/35.52 | 7.70/76.40 | Homo sapiens |
| 52 | IDH1 1 | 14/140 | 48 | 6.48/47.15 | 7.80/73.00 | Sus scrofa |
| 53 | MAPK1 1 | 12/169 | 42 | 6.50/41.68 | 7.80/65.90 | Sus scrofa |
| 54 | AKR1A1 1 | 9/91 | 39 | 6.51/36.84 | 8.10/61.40 | Sus scrofa |
| 55 | PCBP1 1 | 16/184 | 60 | 6.66/37.99 | 8.20/66.50 | Sus scrofa |
| 56 | ANXA2 2 | 39/81 | 77 | 6.49/38.79 | 8.30/53.60 | Sus scrofa |
| 57 | AKR1A1 2 | 26/75 | 64 | 6.51/36.84 | 8.30/58.80 | Sus scrofa |
| 58 | 6PGD 1 | 24/207 | 50 | 6.48/53.66 | 8.40/79.50 | Sus scrofa |
| 59 | CS 1 | 17/126 | 38 | 6.92/48.90 | 8.60/74.20 | Sus scrofa |
| 60 | CKMT1 1 | 20/222 | 47 | 8.60/47.30 | 8.70/71.60 | Sus scrofa |
| 61 | IPP1 1 | 7/90 | 43 | 5.44/33.25 | 4.00/55.50 | Sus scrofa |
| 62 | ATP5F1B 1 | 10/87 | 28 | 5.15/56.25 | 4.80/48.80 | Bos taurus |
| 63 | ACT 2 | 18/50 | 66 | 5.12/42.52 | 5.40/58.80 | Molgula oculata |
| 64 | ANXA4 2 | 33/71 | 76 | 5.10/48.40 | 5.60/55.30 | Sus scrofa |
| 65 | PSME2 1 | 5/62 | 33 | 5.41/27.45 | 5.40/46.40 | Sus scrofa |
| 66 | PHB 1 | 8/97 | 48 | 5.57/29.86 | 5.50/44.90 | Homo sapiens |
| 67 | KRT77 2 | 52/64 | 74 | 5.48/57.39 | 6.60/53.50 | Rattus norvegicus |
| 68 | ERP29 1 | 8/84 | 36 | 6.85/29.32 | 7.20/46.80 | Sus scrofa |
| 69 | HSP27 2 | 5/63 | 30 | 6.23/22.98 | 7.20/45.80 | Sus scrofa |
| 70 | TST 1 | 16/164 | 56 | 6.72/33.65 | 8.50/47.70 | Sus scrofa |
| 71 | TST 1 | 19/188 | 52 | 6.72/33.65 | 8.60/47.60 | Sus scrofa |
| 72 | GNB2L1 1 | 18/249 | 78 | 7.60/35.51 | 8.70/47.00 | Sus scrofa |
| 73 | PGAM1 1 | 13/151 | 68 | 6.67/28.90 | 8.10/43.30 | Homo sapiens |
| 74 | TPI1 2 | 11/147 | 58 | 6.54/26.88 | 8.30/41.70 | Sus scrofa |
| 75 | MYL12B 1 | 7/80 | 50 | 4.66/19.85 | 4.00/23.80 | Echinops telfairi |
| 76 | CFL1 1 | 9/121 | 62 | 8.16/18.79 | 9.20/20.20 | Sus scrofa |
| Spot No. | Gene Name 3 | Ratio CR/C 4 | p-Value |
|---|---|---|---|
| up-regulated protein spot | |||
| 30 | KRT20 1 | 2.23 | <0.001 * |
| down-regulated protein spots | |||
| 18 | PRPF19 1 | 0.30 | <0.001 * |
| 33 | ACTR3 1 | 0.46 | 0.003 ** |
| 34 | ALDH9A1 1 | 0.50 | 0.047 ** |
| 39 | G6PD 1 | 0.18 | <0.001 ** |
| 44 | CAP1 1 | 0.29 | <0.001 * |
| 46 | EEF1G 1 | 0.37 | 0.016 ** |
| 48 | SERPINB1 1 | 0.31 | <0.001 * |
| 56 | ANXA2 2 | 0.33 | <0.001 ** |
| 57 | AKR1A1 2 | 0.12 | <0.001 ** |
| 60 | CKMT1 1 | 0.43 | 0.004 ** |
| 63 | ACT 2 | 0.41 | 0.001 * |
| 64 | ANXA4 2 | 0.48 | 0.001 * |
| 67 | KRT77 2 | 0.41 | 0.002 * |
| 69 | HSP27 2 | 0.29 | <0.001 * |
| 74 | TPI1 2 | 0.13 | <0.001 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herosimczyk, A.; Lepczyński, A.; Werkowska, M.; Barszcz, M.; Taciak, M.; Tuśnio, A.; Ciechanowicz, A.K.; Kucia, M.; Susfał, K.; Cabała, S.; et al. Dietary Inclusion of Dried Chicory Root Affects Cecal Mucosa Proteome of Nursery Pigs. Animals 2022, 12, 1710. https://doi.org/10.3390/ani12131710
Herosimczyk A, Lepczyński A, Werkowska M, Barszcz M, Taciak M, Tuśnio A, Ciechanowicz AK, Kucia M, Susfał K, Cabała S, et al. Dietary Inclusion of Dried Chicory Root Affects Cecal Mucosa Proteome of Nursery Pigs. Animals. 2022; 12(13):1710. https://doi.org/10.3390/ani12131710
Chicago/Turabian StyleHerosimczyk, Agnieszka, Adam Lepczyński, Martyna Werkowska, Marcin Barszcz, Marcin Taciak, Anna Tuśnio, Andrzej Krzysztof Ciechanowicz, Magdalena Kucia, Karolina Susfał, Sandra Cabała, and et al. 2022. "Dietary Inclusion of Dried Chicory Root Affects Cecal Mucosa Proteome of Nursery Pigs" Animals 12, no. 13: 1710. https://doi.org/10.3390/ani12131710