
Influenza A in Wild Boars: Viral Circulation in the Emilia-Romagna Region (Northern Italy) between 2017 and 2022

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Sampling
2.2. SwIAVs Detection, Subtyping and Viral Isolation
2.3. Genetic Characterization of the Isolated Viruses
3. Results
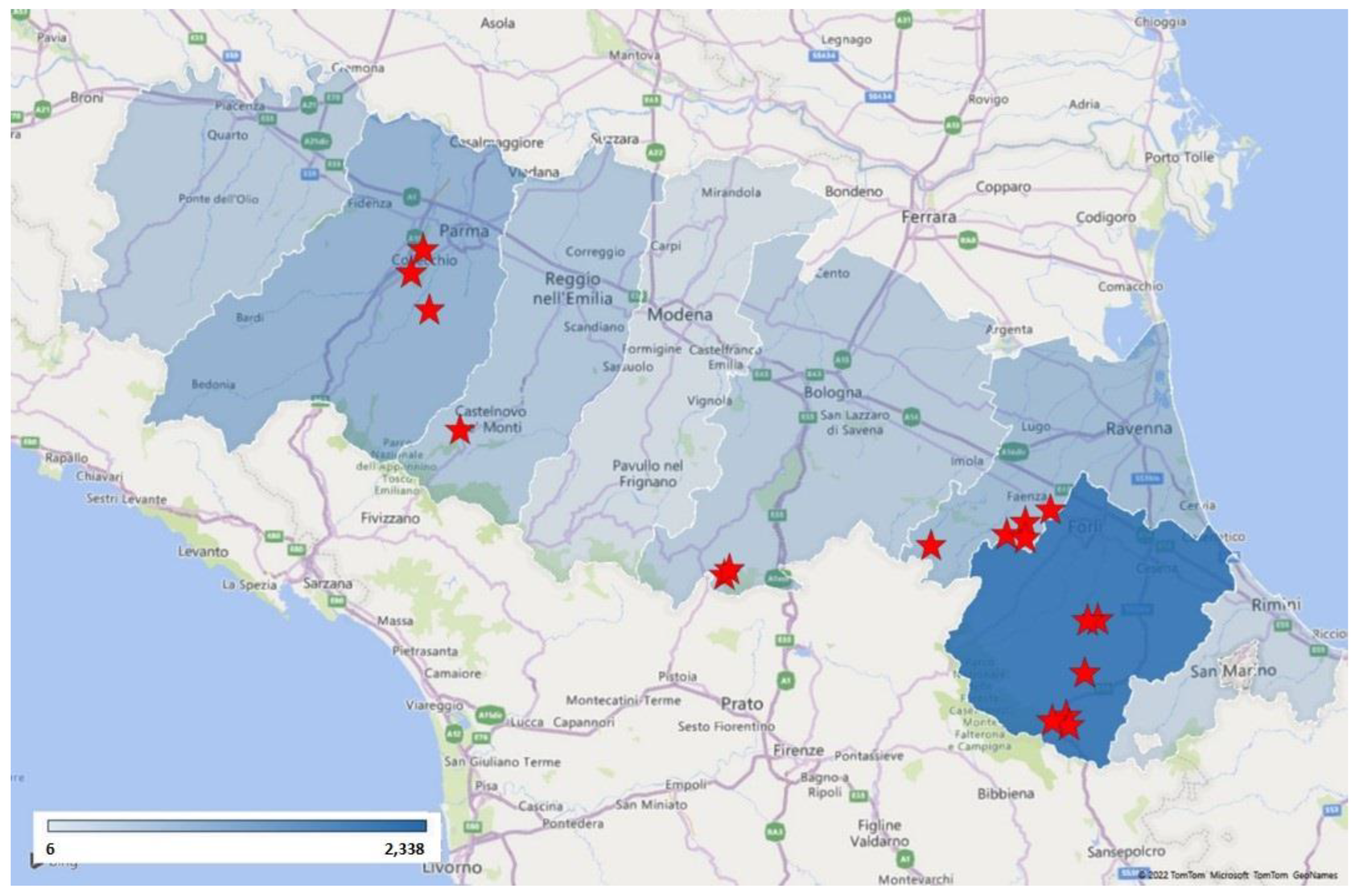
3.1. SwIAV Subtypes Circulation (Years 2017–2022)
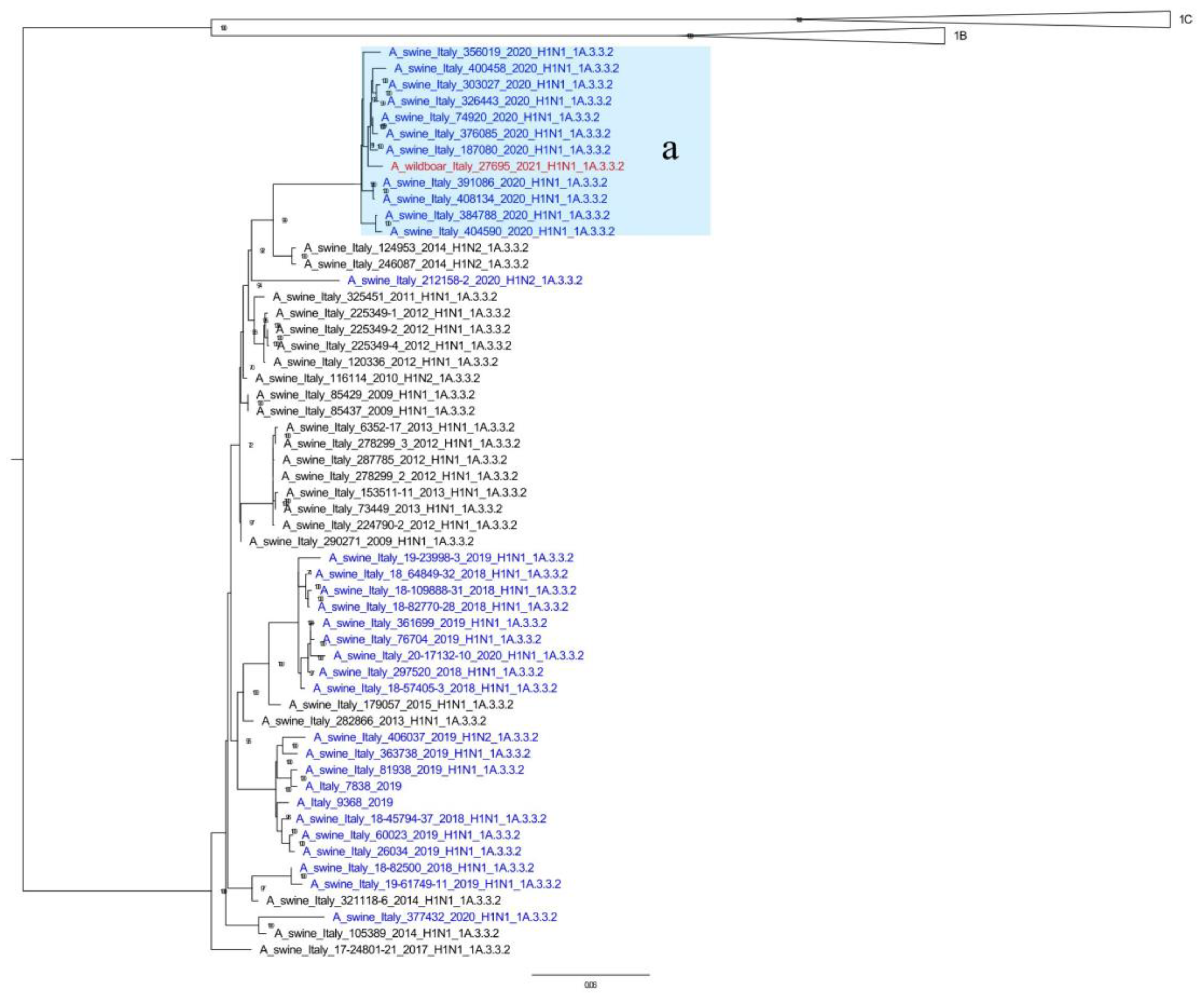
3.2. SwIAV Genotypes and Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kock, R. Drivers of disease emergence and spread: Is wildlife to blame? Onderstepoort J. Vet. Res. 2014, 81, 4. [Google Scholar] [CrossRef] [PubMed]
- Glud, H.A.; George, S.; Skovgaard, K.; Larsen, L.E. Zoonotic and reverse zoonotic transmission of viruses between humans and pigs. APMIS 2021, 129, 675–693. [Google Scholar] [CrossRef] [PubMed]
- Kaden, V.; Lange, E.; Starick, E.; Bruer, W.; Krakowski, W.; Klopries, M. Epidemiological survey of swine influenza A virus in selected wild boar populations in Germany. Vet. Microbiol. 2008, 131, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Kuntz-Simon, G.; Madec, F. Genetic and Antigenic Evolution of Swine Influenza Viruses in Europe and Evaluation of Their Zoonotic Potential. Zoonoses Public Health 2009, 56, 310–325. [Google Scholar] [CrossRef]
- Nelli, R.K.; Kuchipudi, S.V.; White, G.A.; Baquero Perez, B.; Dunham, S.P.; Chang, K.-C. Comparative distribution of human and avian type sialic acid influenza receptors in the pig. BMC Vet. Res. 2010, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Nelson, M.I.; Worobey, M. Origins of the 1918 Pandemic: Revisiting the Swine “Mixing Vessel” Hypothesis. Am. J. Epidemiol. 2018, 187, 2498–2502. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.-Y.; Tang, Y.-S.; Shaw, P.-C. Structure and Function of Influenza Virus Ribonucleoprotein. Subcell Biochem. 2018, 88, 95–128. [Google Scholar] [CrossRef]
- Dou, D.; Revol, R.; Östbye, H.; Wang, H.; Daniels, R. Influenza A Virus Cell Entry, Replication, Virion Assembly and Movement. Front. Immunol. 2018, 9, 1581. [Google Scholar] [CrossRef]
- ICTV. ICTV 9th Report (2011)—Orthomyxoviridae. Negative Sense RNA Viruses. 2011. Available online: https://talk.ictvonline.org/ictv-reports/ictv_9th_report/negative-sense-rna-viruses-2011/w/negrna_viruses/209/orthomyxoviridae (accessed on 26 April 2022).
- Mena, I.; Nelson, M.I.; Quezada-Monroy, F.; Dutta, J.; Cortes-Fernández, R.; Lara-Puente, J.H.; Castro-Peralta, F.; Cunha, L.F.; Trovão, N.S.; Lozano-Dubernard, B.; et al. Origins of the 2009 H1N1 influenza pandemic in swine in Mexico. eLife 2016, 5, e16777. [Google Scholar] [CrossRef]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of influenza A viruses. Microbiol. Rev. 1992, 56, 152–179. [Google Scholar] [CrossRef]
- Fouchier, R.A.M.; Munster, V.; Wallensten, A.; Bestebroer, T.M.; Herfst, S.; Smith, D.; Rimmelzwaan, G.F.; Olsen, B.; Osterhaus, A.D.M.E. Characterization of a Novel Influenza A Virus Hemagglutinin Subtype (H16) Obtained from Black-Headed Gulls. J. Virol. 2005, 79, 2814–2822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chastagner, A.; Hervé, S.; Quéguiner, S.; Hirchaud, E.; Lucas, P.; Gorin, S.; Béven, V.; Barbier, N.; Deblanc, C.; Blanchard, Y.; et al. Genetic and Antigenic Evolution of European Swine Influenza A Viruses of HA-1C (Avian-Like) and HA-1B (Human-Like) Lineages in France from 2000 to 2018. Viruses 2020, 12, 1304. [Google Scholar] [CrossRef]
- Anderson, T.K.; Macken, C.A.; Lewis, N.S.; Scheuermann, R.H.; Van Reeth, K.; Brown, I.H.; Swenson, S.L.; Simon, G.; Saito, T.; Berhane, Y.; et al. A Phylogeny-Based Global Nomenclature System and Automated Annotation Tool for H1 Hemagglutinin Genes from Swine Influenza A Viruses. mSphere 2016, 1, e00275-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henritzi, D.; Petric, P.P.; Lewis, N.S.; Graaf, A.; Pessia, A.; Starick, E.; Breithaupt, A.; Strebelow, G.; Luttermann, C.; Parker, L.M.K.; et al. Surveillance of European Domestic Pig Populations Identifies an Emerging Reservoir of Potentially Zoonotic Swine Influenza A Viruses. Cell Host Microbe 2020, 28, 614–627. [Google Scholar] [CrossRef] [PubMed]
- Zell, R.; Scholtissek, C.; Ludwig, S. Genetics, Evolution, and the Zoonotic Capacity of European Swine Influenza Viruses. Tuberculosis 2012, 370, 29–55. [Google Scholar] [CrossRef]
- OIE. OIE (2018): Influenza A virus of Swine (NB: Version Adopted in May 2015). Chapter 3.9.7 in Manual of Diagnostic Tests and Vaccines for Terrestrial Animals 2021. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.09.07_INF_A_SWINE.pdf (accessed on 26 April 2022).
- Ma, W. Swine influenza virus: Current status and challenge. Virus Res. 2020, 288, 198118. [Google Scholar] [CrossRef] [PubMed]
- Vittecoq, M.; Grandhomme, V.; Simon, G.; Herve, S.; Blanchon, T.; Renaud, F.; Thomas, F.; Gauthier-Clerc, M.; van der Werf, S. Study of influenza A virus in wild boars living in a major duck wintering site. Infect. Genet. Evol. 2011, 12, 483–486. [Google Scholar] [CrossRef]
- Foni, E.; Garbarino, C.; Chiapponi, C.; Baioni, L.; Zanni, I.; Cordioli, P. Epidemiological survey of swine influenza A virus in the wild boar population of two Italian provinces. Influ. Other Respir. Viruses 2013, 7, 16–20. [Google Scholar] [CrossRef]
- Delogu, M.; Cotti, C.; Vaccari, G.; Raffini, E.; Frasnelli, M.; Nicoloso, S.; Biacchessi, V.; Boni, A.; Foni, E.; Castrucci, M.R.; et al. Serologic and Virologic Evidence of Influenza a Viruses in Wild Boars (Sus scrofa) from Two Different Locations in Italy. J. Wildl. Dis. 2019, 55, 158–163. [Google Scholar] [CrossRef]
- Vicente, J.; León-Vizcaíno, L.; Gortázar, C.; Cubero, M.J.; González, M.; Martín-Atance, P. Antibodies to Selected Viral and Bacterial Pathogens in European Wild Boars from Southcentral Spain. J. Wildl. Dis. 2002, 38, 649–652. [Google Scholar] [CrossRef] [Green Version]
- Roic, B.; Jemersic, L.; Terzic, S.; Keros, T.; Balatinec, J.; Florijancic, T. Prevalence of antibodies to selected viral pathogens in wild boars (Sus Scrofa) in Croatia in 2005–06 AND 2009–10. J. Wildl. Dis. 2012, 48, 131–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grech-Angelini, S.; Hervé, S.; Rose, N.; Barbier, N.; Casabianca, F.; Maestrini, O.; Falchi, A.; Simon, G. Serological survey of influenza A viruses in domestic and wild Suidae in Corsica (France), a Mediterranean island environment. Prev. Vet. Med. 2018, 157, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Malmsten, A.; Magnusson, U.; Ruiz-Fons, F.; González-Barrio, D.; Dalin, A.-M. A Serologic Survey of Pathogens in Wild Boar (Sus Scrofa) in Sweden. J. Wildl. Dis. 2018, 54, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Schülein, A.; Ritzmann, M.; Christian, J.; Schneider, K.; Neubauer-Juric, A. Exposure of wild boar to Influenza A viruses in Bavaria: Analysis of seroprevalences and antibody subtype specificity before and after the panzootic of highly pathogenic avian influenza viruses A (H5N8). Zoonoses Public Health 2021, 68, 503–515. [Google Scholar] [CrossRef]
- Martin, B.E.; Sun, H.; Carrel, M.; Cunningham, F.L.; Baroch, J.A.; Hanson-Dorr, K.C.; Young, S.; Schmit, B.; Nolting, J.M.; Yoon, K.-J.; et al. Feral Swine in the United States Have Been Exposed to both Avian and Swine Influenza A Viruses. Appl. Environ. Microbiol. 2017, 83, e01346-17. [Google Scholar] [CrossRef] [Green Version]
- Carnevali, L.; Pedrotti, L.; Riga, F.; Toso, S. Banca Dati Ungulati: Status, distribuzione, consistenza, gestione e prelievo venatorio delle popolazioni di Ungulati in Italia. Rapporto 2001–2005 (2009). Biol. Cons. Fauna 2009, 117, 1–168. [Google Scholar]
- Chiapponi, C.; Prosperi, A.; Moreno, A.; Baioni, L.; Faccini, S.; Manfredi, R.; Zanni, I.; Gabbi, V.; Calanchi, I.; Fusaro, A.; et al. Genetic Variability among Swine Influenza Viruses in Italy: Data Analysis of the Period 2017–2020. Viruses 2021, 14, 47. [Google Scholar] [CrossRef]
- Slomka, M.J.; Densham, A.L.E.; Coward, V.J.; Essen, S.; Brookes, S.M.; Irvine, R.M.; Spackman, E.; Ridgeon, J.; Gardner, R.; Hanna, A.; et al. Original Article: Real time reverse transcription (RRT)-polymerase chain reaction (PCR) methods for detection of pandemic (H1N1) 2009 influenza virus and European swine influenza A virus infections in pigs. Influ. Other Respir. Viruses 2010, 4, 277–293. [Google Scholar] [CrossRef] [Green Version]
- Chiapponi, C.; Zanni, I.; Garbarino, C.; Barigazzi, G.; Foni, E. Comparison of the usefulness of the CACO-2 cell line with standard substrates for isolation of swine influenza A viruses. J. Virol. Methods 2010, 163, 162–165. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Harder, T.C.; Grosse Beilage, E.; Lange, E.; Meiners, C.; Döhring, S.; Pesch, S.; Noé, T.; Grund, C.; Beer, M.; Starick, E.; et al. Expanded Cocirculation of Stable Subtypes, Emerging Lineages, and New Sporadic Reassortants of Porcine Influenza Viruses in Swine Populations in Northwest Germany. J. Virol. 2013, 87, 10460–10476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, S.J.; Langat, P.; Reid, S.M.; Lam, T.T.; Cotten, M.; Kelly, M.; Van Reeth, K.; Qiu, Y.; Simon, G.; Bonin, E.; et al. Molecular Epidemiology and Evolution of Influenza Viruses Circulating within European Swine be-tween 2009 and 2013. J. Virol. 2015, 89, 9920–9931. [Google Scholar] [CrossRef] [Green Version]
- Chiapponi, C.; Ebranati, E.; Pariani, E.; Faccini, S.; Luppi, A.; Baioni, L.; Manfredi, R.; Carta, V.; Merenda, M.; Affanni, P.; et al. Genetic analysis of human and swine influenza A viruses isolated in Northern Italy during 2010–2015. Zoonoses Public Health 2017, 65, 114–123. [Google Scholar] [CrossRef] [Green Version]
- Scholtissek, C.; Bürger, H.; Kistner, O.; Shortridge, K. The nucleoprotein as a possible major factor in determining host specificity of influenza H3N2 viruses. Virology 1985, 147, 287–294. [Google Scholar] [CrossRef]
- Ma, W.; Lager, K.M.; Vincent, A.L.; Janke, B.H.; Gramer, M.R.; Richt, J.A. The Role of Swine in the Generation of Novel Influenza Viruses. Zoonoses Public Health 2009, 56, 326–337. [Google Scholar] [CrossRef]
- Vengust, G.; Valencak, Z.; Bidovec, A. A Serological Survey of Selected Pathogens in Wild Boar in Slovenia. J. Vet. Med. Ser. B 2006, 53, 24–27. [Google Scholar] [CrossRef]
- Kovalenko, G.; Molozhanova, A.; Halka, I.; Nychyk, S. Antibody Prevalence to Influenza Type A in Wild Boar of Northern Ukraine. Vector-Borne Zoonotic Dis. 2017, 17, 836–839. [Google Scholar] [CrossRef]
- OIE. African Swine Fever (ASF—Situation Report 7). Available online: https://www.oie.int/app/uploads/2022/03/asf-report7.pdf (accessed on 9 May 2022).
- Istituto Zooprofilattico Sperimentale dell’Umbria e delle Marche “Togo Rosati”. African Swine Fever Emergence Management. Available online: http://www.izsum.it/IZSUM/Common/pages02/wfDettListaDoppia.aspx?EDIT=False&ID=24719&IDMAP=48 (accessed on 9 May 2022).




{kind=link}
{kind=link}
{kind=link}
| Date | Laboratory Identification | Sampling Province | Subtype |
|---|---|---|---|
| 9 January 2018 | 2018/7179 | Reggio Emilia | Not typed |
| 29 December 2018 | 2019/3529 | Ravenna | H1avN1 (clade HA-1C) |
| 8 January 2019 | 2019/7109 | Ravenna | Not typed |
| 22 January 2019 | 2019/25042 | Ravenna | Not typed |
| 15 November 2019 | 2019/395623 | Ravenna | Not typed |
| 28 January 2020 | 2020/32567 | Ravenna | Not typed |
| 8 July 2020 | 2020/200093 | Parma | H1avNx (clade HA-1C) |
| 27 July 2020 | 2020/223686 | Parma | Not typed |
| 30 July 2020 | 2020/227865 | Parma | H1avN1 (clade HA-1C) |
| 30 November 2020 | 2020/390371 | Forlì-Cesena | Not typed |
| 1 December 2020 | 2020/392593-3 | Bologna | H1pdm09N1 (clade HA-1A) |
| 1 December 2020 | 2020/392593-4 | Bologna | H1N2 (clade HA-1B) |
| 10 December 2020 | 2020/403848 | Forlì-Cesena | Not typed |
| 22 December 2020 | 2020/421894 | Forlì-Cesena | Not typed |
| 4 January 2021 | 2021/214 | Forlì-Cesena | Not typed |
| 26 January 2021 | 2021/27695 | Forlì-Cesena | H1pdm09N1 (clade HA-1A) |
| 26 January 2021 | 2021/27709 | Forlì-Cesena | H1pdm09N1 (clade HA-1A) |
| Subtype | Nomenclature | HA | NA | PB2 | PB1 | PA | NP | M | NS | % | Wild Boars’ ID |
|---|---|---|---|---|---|---|---|---|---|---|---|
| H1N1 | A | 1C.2.1 | av | av | av | av | av | av | av | 58.3% | |
| 1C.2.1 | av | av | av | av | av | av | av | - | 2020/227865 | ||
| 1C.2 | av | av | av | av | av | av | av | 19.3% | |||
| 1C.2.2 | av | av | av | av | av | av | av | 4.3% | |||
| M | 1C.2.1 | av | av | av | av | av | pdm | av | 0.5% | ||
| 1C.2 | av | av | av | av | av | pdm | av | 1.6% | |||
| U | 1C.2.1 | av | pdm | pdm | pdm | pdm | pdm | pdm | 1.1% | ||
| 1C.2.1 | av | pdm | pdm | pdm | pdm | pdm | pdm | - | 2019/3529 | ||
| P | 1A.3.3.2 | pdm | pdm | pdm | pdm | pdm | pdm | pdm | 8.6% | ||
| 31 | 1A.3.3.2 | av | pdm | pdm | pdm | pdm | pdm | av | 4.3% | ||
| 1A.3.3.2 | av | pdm | pdm | pdm | pdm | pdm | av | - | 2021/27695 | ||
| 32 | 1A.3.3.2 | av | pdm | pdm | pdm | pdm | pdm | pdm | 1.6% | ||
| H | 1B.1.2.2 | av | av | av | av | av | av | av | 0.5% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prosperi, A.; Soliani, L.; Canelli, E.; Baioni, L.; Gabbi, V.; Torreggiani, C.; Manfredi, R.; Calanchi, I.; Pupillo, G.; Barsi, F.; et al. Influenza A in Wild Boars: Viral Circulation in the Emilia-Romagna Region (Northern Italy) between 2017 and 2022. Animals 2022, 12, 1593. https://doi.org/10.3390/ani12121593
Prosperi A, Soliani L, Canelli E, Baioni L, Gabbi V, Torreggiani C, Manfredi R, Calanchi I, Pupillo G, Barsi F, et al. Influenza A in Wild Boars: Viral Circulation in the Emilia-Romagna Region (Northern Italy) between 2017 and 2022. Animals. 2022; 12(12):1593. https://doi.org/10.3390/ani12121593
Chicago/Turabian StyleProsperi, Alice, Laura Soliani, Elena Canelli, Laura Baioni, Valentina Gabbi, Camilla Torreggiani, Roberta Manfredi, Irene Calanchi, Giovanni Pupillo, Filippo Barsi, and et al. 2022. "Influenza A in Wild Boars: Viral Circulation in the Emilia-Romagna Region (Northern Italy) between 2017 and 2022" Animals 12, no. 12: 1593. https://doi.org/10.3390/ani12121593