
Identification of Estrus in Sows Based on Salivary Proteomics

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Animals and Sample Collection
2.2. Sample Digestion and Enzymatic Hydrolysis
2.3. Mass Spectrometry Analysis
2.4. Database Retrieval, Protein Identification, and Quantification
2.5. Data Analysis
3. Results
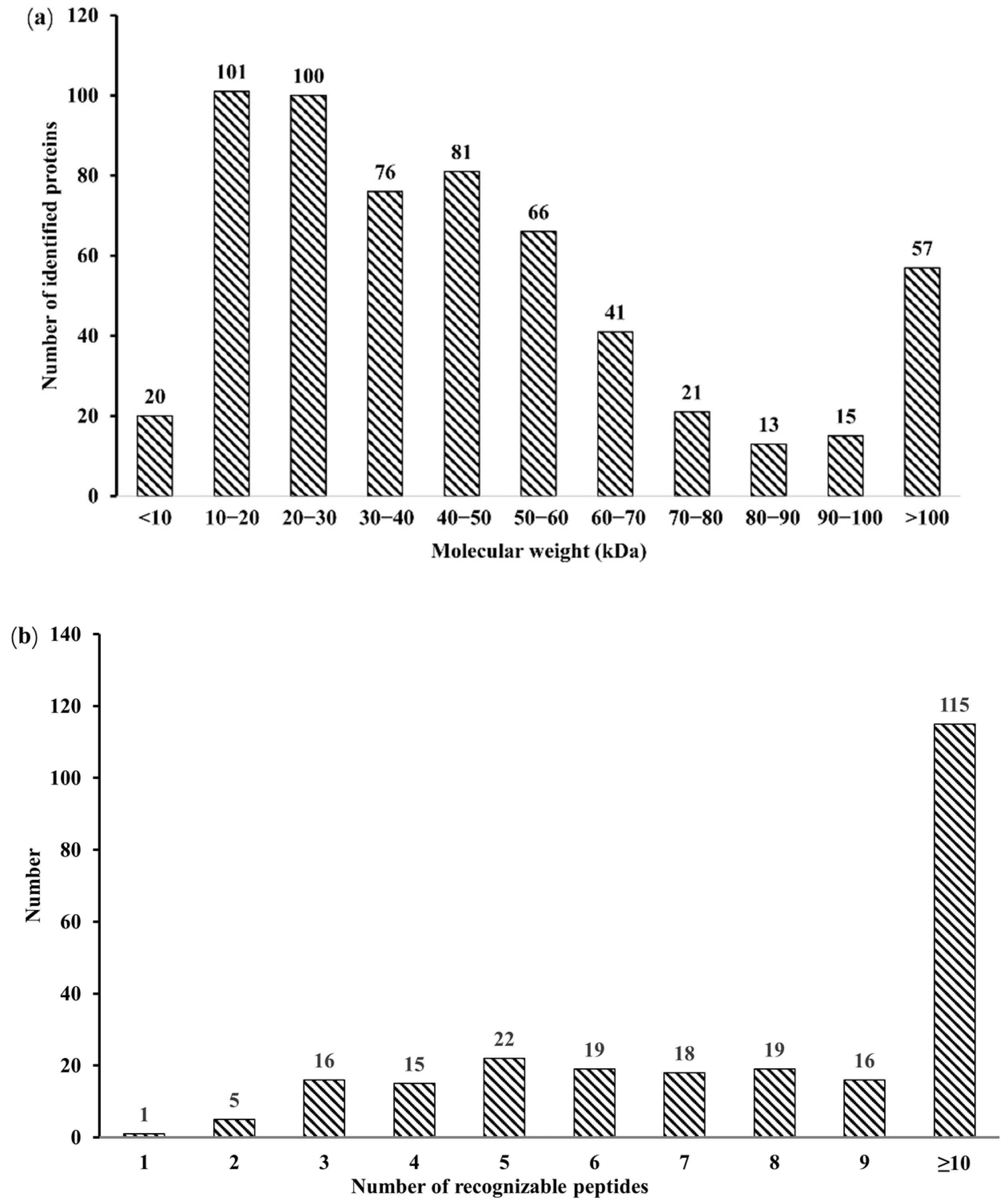
3.1. Identification of Salivary Proteins in the Oestrus Cycle of Large White Sows
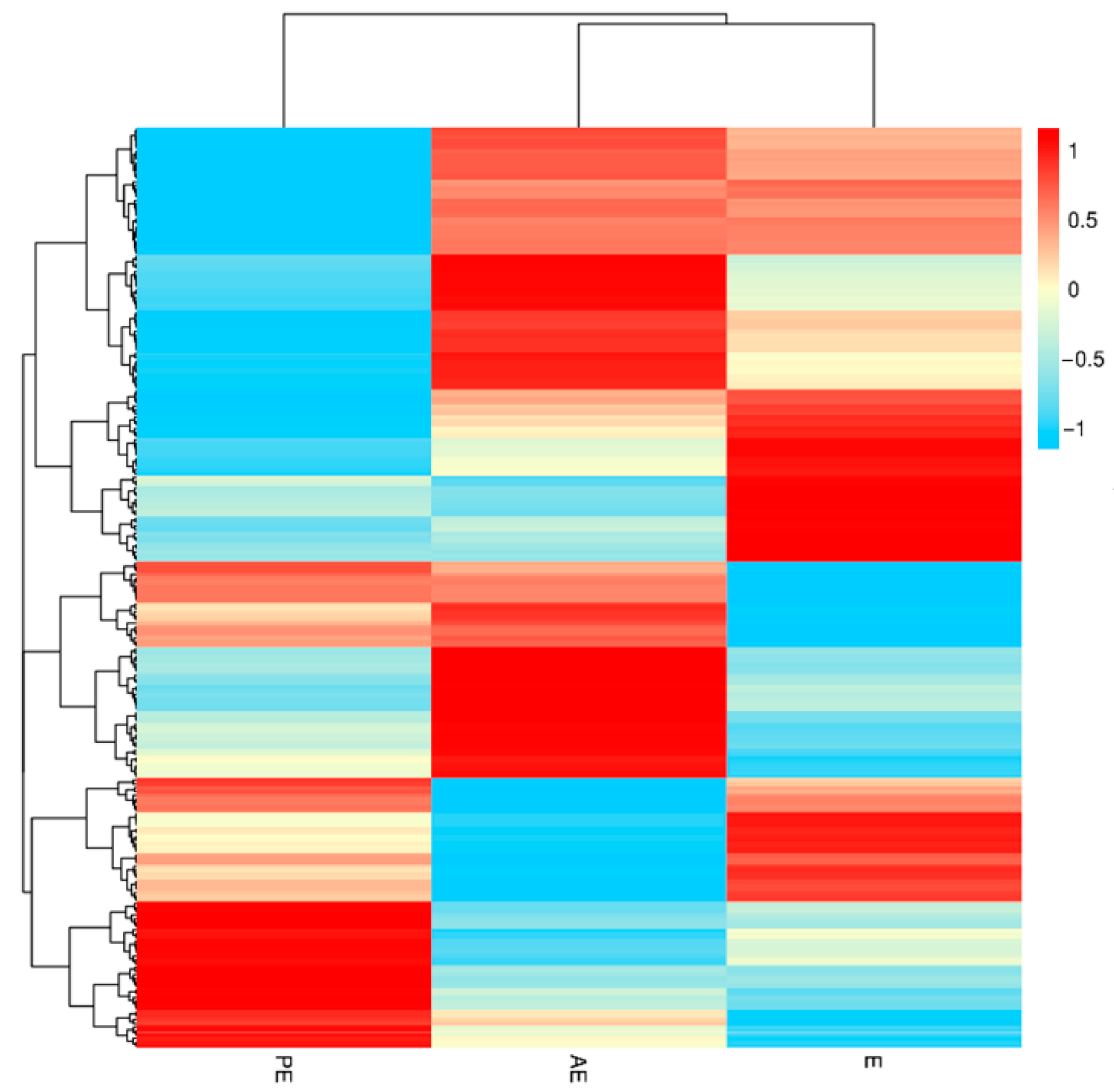
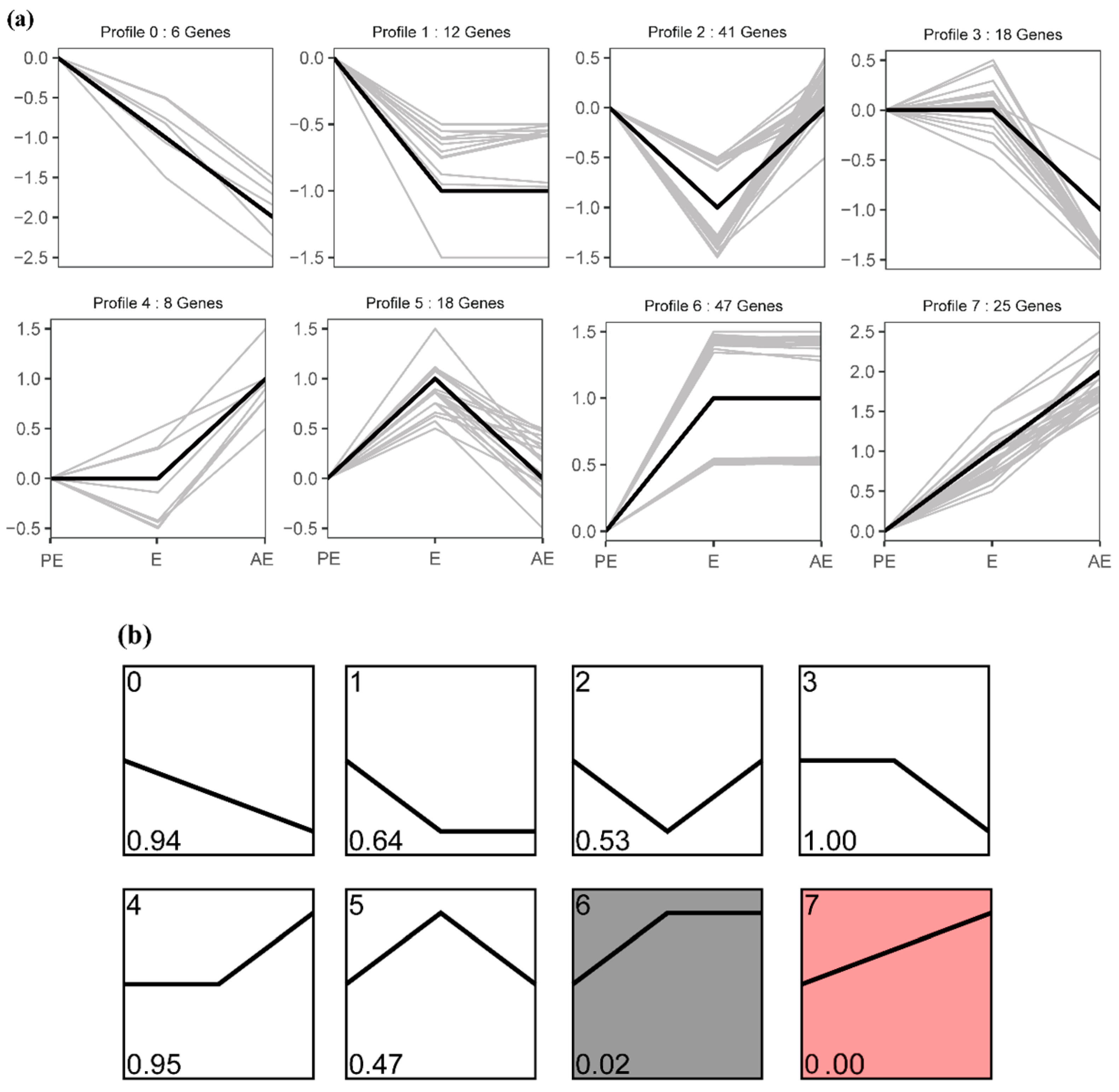
3.2. Patterns of the Temporal Shifts of Salivary Proteins in the Oestrus Cycle of Large White Sows
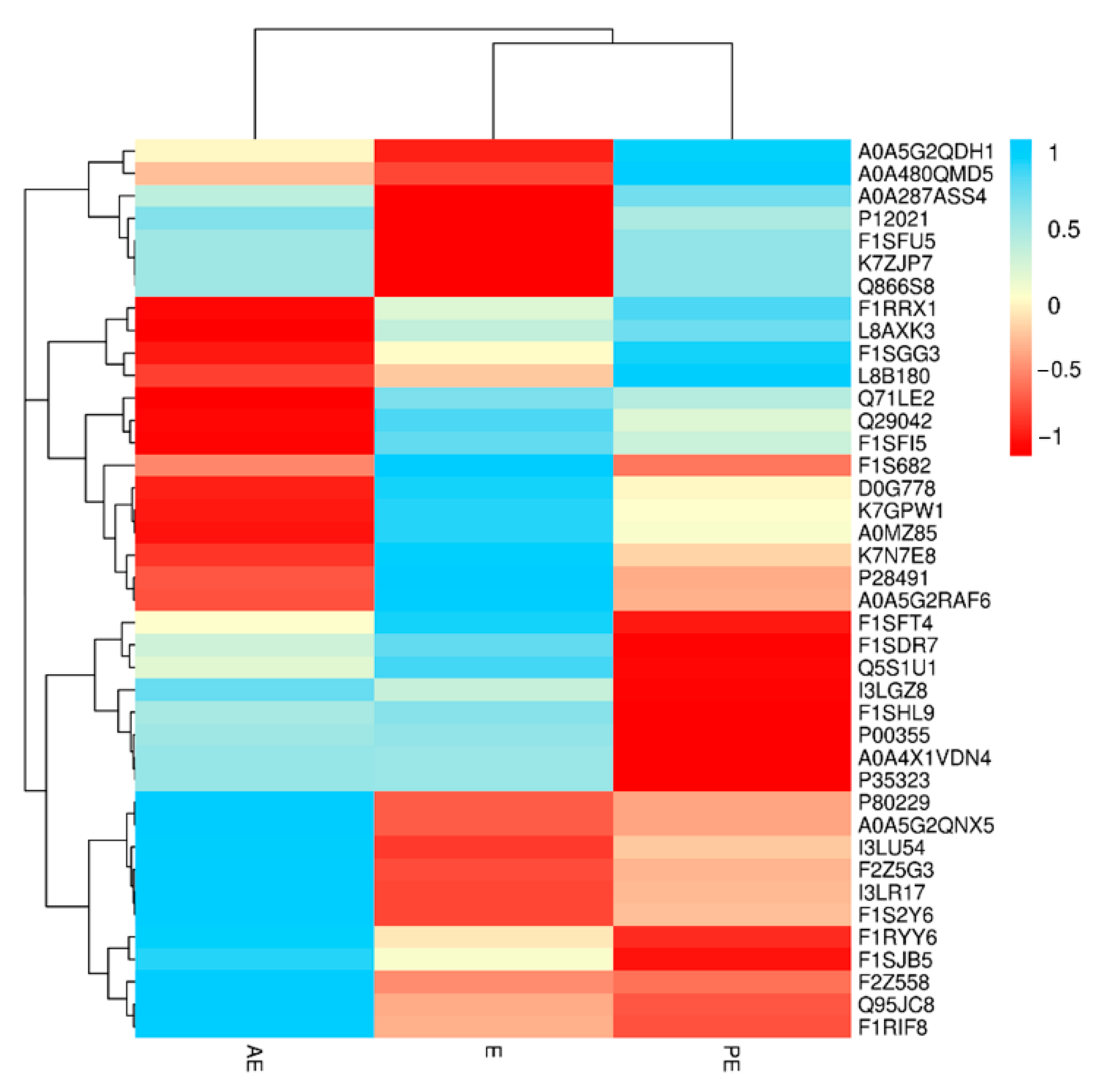
3.3. Screening and Analysis of Salivary Differential Proteins in the Oestrus Cycle of Large White Sows
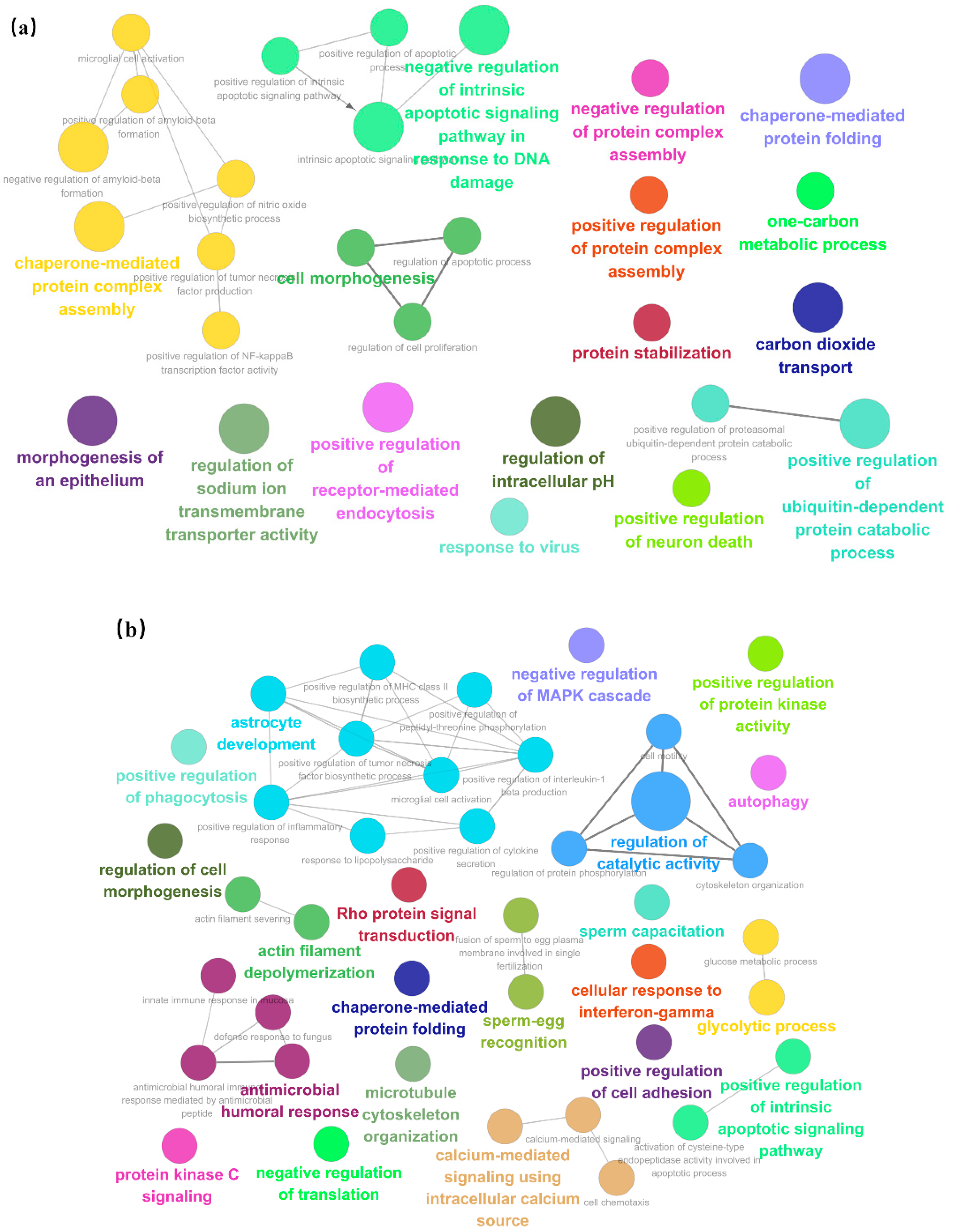
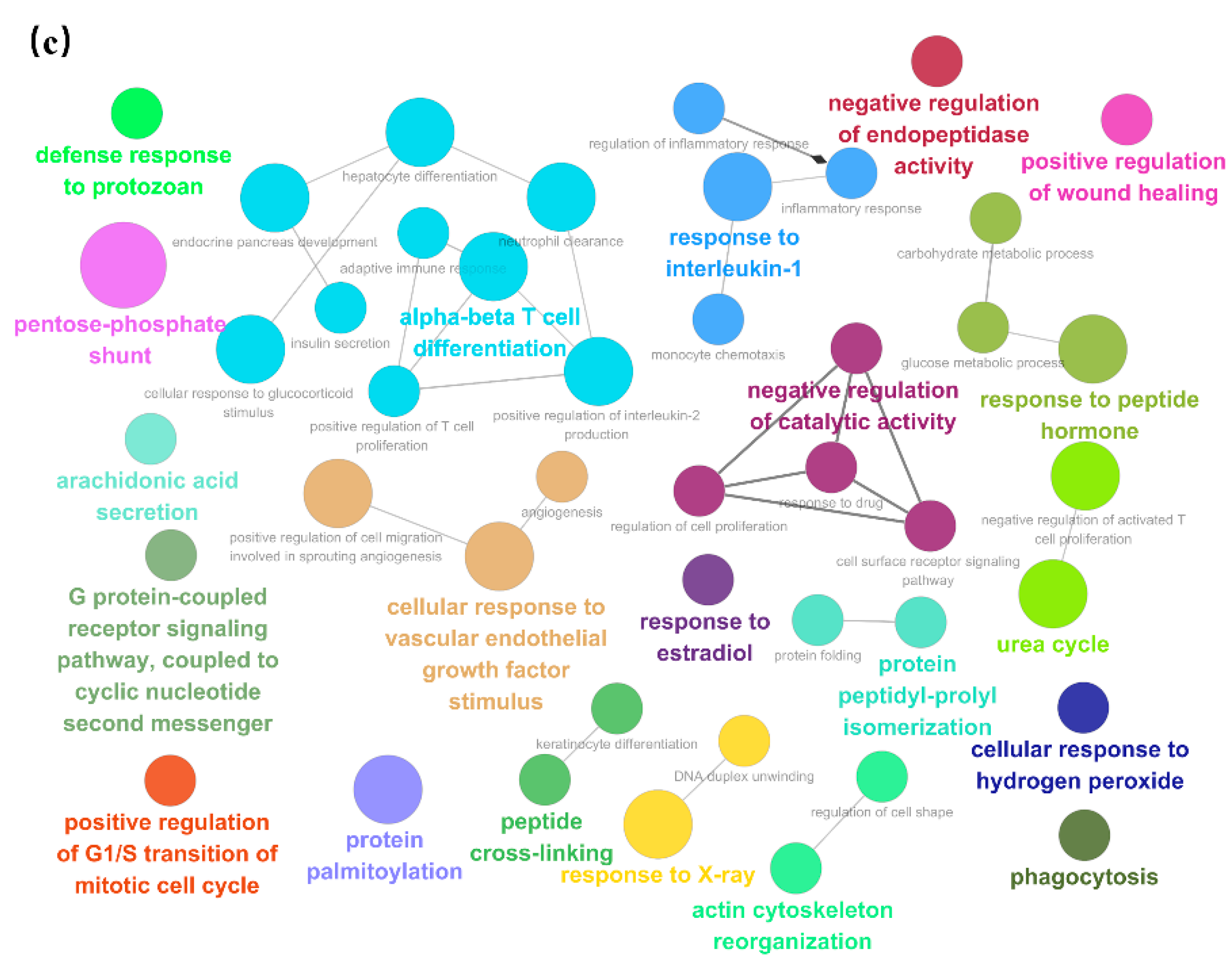
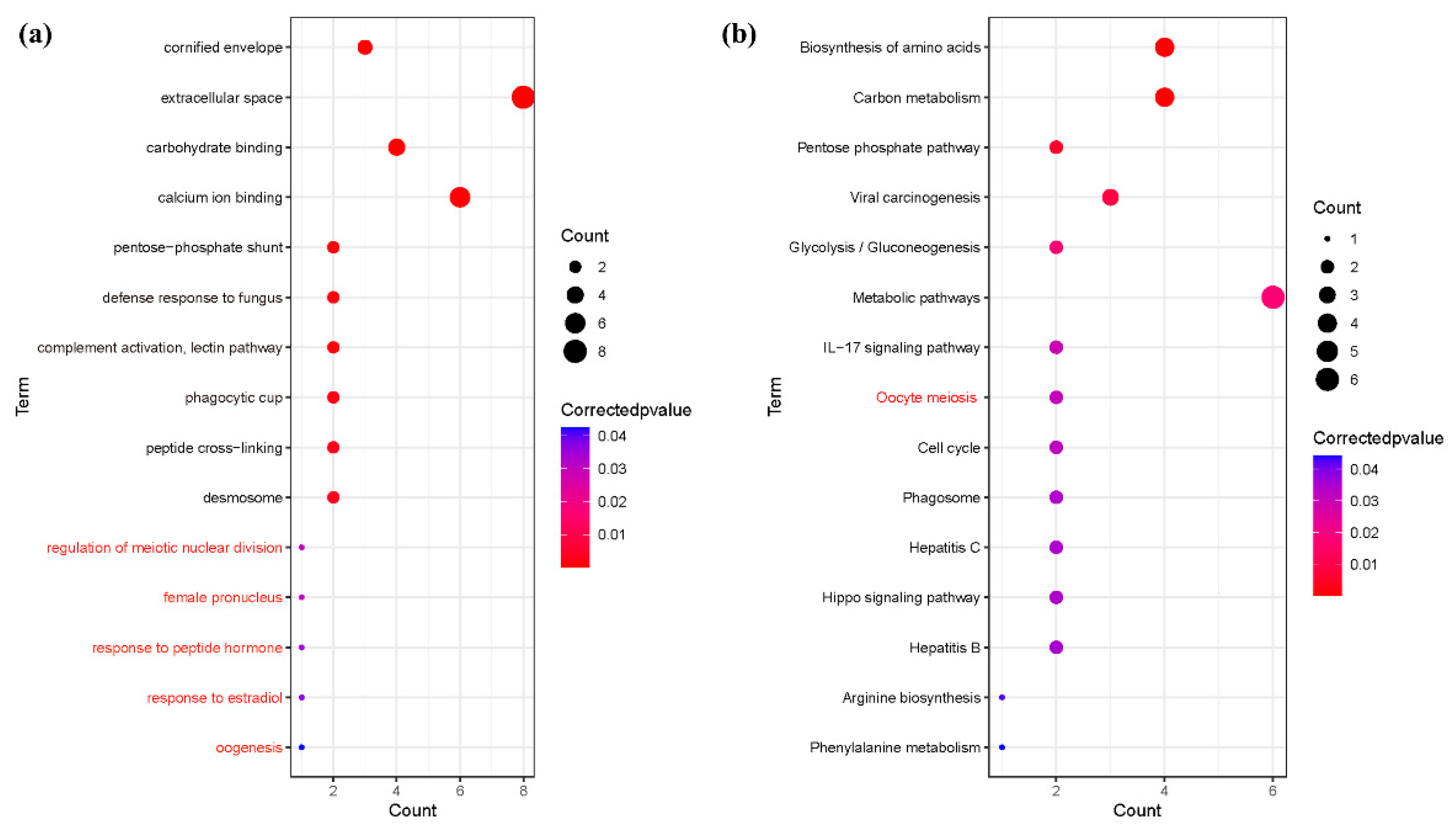
3.4. Enrichment of Salivary Fifferential Proteins in the Estrus Cycle of Large White Sows Based on GO and KEGG Analysis
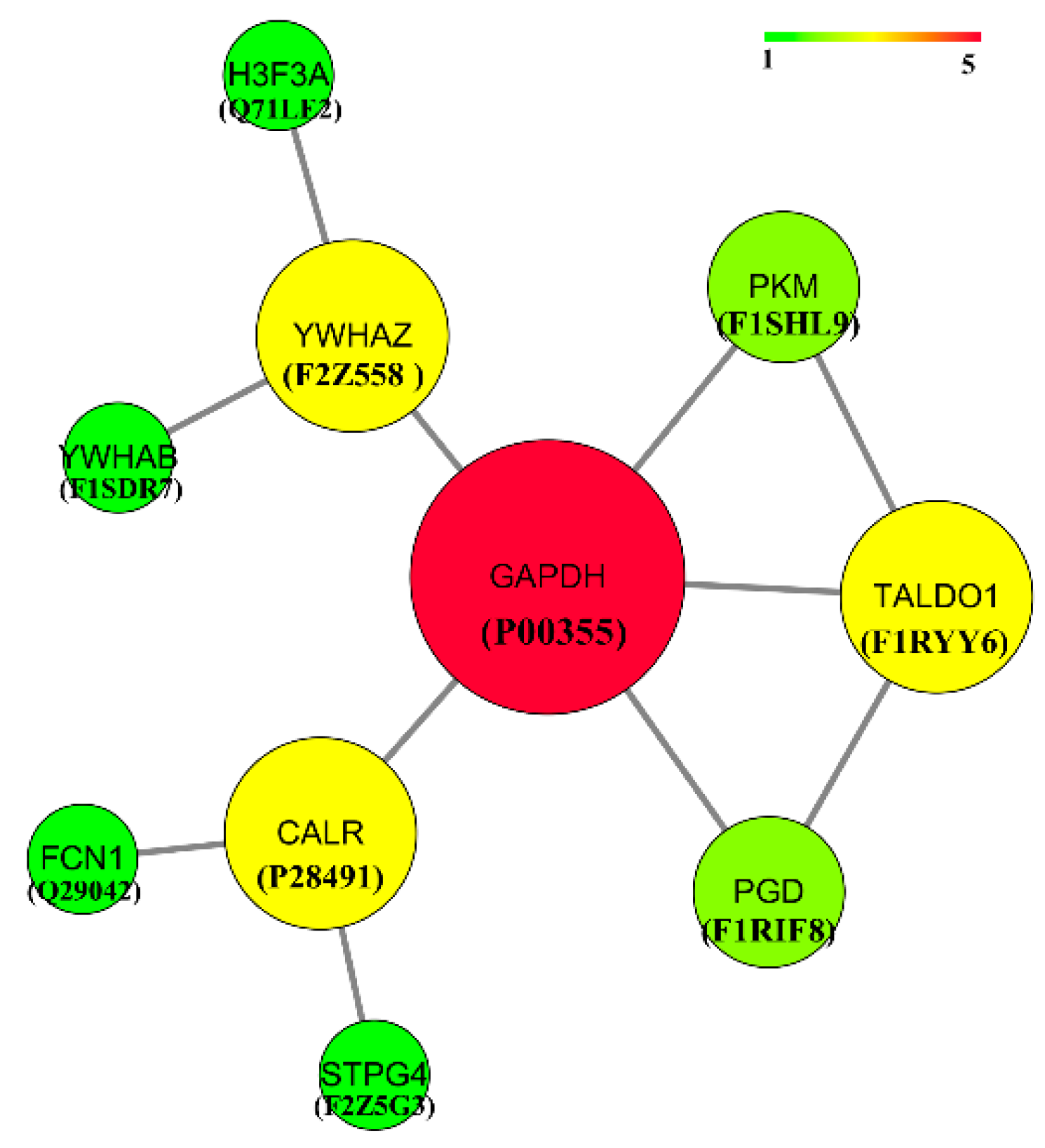
3.5. Protein Interaction Network Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rasbech, N.O. A review of the causes of reproductive failure in swine. Br. Vet. J. 1969, 125, 599–616. [Google Scholar] [CrossRef]
- Alagendran, S.; Saibaba, G.; Muthukumar, S.; Rajkumar, R.; Guzman, R.G.; Archunan, G. Characterization of salivary protein during ovulatory phase of menstrual cycle through MALDI-TOF/MS. Indian J. Dent. Res. 2013, 24, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, J.; Donadeu, F.X. Circulating microRNA Profiles during the Bovine Oestrous Cycle. PLoS ONE 2016, 11, e0158160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achiraman, S.; Archunan, G.; Sankarganesh, D.; Rajagopal, T.; Rengarajan, R.L.; Kokilavani, P.; Kamalakkannan, S.; Kannan, S. Biochemical analysis of female mice urine with reference to endocrine function: A key tool for estrus detection. Zool. Sci. 2011, 28, 600–605. [Google Scholar] [CrossRef]
- Zhang, L.; Farrell, J.J.; Zhou, H.; Elashoff, D.; Akin, D.; Park, N.H.; Chia, D.; Wong, D.T. Salivary transcriptomic biomarkers for detection of resectable pancreatic cancer. Gastroenterology 2010, 138, 949–957.e1–7. [Google Scholar] [CrossRef] [Green Version]
- Yoshizawa, J.M.; Wong, D.T. Salivary microRNAs and oral cancer detection. Methods Mol. Biol. 2013, 936, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Wong, D.T. Salivaomics. J. Am. Dent. Assoc. 2012, 143, 19s–24s. [Google Scholar] [CrossRef]
- Denny, P.; Hagen, F.K.; Hardt, M.; Liao, L.; Yan, W.; Arellanno, M.; Bassilian, S.; Bedi, G.S.; Boontheung, P.; Cociorva, D.; et al. The proteomes of human parotid and submandibular/sublingual gland salivas collected as the ductal secretions. J. Proteome Res. 2008, 7, 1994–2006. [Google Scholar] [CrossRef] [Green Version]
- Khurshid, Z.; Zafar, M.S.; Khan, R.S.; Najeeb, S.; Slowey, P.D.; Rehman, I.U. Role of Salivary Biomarkers in Oral Cancer Detection. Adv. Clin. Chem. 2018, 86, 23–70. [Google Scholar] [CrossRef]
- Hofman, L.F. Human saliva as a diagnostic specimen. J. Nutr. 2001, 131, 1621s–1625s. [Google Scholar] [CrossRef]
- Kittawornrat, A.; Prickett, J.; Wang, C.; Olsen, C.; Irwin, C.; Panyasing, Y.; Ballagi, A.; Rice, A.; Main, R.; Johnson, J.; et al. Detection of Porcine reproductive and respiratory syndrome virus (PRRSV) antibodies in oral fluid specimens using a commercial PRRSV serum antibody enzyme-linked immunosorbent assay. J. Vet. Diagn. Investig. 2012, 24, 262–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romagosa, A.; Gramer, M.; Joo, H.S.; Torremorell, M. Sensitivity of oral fluids for detecting influenza A virus in populations of vaccinated and non-vaccinated pigs. Influenza Other Respir. Viruses 2012, 6, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Grau, F.R.; Schroeder, M.E.; Mulhern, E.L.; McIntosh, M.T.; Bounpheng, M.A. Detection of African swine fever, classical swine fever, and foot-and-mouth disease viruses in swine oral fluids by multiplex reverse transcription real-time polymerase chain reaction. J. Vet. Diagn. Investig. 2015, 27, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Prickett, J.R.; Zimmerman, J.J. The development of oral fluid-based diagnostics and applications in veterinary medicine. Anim. Health Res. Rev. 2010, 11, 207–216. [Google Scholar] [CrossRef]
- Pietschmann, J.; Mur, L.; Blome, S.; Beer, M.; Pérez-Sánchez, R.; Oleaga, A.; Sánchez-Vizcaíno, J.M. African swine fever virus transmission cycles in Central Europe: Evaluation of wild boar-soft tick contacts through detection of antibodies against Ornithodoros erraticus saliva antigen. BMC Vet. Res. 2016, 12, 1. [Google Scholar] [CrossRef] [Green Version]
- Saleh, L.H. The use of saliva for the detection of IgG and anti-bodies against rubella virus: Comparison of indirect ELISA and antibody capture immunoassay. J. Egypt Public Health Assoc. 1991, 66, 123–134. [Google Scholar]
- Mortimer, P.P.; Parry, J.V. Detection of antibody to HIV in saliva: A brief review. Clin. Diagn. Virol. 1994, 2, 231–243. [Google Scholar] [CrossRef]
- Hashida, S.; Hashinaka, K.; Ishikawa, S.; Ishikawa, E. More reliable diagnosis of infection with human immunodeficiency virus type 1 (HIV-1) by detection of antibody IgGs to pol and gag proteins of HIV-1 and p24 antigen of HIV-1 in urine, saliva, and/or serum with highly sensitive and specific enzyme immunoassay (immune complex transfer enzyme immunoassay): A review. J. Clin. Lab. Anal. 1997, 11, 267–286. [Google Scholar] [CrossRef]
- Michalke, B.; Rossbach, B.; Göen, T.; Schäferhenrich, A.; Scherer, G. Saliva as a matrix for human biomonitoring in occupational and environmental medicine. Int. Arch. Occup. Environ. Health 2015, 88, 1–44. [Google Scholar] [CrossRef]
- Lindsay, A.; Costello, J.T. Realising the Potential of Urine and Saliva as Diagnostic Tools in Sport and Exercise Medicine. Sports Med. 2017, 47, 11–31. [Google Scholar] [CrossRef] [Green Version]
- Chojnowska, S.; Baran, T.; Wilińska, I.; Sienicka, P.; Cabaj-Wiater, I.; Knaś, M. Human saliva as a diagnostic material. Adv. Med. Sci. 2018, 63, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Golatowski, C.; Salazar, M.G.; Dhople, V.M.; Hammer, E.; Kocher, T.; Jehmlich, N.; Völker, U. Comparative evaluation of saliva collection methods for proteome analysis. Clin. Chim. Acta 2013, 419, 42–46. [Google Scholar] [CrossRef] [PubMed]
- Henson, B.S.; Wong, D.T. Collection, storage, and processing of saliva samples for downstream molecular applications. Methods Mol. Biol. 2010, 666, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Nunes, L.A.; Mussavira, S.; Bindhu, O.S. Clinical and diagnostic utility of saliva as a non-invasive diagnostic fluid: a systematic review. Biochem. Med. 2015, 25, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Krzymowski, T.; Stefańczyk, S.; Krzymowska, H.; Lipski, M.; Olejniczak, J. Testosterone, estradiol and progesterone in sow’s follicular fluid during different stages of estrous cycle. Bull. Acad. Pol. Sci. Biol. 1979, 27, 417–421. [Google Scholar] [PubMed]
- Sandberg, A.; Lindell, G.; Källström, B.N.; Branca, R.M.; Danielsson, K.G.; Dahlberg, M.; Larson, B.; Forshed, J.; Lehtiö, J. Tumor proteomics by multivariate analysis on individual pathway data for characterization of vulvar cancer phenotypes. Mol. Cell. Proteom. 2012, 11, M112.016998. [Google Scholar] [CrossRef] [Green Version]
- Ernst, J.; Bar-Joseph, Z. STEM: A tool for the analysis of short time series gene expression data. BMC Bioinform. 2006, 7, 191. [Google Scholar] [CrossRef] [Green Version]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Ing, N.H.; Zhang, Y. Cell-specific expression of estrogen-responsive genes in the uteri of cyclic, early pregnant and ovariectomized ewes. Theriogenology 2004, 62, 403–414. [Google Scholar] [CrossRef]
- Ramirez, V.D.; Kipp, J.L.; Joe, I. Estradiol, in the CNS, targets several physiologically relevant membrane-associated proteins. Brain Res. Brain Res. Rev. 2001, 37, 141–152. [Google Scholar] [CrossRef]
- Joe, I.; Ramirez, V.D. Binding of estrogen and progesterone-BSA conjugates to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and the effects of the free steroids on GAPDH enzyme activity: Physiological implications. Steroids 2001, 66, 529–538. [Google Scholar] [CrossRef]
- Zhang, D.X.; Li, X.P.; Sun, S.C.; Shen, X.H.; Cui, X.S.; Kim, N.H. Involvement of ER-calreticulin-Ca2+ signaling in the regulation of porcine oocyte meiotic maturation and maternal gene expression. Mol. Reprod. Dev. 2010, 77, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Ramos, E.; Sandoval-Hernández, A.; Tecalco-Cruz, A.C. Differential expression and molecular interactions of chromosome region maintenance 1 and calreticulin exportins in breast cancer cells. J. Steroid Biochem. Mol. Biol. 2019, 185, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Salama, S.A.; Mohammad, M.A.; Diaz-Arrastia, C.R.; Kamel, M.W.; Kilic, G.S.; Ndofor, B.T.; Abdel-Baki, M.S.; Theiler, S.K. Estradiol-17β upregulates pyruvate kinase M2 expression to coactivate estrogen receptor-α and to integrate metabolic reprogramming with the mitogenic response in endometrial cells. J. Clin. Endocrinol. Metab. 2014, 99, 3790–3799. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.N.; Renaud, H.; Guo, Y. Bioinformatics-based identification of miR-542-5p as a predictive biomarker in breast cancer therapy. Hereditas 2018, 155, 17. [Google Scholar] [CrossRef]
- Frasor, J.; Chang, E.C.; Komm, B.; Lin, C.Y.; Vega, V.B.; Liu, E.T.; Miller, L.D.; Smeds, J.; Bergh, J.; Katzenellenbogen, B.S. Gene expression preferentially regulated by tamoxifen in breast cancer cells and correlations with clinical outcome. Cancer Res. 2006, 66, 7334–7340. [Google Scholar] [CrossRef] [Green Version]
- Lu, N.; Li, M.; Lei, H.; Jiang, X.; Tu, W.; Lu, Y.; Xia, D. Butyric acid regulates progesterone and estradiol secretion via cAMP signaling pathway in porcine granulosa cells. J. Steroid. Biochem. Mol. Biol. 2017, 172, 89–97. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | MS/MS | Peptide Number | Protein Number |
|---|---|---|---|
| PE | 4003 | 3405 | 231 |
| E | 2200 | 2179 | 219 |
| AE | 2813 | 2589 | 229 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Song, C.; Qi, K.; Liu, Y.; Dou, Y.; Li, X.; Qiao, R.; Wang, K.; Han, X.; Li, X. Identification of Estrus in Sows Based on Salivary Proteomics. Animals 2022, 12, 1656. https://doi.org/10.3390/ani12131656
Li C, Song C, Qi K, Liu Y, Dou Y, Li X, Qiao R, Wang K, Han X, Li X. Identification of Estrus in Sows Based on Salivary Proteomics. Animals. 2022; 12(13):1656. https://doi.org/10.3390/ani12131656
Chicago/Turabian StyleLi, Chenlei, Chenglei Song, Kunlong Qi, Yingke Liu, Yaqing Dou, Xiuling Li, Ruimin Qiao, Kejun Wang, Xuelei Han, and Xinjian Li. 2022. "Identification of Estrus in Sows Based on Salivary Proteomics" Animals 12, no. 13: 1656. https://doi.org/10.3390/ani12131656