
Diversity, Distribution, and Biogeography of Freshwater Fishes in Guangxi, China


 , ,
, ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
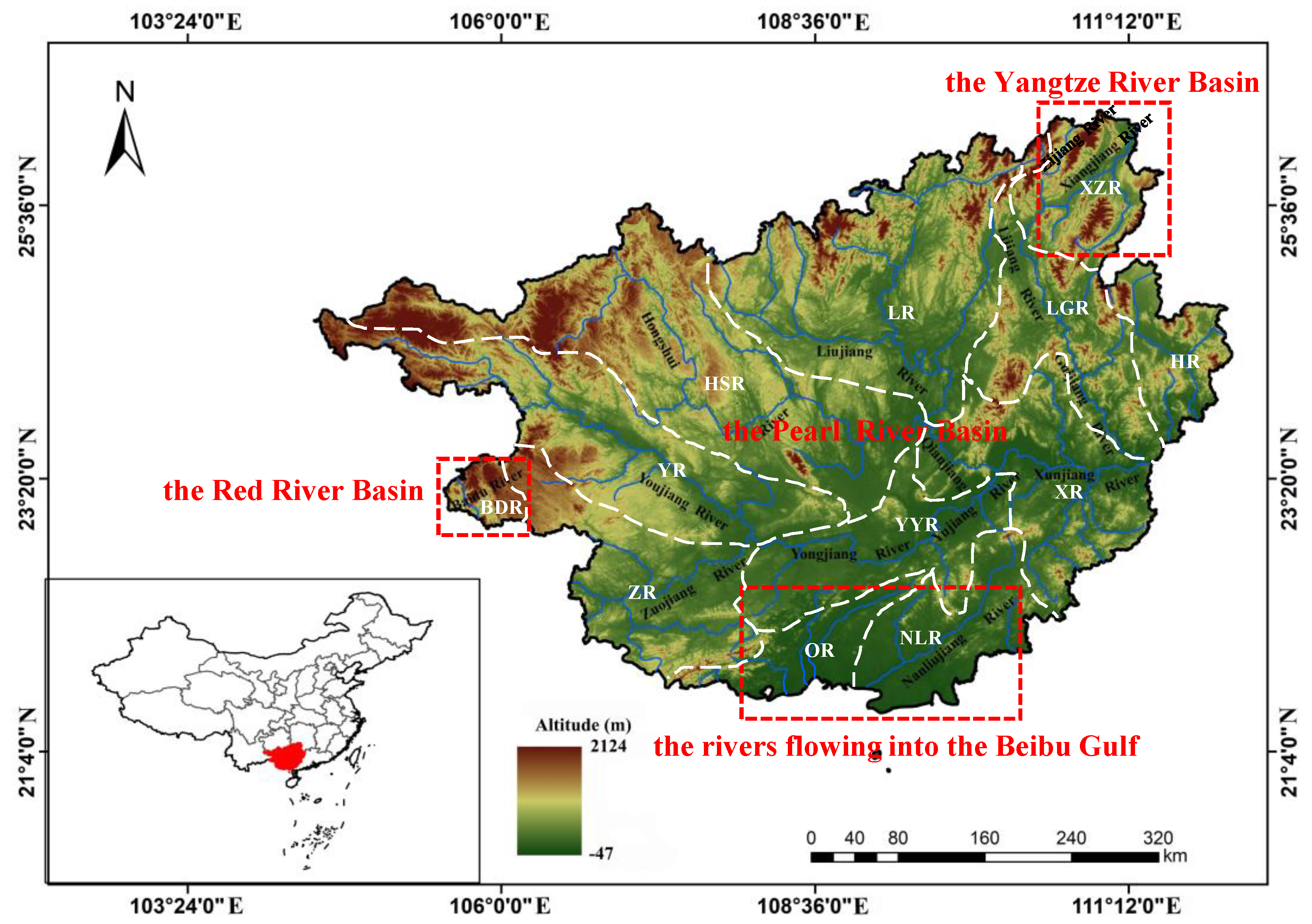
2.1. Study Area
2.2. Data Sources
2.3. Data Processing
3. Results
3.1. Faunal Composition
3.2. Fish Distribution
3.3. Biodiversity at Three Taxa Levels
3.4. Taxonomic Diversity Index
3.5. Beta Diversity Patterns
4. Discussion
4.1. Change of Freshwater Fish Species Composition
4.2. Spatial Patterns of Fish Diversity
4.3. Biogeographic Analysis of Fish
4.4. Fish Protection
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef]
- Collen, B.; Whitton, F.; Dyer, E.E.; Baillie, J.E.M.; Cumberlidge, N.; Darwall, W.R.T.; Pollock, C.; Richman, N.I.; Soulsby, A.M.; Bohm, M. Global patterns of freshwater species diversity, threat and endemism. Glob. Ecol. Biogeogr. 2014, 23, 40–51. [Google Scholar] [CrossRef] [Green Version]
- Su, G.; Logez, M.; Xu, J.; Tao, S.; Villéger, S.; Brosse, S. Human impacts on global freshwater fish biodiversity. Science 2021, 371, 835–838. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.W.; Nie, Y.G.; Miao, H.X.; Lu, H.; Hu, Y.B. Advancements of the researches on biodiversity loss mechanisms. Sci. Bull. 2014, 59, 430–437. [Google Scholar] [CrossRef]
- Higgins, J.; Zablocki, J.; Newsock, A.; Krolopp, A.; Tabas, P.; Salama, M. Durable freshwater protection: A framework for establishing and maintaining long-term protection for freshwater ecosystems and the values they sustain. Sustainability 2021, 13, 1950. [Google Scholar] [CrossRef]
- Chen, W.J.; Liu, L.; Wang, J.J.; Zhou, L. Threatened freshwater fish need protection. Science 2021, 374, 164. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Z.; Xue, Y.; Zhang, C.L.; Ren, Y.P.; Xu, B.D.; Chen, Y. Sampling intensity influences the estimation of functional diversity indices of fish communities. Ecol. Indic. 2021, 121, 107169. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. The taxonomic distinctness measure of biodiversity: Weighting of step lengths between hierarchical levels. Mar. Ecol. Prog. Ser. 1999, 184, 21–29. [Google Scholar] [CrossRef]
- Jiang, Z.G.; Ji, L.Q. Avian-mammalian species diversity in nine representative sites in China. Biodivers. Sci. 1999, 7, 220–225. [Google Scholar]
- Margules, C.R.; Pressey, R.L. Systematic conservation planning. Nature 2000, 405, 243–253. [Google Scholar] [CrossRef]
- Heino, J.; Tolonen, K.T. Ecological drivers of multiple facets of beta diversity in a lentic macroinvertebrate metacommunity. Limnol. Oceanogr. 2017, 62, 2431–2444. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2010, 19, 134–143. [Google Scholar] [CrossRef]
- Tuya, F.; Herrero, B.A.; Bosch, N. Reef fish at a remote tropical island (Principe island, gulf of guinea): Disentangling taxonomic, functional and phylogenetic diversity patterns with depth. Mar. Freshw. Res. 2018, 69, 395–402. [Google Scholar] [CrossRef]
- Shi, Z.; Zhang, J.; Wu, H.; Yang, J.; Hu, M. Taxonomic diversity pattern and composition of fish species in the upper reaches of Ganjiang River, Jiangxi, China. PLoS ONE 2020, 15, e0241762. [Google Scholar] [CrossRef]
- Fisheries Research Institute of Guangxi Zhuang Autonomous Region. Freshwater Fishes from Guangxi, 2nd ed.; Guangxi People’s Publishing House: Nanning, China, 2006. [Google Scholar]
- Zhang, P.L.; Huang, T.F.; Wu, T.; Huang, X.L.; Zhang, Y.X.; Liu, Z.X. Checklist, distribution and conservation of typical cave fish in China. Carsol. Sin. 2019, 38, 937–945. [Google Scholar]
- Lan, J.H. Cave Fishes of Guangxi; Science Press: Beijing, China, 2013. [Google Scholar]
- Li, G.F. Fishes of the Pearl River; Science Press: Beijing, China, 2018. [Google Scholar]
- Li, G.F. Investigation and Research on Main River Fish Resources in Guangxi of the Pearl River Basin; Science Press: Beijing, China, 2020. [Google Scholar]
- Shi, R.D.; Wu, Z.Q.; Huang, L.L.; Feng, W.L.; Zhu, Z.J.; Ding, Y. Fish Species Diversity of the Upper Xiangjiang River in North Guangxi Province. J. Guangxi Norm. Univ. Nat. Sci. Ed. 2015, 33, 127–136. [Google Scholar]
- Deng, M.X.; Huang, L.L.; Yin, C.; Mo, W.M.; Huang, J.; Wu, Z.Q. Ecological Health Assessment of Guijiang River Based on Fish Index of Biotic Integrity. Ind. Saf. Environ. 2018, 44, 87–90. [Google Scholar]
- Wang, X.; Liu, F.; Yu, D.; Liu, H.Z. Mitochondrial divergence suggests unexpected high species diversity in the Opsariichthine fishes (Teleostei: Cyprinidae) and the revalidation of Opsariichthys macrolepis. Ecol. Evol. 2019, 9, 2664–2677. [Google Scholar] [CrossRef] [Green Version]
- Shuai, F.M.; Li, X.H.; He, A.Y.; Liu, Q.F.; Zhang, Q.Y.; Wu, Z.; Zhu, S.L. Fish diversity and distribution pattern of the Pearl River system in Guangxi. Acta Hydrobiol. Sin. 2020, 44, 819–828. [Google Scholar]
- Chen, Y.X.; He, D.K.; Chen, Y.F.; Sui, X.Y. Three new species of cobitid fish genus Cobitis (Teleostei, Cobitidae) from the river pearl basin of China. Folia Zool. 2015, 64, 1–16. [Google Scholar] [CrossRef]
- Yuan, L.; Zhang, E.; Huang, Y. Revision of the labeonine genus Three new species of cobitid Sinocrossocheilus (teleostei: Cyprinidae) from south china. Zootaxa 2008, 1809, 36–48. [Google Scholar] [CrossRef]
- Du, L.N.; Yang, J.; Min, R.; Chen, X.Y.; Yang, J.X. A review of the cypriniform tribe yunnanilini prokofiev, 2010 from China, with an emphasis on five genera based on morphologies and complete mitochondrial genomes of some species. Zool. Res. 2021, 42, 310–334. [Google Scholar] [CrossRef]
- Eschmeyer, W.N.; Fricke, R. Catalog of Fishes: Genera, Species, References. 2021. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 14 November 2021).
- Jiang, Z.G.; Jiang, J.P.; Wang, Y.Z. Red List of China’s Vertebrates. Biodivers. Sci. 2016, 24, 500–551. [Google Scholar]
- Clarke, K.; Warwick, R. A taxonomic distinctness index and its statistical properties. J. Appl. Ecol. 1998, 35, 523–531. [Google Scholar] [CrossRef]
- Carvalho, J.C.; Cardoso, P.; Gomes, P. Determining the relative roles of species replacement and species richness differences in generating beta-diversity patterns. Glob. Ecol. Biogeogr. 2012, 21, 760–771. [Google Scholar] [CrossRef]
- Baselga, A. The relationship between species replacement, dissimilarity derived from nestedness, and nestedness. Glob. Ecol. Biogeogr. 2012, 21, 1223–1232. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version. 2016. Available online: https://github.com/vegandevs/vegan (accessed on 7 September 2016).
- Chen, X.Y. Checklist of Fishes of Yunnan. Zool. Res. 2013, 34, 281–343. [Google Scholar]
- Cai, G.F. Geological structure and development history of major river systems in the Pearl River Basin. Pearl River 1981, 2, 13–18. [Google Scholar]
- Wei, Y.L.; Li, C.Z.; Chen, W.H. Characteristics and formation and evolution analysis of the karst landscape of Guangxi. Guangxi Sci. 2018, 25, 465–504. [Google Scholar]
- Almela, V.D.; Palmer, S.C.F.; Andreou, D.; Gillingham, P.K.; Travis, J.M.J.; Britton, J.R. Predicting the influence of river network configuration, biological traits and habitat quality interactions on riverine fish invasions. Divers. Distrib. 2022, 28, 257–270. [Google Scholar] [CrossRef]
- Wang, J.; Chen, L.; Tang, W.J.; Heino, J.; Jiang, X.M. Effects of dam construction and fish invasion on the species, functional and phylogenetic diversity of fish assemblages in the Yellow River Basin. J. Environ. Manag. 2021, 293, 112863. [Google Scholar] [CrossRef]
- Wang, K.C.; Wu, Z.Q.; Wu, L.; Tan, H.Y.; Zhao, L.C.; Liu, B.W. Morphological Variation Analysis on Redbelly Tilapia (Coptodon zillii) of Hongshui River in Guangxi. Southwest China J. Agric. Sci. 2018, 31, 2208–2216. [Google Scholar]
- Gu, D.E.; Yu, F.D.; Xu, M.; Wei, H.; Mu, X.D. Temperature effects on the distribution of two invasive Tilapia species (Tilapia zillii and Oreochromis niloticus) in the rivers of south China. J. Freshw. Ecol. 2018, 33, 521–534. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.Y.; Huang, X.X.; Lu, Z.Z. Taxonomic Diversity Pattern of Freshwater Fish in the Pearl River Basin. Yunnan Geogr. Environ. Res. 2017, 29, 41–47. [Google Scholar]
- Wang, S.K. The distribution of freshwater fish and the relationship between species diversity and basin characteristics of main rivers in China. Biodivers. Sci. 1997, 5, 197–201. [Google Scholar]
- Guo, Q.; Kelt, D.A.; Sun, Z.; Liu, H.; Hu, L.; Ren, H. Global variation in elevational diversity patterns. Sci. Rep. 2013, 3, 3007. [Google Scholar] [CrossRef]
- Yu, X.D.; Luo, T.H.; Zhou, H.Z. Large-scale patterns in species diversity of fishes in the Yangtze River Basin. Biodivers. Sci. 2005, 13, 473–495. [Google Scholar] [CrossRef]
- Wang, Y.H.; Hu, C.Y.; Song, N.; Wang, X.Y.; Gao, T.X. Taxonomic diversity of common fish species in Zhoushan coastal waters. J. Dalian Ocean Univ. 2020, 36, 488–494. [Google Scholar]
- Li, J.; Li, X.H. A New Record of Fish Tanichthys albonubes (Cypriniformes: Cyprinidae) in Guangxi, China. Chin. J. Zool. 2011, 46, 136–140. [Google Scholar]
- Yao, M.; You, H.E.; Peng, Z.G. Lanlabeo duanensis, a new genus and species of Labeonin fish (Teleostei: Cyprinidae) from Southern China. Zootaxa 2018, 4471, 556–568. [Google Scholar] [CrossRef]
- Zheng, L.P.; You, H.; Yang, J.X.; Wu, L.B.; Peng, Z.G. A new genus and species of Labeonini (Teleostei: Cyprinidae) from the Pearl River in China. PLoS ONE 2018, 13, e0199973. [Google Scholar] [CrossRef]
- Zhang, E.; Zhou, W. Sinigarra napoense, a new genus and species of Labeonin fishes (Teleostei: Cyprinidae) from Guangxi Province, South China. Zootaxa 2012, 3586, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Yang, J.; Chen, X.Y. Stenorynchoacrum xijiangensis, a new genus and a new species of Labeoninae fish from Guangxi, China (Teleostei: Cyprinidae). Zootaxa 2014, 3793, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Q.; Sun, H.Y.; He, D.K.; Chen, Y.F. Freshwater fish diversity in the upper and middle reaches of the Lancang-Mekong River. Biodivers. Sci. 2019, 27, 1090–1100. [Google Scholar]
- Sholihah, A.; Delrieu-Trottin, E.; Condamine, F.L.; Wowor, D.; Hubert, N. Impact of pleistocene eustatic fluctuations on evolutionary dynamics in southeast Asian biodiversity hotspots. Syst. Biol. 2021, 70, 940–960. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.H.; Zhang, C.G. Fish fauna and zoogeographical analysis of Shiwanda Shan Mountains, Guangxi, China. Biodivers. Sci. 2001, 9, 336–344. [Google Scholar]
- Chen, Y.Y.; Cao, W.X.; Zheng, C.Y. Ichthyofauna of the Zhujiang River with a discussion on zoogeographical divisions for freshwater fishes. Acta Hydrobiol. Sin. 1986, 10, 228–236. [Google Scholar]
- Dai, Y.G. Fish fauna and zoogeographical division in the Duliu River, Guizhou, China. J. Fish. China 2011, 35, 871–879. [Google Scholar] [CrossRef]
- Liu, L.G.; Wang, W.B.; Yang, C.Y.; Luo, Y.S.; Yang, P.H. Survey on status of fish resources in main stream of Zijiang River of Dongting Lake water system. South China Fish. Sci. 2014, 10, 1–10. [Google Scholar]
- Wiersma, Y.F.; Urban, D.L. Beta diversity and nature reserve system design in the Yukon, Canada. Conserv. Biol. 2005, 19, 1262–1272. [Google Scholar] [CrossRef] [Green Version]
- Bush, A.; Harwood, T.; Hoskins, A.J.; Mokany, K.; Ferrier, S. Current uses of beta-diversity in biodiversity conservation: A response to Socolar. Trends Ecol. Evol. 2016, 31, 337–338. [Google Scholar] [CrossRef] [PubMed]
- Socolar, J.B.; Gilroy, J.J.; Kunin, W.E.; Edwards, D.P. How should beta-diversity inform biodiversity conservation? Trends Ecol. Evol. 2016, 31, 67–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Fang, Y.H.; Liang, Y.Q.; Huang, D.M. Fish assemblages and taxonomic diversity in main stream of Hongshui River. J. Fish. China 2019, 32, 48–53. [Google Scholar]
- Wang, S.; Xie, Y. Red List of Chinese Species Volume I: Red List; Higher Education Press: Beijing, China, 2004. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| River Basin | Sub−Basins Abbreviation | Length (km) | Area (km2) | Average Gradient (%) | Average Precipitation (mm) | Average Temperature (°C) | Annual Average Runoff (×108 m3) | Average Altitude (m) |
|---|---|---|---|---|---|---|---|---|
| Pearl River | XR | 291 | 0.0895 | 1418 | 21.6 | 1230 | 25 | |
| YYR | 426 | 20,593 | 0.0199 | 1320 | 21.6 | 410 | 65 | |
| HR | 352 | 11,536 | 0.0580 | 1555 | 21.7 | 112 | 90 | |
| LGR | 426 | 19,288 | 0.0247 | 1890 | 19.8 | 42 | 103 | |
| LR | 773 | 51,713 | 0.0168 | 1600 | 19.0 | 410 | 99.3 | |
| HSR | 659 | 52,600 | 0.0380 | 1100 | 18.0 | 696 | 242 | |
| YR | 707 | 38,612 | 0.0280 | 1187 | 17.1 | 172 | 120 | |
| ZR | 346 | 32,068 | 0.0366 | 1300 | 20.7 | 174 | 89 | |
| Into the Beibu Gulf | NLR | 287 | 9232 | 0.0377 | 1458 | 21.9 | 77 | 46 |
| OR | 692 | 17,322 | 0.0354 | 1386 | 22.0 | 62 | 58 | |
| Yangtze River | XZR | 221 | 3414 | 0.0360 | 1236 | 17.0 | 44.9 | 225 |
| Red River | BDR | 62 | 1758 | 0.0700 | 1465 | 19.2 | 12 | 358 |
| Year | XR | YYR | HR | LGR | LR | HSR | YR | ZR | NLR | OR | XZR | BDR | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Δ+ | 2021 | 41.7 | 42.0 | 45.0 | 43.3 | 43.1 | 41.0 | 40.6 | 42.8 | 40.6 | 45.9 | 48.3 | 44.1 |
| 2006 | 41.8 | 41.1 | 43.6 | 42.4 | 42.9 | 42.1 | 40.6 | 42.6 | 40.2 | 45.6 | 48.0 | 43.0 | |
| Λ+ | 2021 | 480.9 | 471.8 | 460.3 | 419.4 | 417.5 | 394.9 | 432.0 | 433.1 | 449.9 | 454.8 | 418.2 | 381.2 |
| 2006 | 484.2 | 470.1 | 331.1 | 423.5 | 439.3 | 424.4 | 448.6 | 440.5 | 441.2 | 448.0 | 416.2 | 409.1 |
| Sub−Basin | β | |||||
|---|---|---|---|---|---|---|
| Sørensen Index | Jaccard Index | |||||
| βsor | βsim | βsne | βjac | βjtu | βjne | |
| XR | 0.45 ± 0.14 | 0.29 ± 0.13 | 0.16 ± 0.14 | 0.61 ± 0.12 | 0.43 ± 0.16 | 0.18 ± 0.15 |
| YYR | 0.45 ± 0.14 | 0.29 ± 0.14 | 0.16 ± 0.14 | 0.61 ± 0.13 | 0.43 ± 0.16 | 0.18 ± 0.15 |
| HR | 0.55 ± 0.11 | 0.31 ± 0.14 | 0.24 ± 0.14 | 0.7 ± 0.08 | 0.46 ± 0.15 | 0.25 ± 0.16 |
| LGR | 0.47 ± 0.16 | 0.2 ± 0.09 | 0.28 ± 0.18 | 0.63 ± 0.14 | 0.32 ± 0.12 | 0.31 ± 0.18 |
| LR | 0.47 ± 0.17 | 0.2 ± 0.09 | 0.27 ± 0.17 | 0.62 ± 0.14 | 0.33 ± 0.12 | 0.3 ± 0.17 |
| HSR | 0.53 ± 0.14 | 0.29 ± 0.09 | 0.24 ± 0.16 | 0.68 ± 0.11 | 0.44 ± 0.11 | 0.24 ± 0.15 |
| YR | 0.46 ± 0.13 | 0.3 ± 0.1 | 0.16 ± 0.12 | 0.62 ± 0.11 | 0.46 ± 0.12 | 0.16 ± 0.11 |
| ZR | 0.42 ± 0.15 | 0.22 ± 0.06 | 0.2 ± 0.13 | 0.58 ± 0.13 | 0.36 ± 0.08 | 0.22 ± 0.11 |
| NLR | 0.51 ± 0.11 | 0.32 ± 0.11 | 0.2 ± 0.13 | 0.67 ± 0.09 | 0.47 ± 0.13 | 0.2 ± 0.14 |
| OR | 0.5 ± 0.11 | 0.35 ± 0.1 | 0.15 ± 0.13 | 0.66 ± 0.09 | 0.51 ± 0.11 | 0.15 ± 0.13 |
| XZR | 0.51 ± 0.13 | 0.37 ± 0.16 | 0.14 ± 0.12 | 0.67 ± 0.1 | 0.52 ± 0.2 | 0.15 ± 0.15 |
| BDR | 0.83 ± 0.03 | 0.4 ± 0.1 | 0.42 ± 0.12 | 0.91 ± 0.02 | 0.57 ± 0.1 | 0.34 ± 0.11 |
| Total | 0.79 ± 0.05 | 0.60 ± 0.06 | 0.19 ± 0.03 | 0.88 ± 0.05 | 0.75 ± 0.05 | 0.13 ± 0.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, J.; Wu, Z.; Huang, L.; Gao, M.; Liu, H.; Sun, Y.; Rad, S.; Du, L. Diversity, Distribution, and Biogeography of Freshwater Fishes in Guangxi, China. Animals 2022, 12, 1626. https://doi.org/10.3390/ani12131626
He J, Wu Z, Huang L, Gao M, Liu H, Sun Y, Rad S, Du L. Diversity, Distribution, and Biogeography of Freshwater Fishes in Guangxi, China. Animals. 2022; 12(13):1626. https://doi.org/10.3390/ani12131626
Chicago/Turabian StyleHe, Jiayang, Zhiqiang Wu, Liangliang Huang, Minhui Gao, Hao Liu, Yangyan Sun, Saeed Rad, and Lina Du. 2022. "Diversity, Distribution, and Biogeography of Freshwater Fishes in Guangxi, China" Animals 12, no. 13: 1626. https://doi.org/10.3390/ani12131626