
Effects of Dietary Fucoidan Supplementation on Serum Biochemical Parameters, Small Intestinal Barrier Function, and Cecal Microbiota of Weaned Goat Kids

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Source of Fucoidan
2.2. Kids, Diet and Experimental Design
2.3. Sample Collection
2.4. Serum Biochemical Parameters
2.5. Intestinal Permeability
2.6. Antioxidant Capacity of Intestinal Mucosa
2.7. Intestinal Mucosal Immunity
2.8. RT-qPCR Analysis
2.9. DNA Extraction
2.10. 16S rRNA Gene Amplification and Illumina novaSeq Sequencing
2.11. Statistical Analysis
3. Results
3.1. Serum Biochemical Parameters
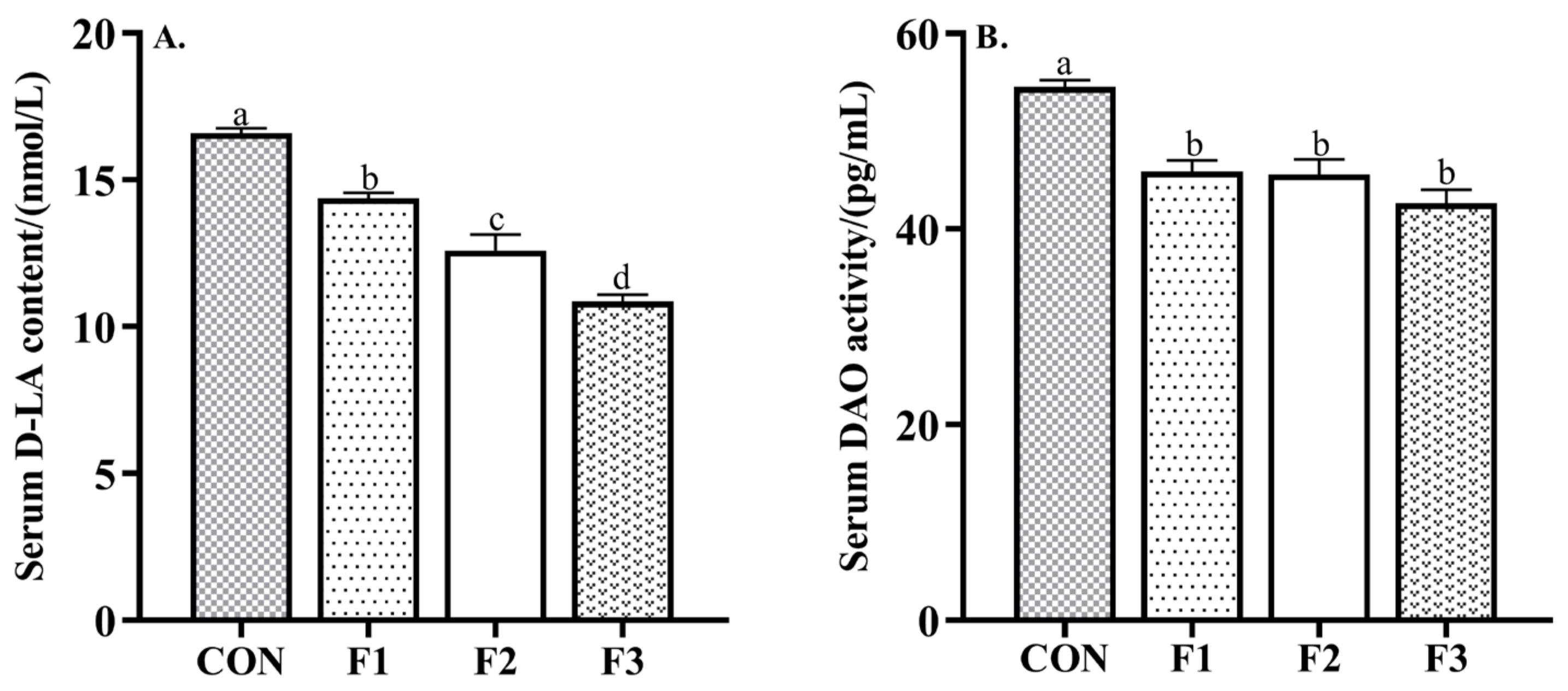
3.2. Intestinal Permeability
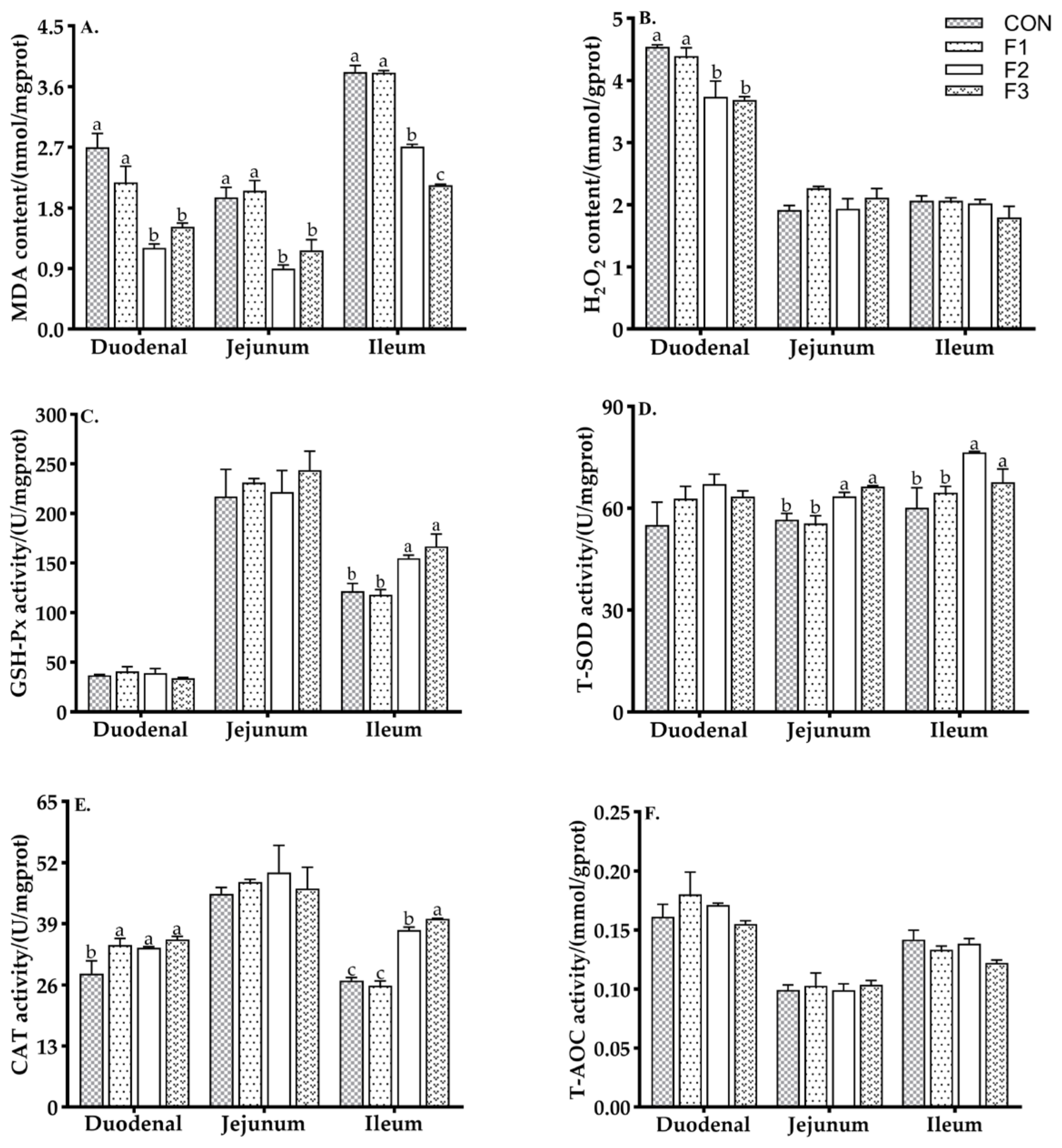
3.3. Antioxidant Capacity of Intestinal Mucosa
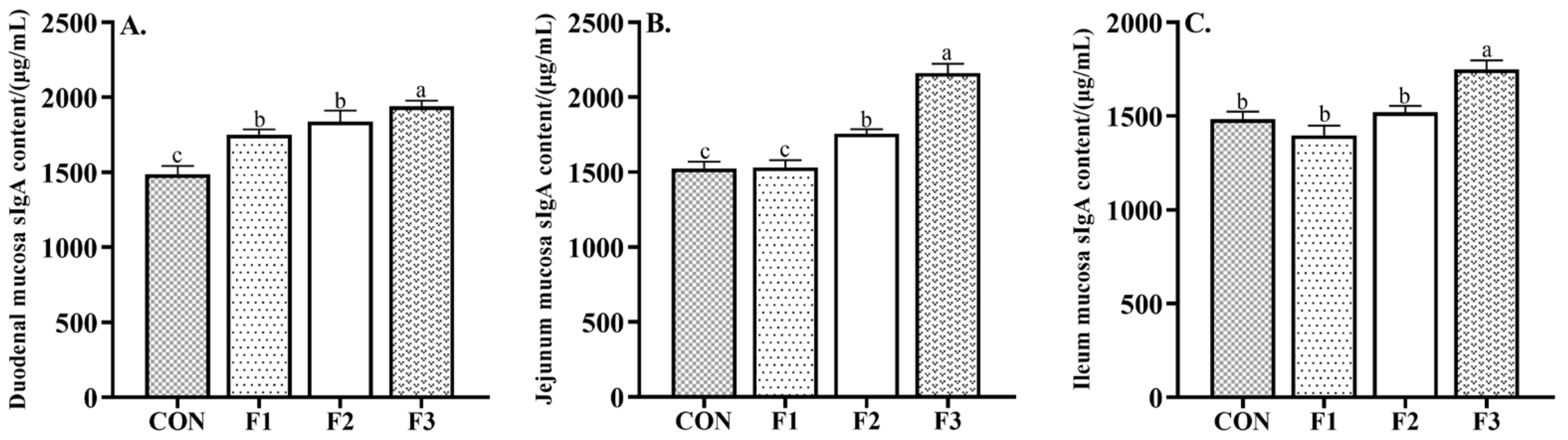
3.4. Intestinal Mucosal Immunity
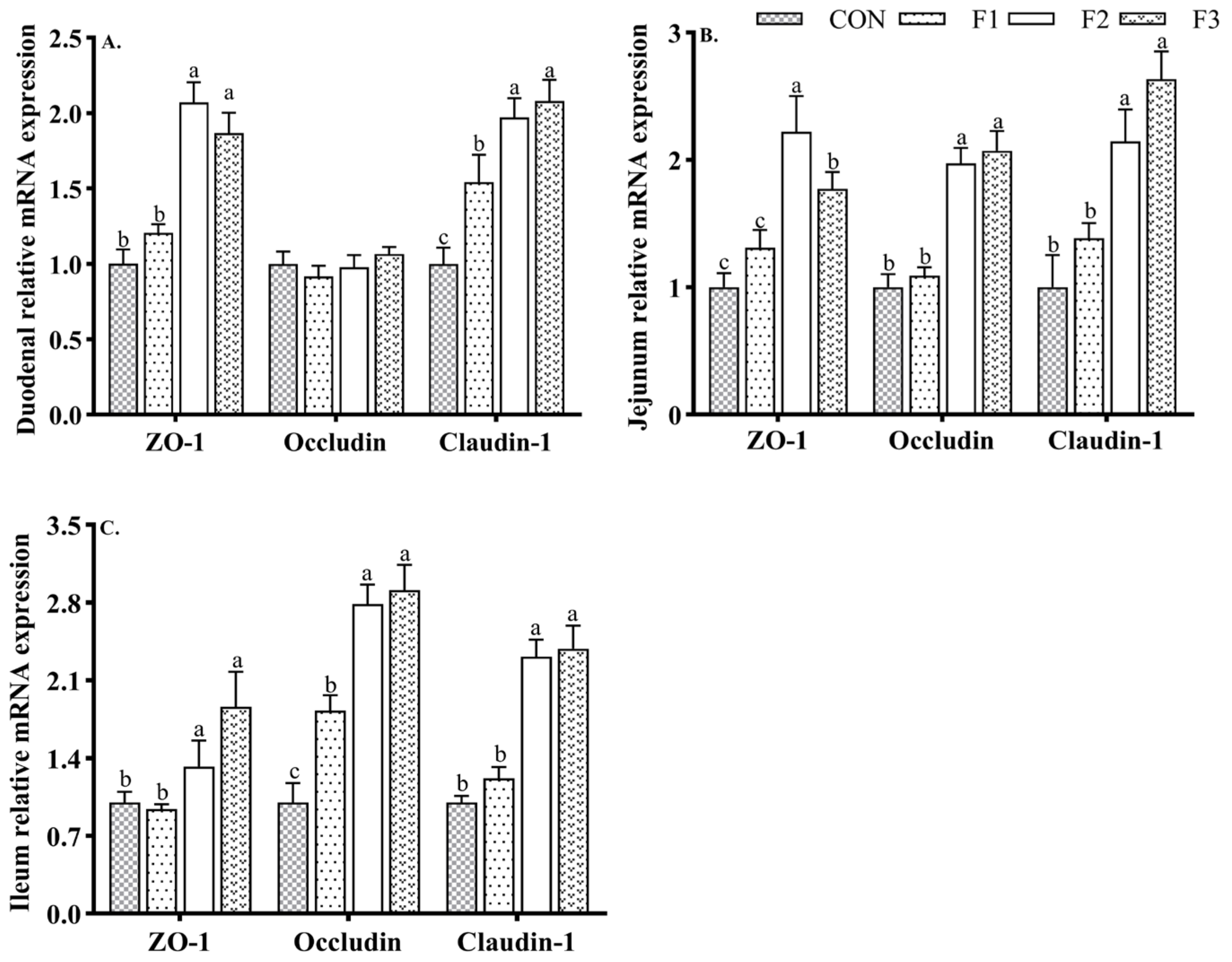
3.5. Intestinal Tight Junction Protein mRNA Expression
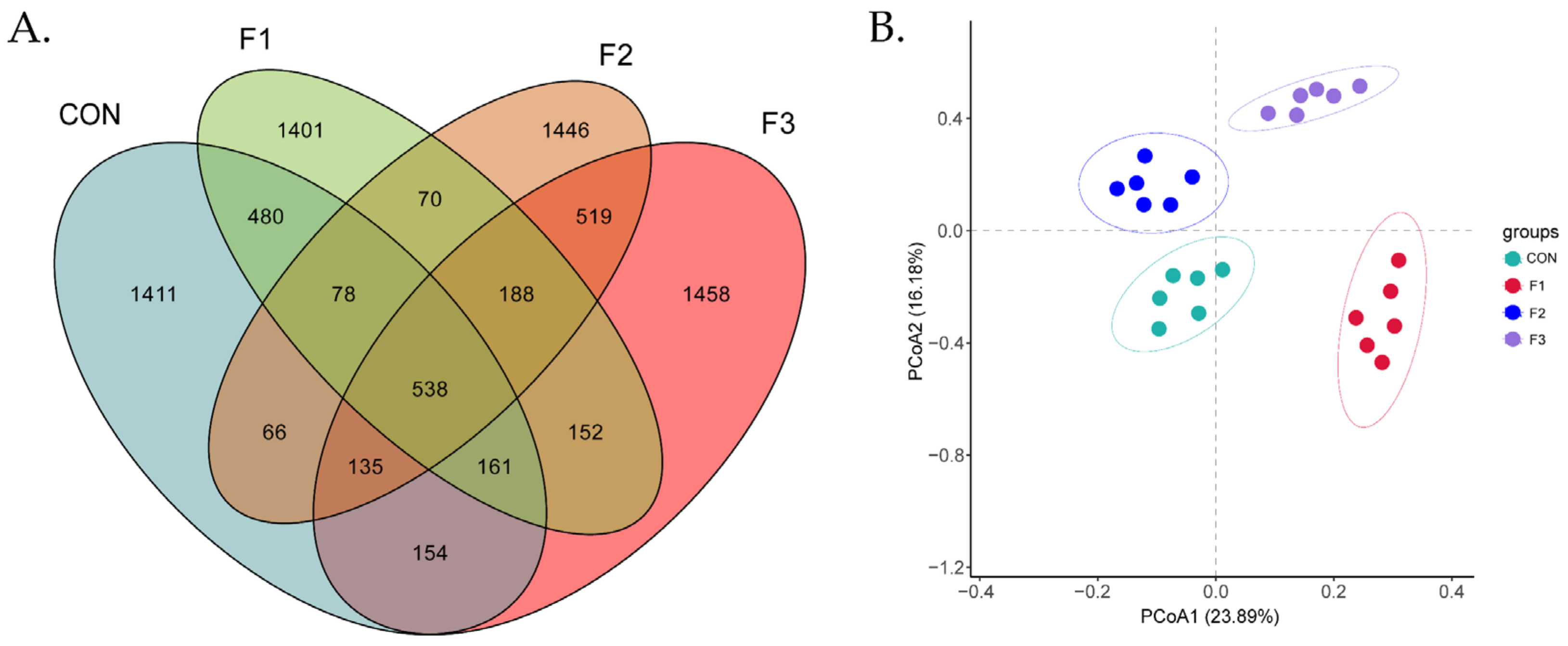
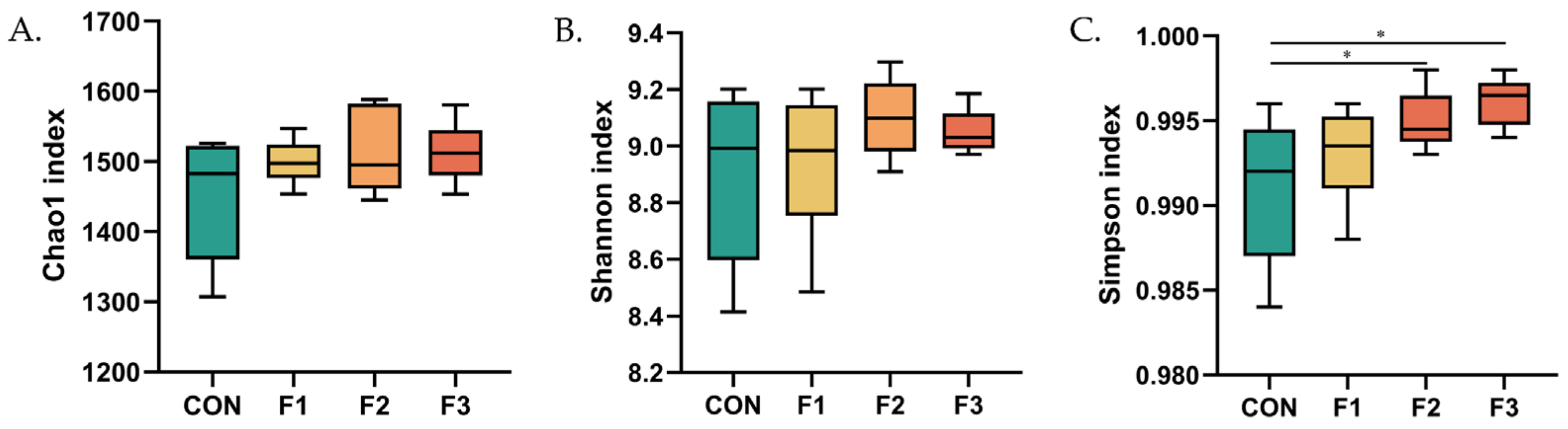
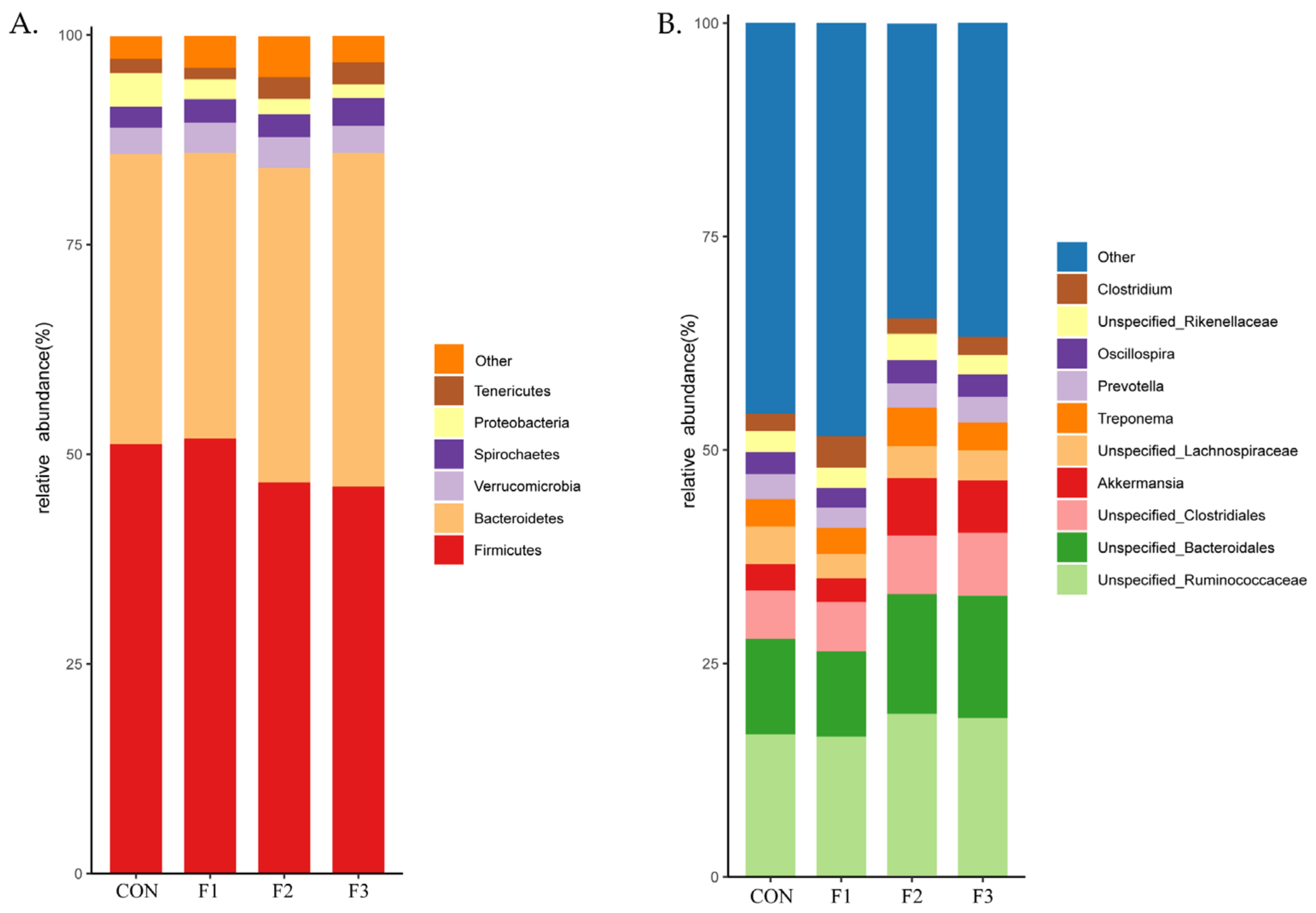
3.6. Alteration in the Composition of the Cecal Bacterial Community
3.7. Alteration of Cecal Microbiota Composition Caused from Fucoidan
4. Discussion
4.1. Serum Biochemical Indices
4.2. Small Intestinal Barrier Function
4.3. Cecal Microbial Community
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Han, Y.S.; Tang, C.H.; Li, Y.; Yu, Y.N.; Zhan, T.F.; Zhao, Q.Y.; Zhang, J.M. Effects of Dietary Supplementation with Clostridium butyricum on Growth Performance, Serum Immunity, Intestinal Morphology, and Microbiota as an Antibiotic Alternative in Weaned Piglets. Animals 2020, 10, 2287. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.Y.; Li, J.; Meng, Q.S.; Wu, D.L.; Xu, M. Effects of butyric acid supplementation of acidified milk on digestive function and weaning stress of cattle calves. Livest. Sci. 2019, 225, 78–84. [Google Scholar] [CrossRef]
- Chen, K.; Liu, Y.; Cheng, Y.; Yan, Q.; Zhou, C.; He, Z.; Zeng, J.; He, J.; Tan, Z. Supplementation of Lactobacillus plantarum or Macleaya cordata Extract Alleviates Oxidative Damage Induced by Weaning in the Lower Gut of Young Goats. Animals 2020, 10, 548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiao, J.Z.; Wu, J.; Wang, M.; Zhou, C.S.; Zhong, R.Z.; Tan, Z.L. Rhubarb Supplementation Promotes Intestinal Mucosal Innate Immune Homeostasis through Modulating Intestinal Epithelial Microbiota in Goat Kids. J. Agric. Food Chem. 2018, 66, 1047–1057. [Google Scholar] [CrossRef]
- Saleem, A.M.; Zanouny, A.I.; Singer, A.M. Growth performance, nutrients digestibility, and blood metabolites of lambs fed diets supplemented with probiotics during pre- and post-weaning period. Asian Australas. J. Anim. Sci. 2017, 30, 523–530. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.C.; Ou, B.H.; Liang, Z.L.; Zhang, R.; Zhao, Z.H. Algae-derived polysaccharides supplementation ameliorates heat stress-induced impairment of bursa of Fabricius via modulating NF-κB signaling pathway in broilers. Poult. Sci. 2021, 100, 101139. [Google Scholar] [CrossRef]
- Guo, Y.; Zhao, Z.H.; Pan, Z.Y.; An, L.L.L.; Balasubramanian, B.; Liu, W.C. New insights into the role of dietary marine-derived polysaccharides on productive performance, egg quality, antioxidant capacity, and jejunal morphology in late-phase laying hens. Poult. Sci. 2020, 99, 2100–2107. [Google Scholar] [CrossRef]
- Zhao, Y.; Balasubramanian, B.; Guo, Y.; Qiu, S.J.; Jha, R.; Liu, W.C. Dietary Enteromorpha polysaccharides supplementation improves breast muscle yield and is associated with modification of mRNA transcriptome in broiler chickens. Front. Vet. Sci. 2021, 8, 663988. [Google Scholar] [CrossRef]
- Liu, W.C.; Zhu, Y.R.; Zhao, Z.H.; Jiang, P.; Yin, F.Q. Effects of Dietary Supplementation of Algae-Derived Polysaccharides on Morphology, Tight Junctions, Antioxidant Capacity and Immune Response of Duodenum in Broilers under Heat Stress. Animals 2021, 11, 2279. [Google Scholar] [CrossRef]
- Zhu, Y.L.; Liu, L.B.; Sun, Z.Y.; Ji, Y.J.; Wang, D.Y.; Mei, L.; Shen, P.L.; Li, Z.X.; Tang, S.; Zhang, H.; et al. Fucoidan as a marine-origin prebiotic modulates the growth and antibacterial ability of Lactobacillus rhamnosus. Int. J. Biol. Macromol. 2021, 180, 599–607. [Google Scholar] [CrossRef]
- Liu, X.; Xi, X.Y.; Jia, A.R.; Zhang, M.S.; Cui, T.T.; Bai, X.F.; Shi, Y.P.; Liu, C.H. A fucoidan from Sargassum fusiforme with novel structure and its regulatory effects on intestinal microbiota in high-fat diet-fed mice. Food Chem. 2021, 358, 129908. [Google Scholar] [CrossRef]
- Zuo, T.; Li, X.M.; Chang, Y.G.; Duan, G.F.; Yu, L.; Zheng, R.; Xue, C.H.; Tang, Q.J. Dietary fucoidan of Acaudina molpadioides and its enzymatically degraded fragments could prevent intestinal mucositis induced by chemotherapy in mice. Food Funct. 2015, 6, 415–422. [Google Scholar] [CrossRef]
- Shi, H.J.; Chang, Y.G.; Yuan, G.; Wang, X.; Tang, Q.J. Dietary fucoidan of Acaudina molpadioides alters gut microbiota and mitigates intestinal mucosal injury induced by cyclophosphamide. Food Funct. 2017, 8, 3383–3393. [Google Scholar] [CrossRef]
- Rattigan, R.; Sweeney, T.; Vigors, S.; Thornton, K.; Rajauria, G.; O’Doherty, J.V. The Effect of Increasing Inclusion Levels of a Fucoidan-Rich Extract Derived from Ascophyllum nodosum on Growth Performance and Aspects of Intestinal Health of Pigs Post-Weaning. Mar. Drugs 2019, 17, 680. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Xue, M.L.; Yang, J.; Pei, Z.Q.; Zhang, N.; Qin, K.P.; Liang, H. Metabolic regulation mechanism of fucoidan via intestinal microecology in diseases. J. Sci. Food Agric. 2021, 101, 4456–4463. [Google Scholar] [CrossRef]
- Luo, J.M.; Wang, Z.; Fan, B.; Wang, L.; Liu, M.Y.; An, Z.Z.; Zhao, X. A comparative study of different fucoidan on the cefoperazone-induced gut microbiota disturbance and intestinal inammation. Food Funct. 2021, 12, 9087–9097. [Google Scholar] [CrossRef]
- Nagamine, T.; Nakazato, K.; Tomioka, S.; Iha, M.; Nakajima, K. Intestinal Absorption of Fucoidan Extracted from the Brown Seaweed, Cladosiphon okamuranus. Mar. Drugs 2014, 13, 48–64. [Google Scholar] [CrossRef]
- Walsh, A.M.; Sweeney, T.; O’Shea, C.J.; Doyle, D.N.; O’Doherty, J.V. Effect of dietary laminarin and fucoidan on selected microbiota, intestinal morphology and immune status of the newly weaned pig. Br. J. Nutr. 2013, 110, 1630–1638. [Google Scholar] [CrossRef] [Green Version]
- Lynch, M.B.; Sweeney, T.; Callan, J.J.; O’Sullivan, J.T.; O’Doherty, J.V. The effect of dietary Laminaria-derived laminarin and fucoidan on nutrient digestibility, nitrogen utilisation, intestinal microflora and volatile fatty acid concentration in pigs. J. Sci. Food Agric. 2010, 90, 430–437. [Google Scholar] [CrossRef]
- Wu, Q.F.; Wu, S.Y.; Cheng, Y.; Zhang, Z.S.; Mao, G.X.; Li, S.J.; Yang, Y.; Zhang, X.; Wu, M.J.; Tong, H.B. Sargassum fusiforme fucoidan modifies gut microbiota and intestinal metabolites during alleviation of hyperglycemia in type 2 diabetic mice. Food Funct. 2021, 12, 3572–3585. [Google Scholar] [CrossRef]
- Shokaiyan, M.; Ashayerizadeh, O.; Shargh, M.S.; Dastar, B. Algal Crude Fucoidan Alone or with Bacillus subtilis DSM 17299 in Broiler Chickens Diet: Growth Performance, Carcass Characteristics, Blood Metabolites, and Morphology of Intestine. Poult. Sci. J. 2019, 7, 87–94. [Google Scholar]
- Ikeda-Ohtsubo, W.; Nadal, A.L.; Zaccaria, E.; Iha, M.; Kitazawa, H.; Kleerebezem, H.; Brugman, S. Intestinal Microbiota and Immune Modulation in Zebrafish by Fucoidan from Okinawa Mozuku (Cladosiphon okamuranus). Front. Nutr. 2020, 7, 67. [Google Scholar] [CrossRef]
- Yang, W.G.; Chen, J.Y.; Guo, G.Z.; Wang, S.N.; Peng, S.; Gao, Z.H.; Zhao, Z.H.; Lan, R.X.; Yin, F.Q. The Effects of Fucoidan Dietary Supplementation on Growth Performance, Serum Antioxidant Capacity, Immune Function Indices and Intestinal Morphology in Weaned Kids. Animals 2022, 12, 574. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hu, C.; Ding, L.; Tang, Y.; Wei, H.; Jiang, C.; Yan, Q.; Dong, Q.; Degen, A.A. Astragalus membranaceus Alters Rumen Bacteria to Enhance Fiber Digestion, Improves Antioxidant Capacity and Immunity Indices of Small Intestinal Mucosa, and Enhances Liver Metabolites for Energy Synthesis in Tibetan Sheep. Animals 2021, 11, 3236. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Balasubramanian, B.; Zhao, Z.H.; Liu, W.C. Marine algal polysaccharides alleviate aflatoxin B1-induced bursa of Fabricius injury by regulating redox and apoptotic signaling pathway in broilers. Poult. Sci. 2020, 100, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.Y.; Ding, L.M.; Wang, X.J.; Yan, Q.; Jiang, C.X.; Hu, C.S.; Wang, G.W.; Zhou, Y.Q.; Henkin, Z.; Degen, A.A. Astragalus root extract improved average daily gain, immunity, antioxidant status and ruminal microbiota of early weaned yak calves. J. Sci. Food Agric. 2020, 101, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef]
- Elkatcha, M.; Soltan, M.; Essi, M. Effect of Pediococcus spp. Supplementation on Growth Performance, Nutrient Digestibility and Some Blood Serum Biochemical Changes of Fattening Lambs. Alex. J. Vet. Sci. 2016, 48, 124. [Google Scholar] [CrossRef]
- Abdolvahabi, S.; Zaeemi, M.; Mohri, M.; Naserian, A.A. Age related changes in serum biochemical profile of Saanen goat kids during the first three months of life. Rev. Méd. Vét. 2016, 167, 106–112. [Google Scholar]
- Klein, R.; Nagy, O.; Tóthová, C.; Chovanová, F. Clinical and Diagnostic Significance of Lactate Dehydrogenase and Its Isoenzymes in Animals. Vet. Med. Int. 2020, 2020, 5346483. [Google Scholar] [CrossRef]
- Piner Benli, P.; Kaya, M.; Dağlıoğlu, Y.K. Fucoidan Protects against Acute Sulfoxaflor-Induced Hematological/Biochemical Alterations and Oxidative Stress in Male Mice. Pharmaceuticals 2022, 15, 16. [Google Scholar] [CrossRef]
- Tavangar, P.; Gharahveysi, S.; Rezaeipour, V.; Irani, M. Efficacy of phytobiotic and toxin binder feed additives individually or in combination on the growth performance, blood biochemical parameters, intestinal morphology, and microbial population in broiler chickens exposed to aflatoxin B1. Trop. Anim. Health Prod. 2021, 53, 335. [Google Scholar] [CrossRef]
- Choi, J.; Raghavendran, H.R.B.; Sung, N.Y.; Kim, J.H.; Chun, B.S.; Ahn, D.H.; Choi, H.S.; Kang, K.W.; Lee, J.W. Effect of fucoidan on aspirin-induced stomach ulceration in rats. Chem. Biol. Interact. 2009, 183, 249–254. [Google Scholar] [CrossRef]
- Wang, Q.Y.; Zeng, X.L.; Zeng, Y.T.; Liu, X.R.W.; Wang, Y.C.; Wang, X.; Li, J.Z.; Wang, Y.Q.; Feng, Z.; Huang, P.F.; et al. Effects of Phragmites australis Shoot Remainder Silage on Growth Performance, Blood Biochemical Parameters, and Rumen Microbiota of Beef Cattle. Front. Vet. Sci. 2022, 9, 778654. [Google Scholar] [CrossRef]
- Musco, N.; Tudisco, R.; Esposito, G.; Iommelli, P.; Totakul, P.; D’Aniello, B.; Lombardi, P.; Amato, R.; Wanapat, M.; Infascelli, F. Effects of Linseed Supplementation on Milk Production, Composition, Odd- and Branched-Chain Fatty Acids, and on Serum Biochemistry in Cilentana Grazing Goats. Animals 2022, 12, 783. [Google Scholar] [CrossRef]
- AbdelWarith, A.W.A.; Younis, E.M.; AlAsgah, N.A.; Gewaily, M.S.; ElTonoby, S.M.; Dawood, M.A.O. Role of Fucoidan on the Growth Behavior and Blood Metabolites and Toxic Effects of Atrazine in Nile Tilapia Oreochromis niloticus (Linnaeus, 1758). Animals 2021, 11, 1448. [Google Scholar] [CrossRef]
- Zhang, J.Q.; Yu, C.Y.; Li, Z.H.; Li, J.; Chen, Y.P.; Wang, T.; Wang, C. Effects of Zinc Oxide Nanoparticles on Growth, Intestinal Barrier, Oxidative Status and Mineral Deposition in 21-Day-Old Broiler Chicks. Biol. Trace Elem. Res. 2021, 200, 1826–1834. [Google Scholar] [CrossRef]
- Izuddin, W.I.; Loh, T.C.; Foo, H.L.; Samsudin, A.A.; Human, A.M. Postbiotic L. plantarum RG14 improves ruminal epithelium growth, immune status and upregulates the intestinal barrier function in post-weaning lambs. Sci. Rep. 2019, 9, 9938. [Google Scholar] [CrossRef]
- Henrichs, B.S.; Brost, K.N.; Hayes, C.A.; Campbell, J.M.; Drackley, J.K. Effects of spray-dried bovine plasma protein in milk replacers fed at a high plane of nutrition on performance, intestinal permeability, and morbidity of Holstein calves. J. Dairy Sci. 2021, 104, 7856–7870. [Google Scholar] [CrossRef]
- Shi, L.G.; Xun, W.J.; Peng, W.Q.; Hu, H.C.; Cao, T.; Hou, G.Y. Effect of the Single and Combined Use of Curcumin and Piperine on Growth Performance, Intestinal Barrier Function, and Antioxidant Capacity of Weaned Wuzhishan Piglets. Front. Vet. Sci. 2020, 7, 418. [Google Scholar] [CrossRef]
- Xue, M.L.; Ji, X.Q.; Liang, H.; Liu, Y.; Wang, B.; Sun, L.L.; Li, W.W. The effect of fucoidan on intestinal flora and intestinal barrier function in rats with breast cancer. Food Funct. 2018, 9, 1214–1223. [Google Scholar] [CrossRef]
- Zhao, J.X.; Li, Q.; Zhang, R.X.; Liu, W.Z.; Ren, Y.S.; Zhang, C.X.; Zhang, J.X. Effect of dietary grape pomace on growth performance, meat quality and antioxidant activity in ram lambs. Anim. Feed Sci. Technol. 2018, 236, 76–85. [Google Scholar] [CrossRef]
- Zhao, J.X.; Li, K.; Su, R.; Liu, W.Z.; Ren, Y.S.; Zhang, C.X.; Du, M.; Zhang, J.X. Effect of dietary Tartary buckwheat extract supplementation on growth performance, meat quality and antioxidant activity in ewe lambs. Meat Sci. 2017, 134, 79–85. [Google Scholar] [CrossRef]
- Li, C.; Chen, J.; Zhao, M.; Liu, M.Q.; Yue, Z.K.; Liu, L.; Li, F.C. Effect of sodium butyrate on slaughter performance, serum indexes and intestinal barrier of rabbits. J. Anim. Physiol. Anim. Nutr. 2021, 106, 156–166. [Google Scholar] [CrossRef]
- Li, C.Y.; Chen, X.Y.; Zhang, B.; Liu, L.; Li, F.C. Sodium butyrate improved intestinal barrier in rabbits. Ital. J. Anim. Sci. 2020, 19, 1472–1482. [Google Scholar] [CrossRef]
- Chen, Y.X.; Xie, Y.N.; Zhong, R.Q.; Han, H.; Liu, L.; Chen, L.; Zhang, H.F.; Yves, B.; Nadia, E. Effects of graded levels of xylo-oligosaccharides on growth performance, serum parameters, intestinal morphology and intestinal barrier function in weaned piglets. J. Anim. Sci. 2021, 99, 183. [Google Scholar] [CrossRef]
- Shang, Z.H.; Wu, J.Y.; Wang, Y.X.; Zhang, Y.; Shang, Y.J. Effect of post-weaning stress diarrhea on intestinal barrier in lambs. Prog. Vet. Med. 2020, 41, 63–67. (In Chinese) [Google Scholar]
- Iraha, A.; Chinen, H.; Hokama, A.; Yonashiro, T.; Kinjo, T.; Kishimoto, K.; Nakamoto, M.; Hirata, T.; Kinjo, N.; Higa, F.; et al. Fucoidan enhances intestinal barrier function by upregulating the expression of claudin-1. World J. Gastroenterol. 2013, 19, 5500–5507. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, M.; Tang, Z.; Zhang, X.; Chen, J.; Sun, Z. Effects of Rhodotorula mucilaginosa fermentation product on the laying performance, egg quality, jejunal mucosal morphology and intestinal microbiota of hens. J. Appl. Microbiol. 2020, 128, 54–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgios, O.; Teixeira, A.G.V.; Foditsch, C.; Bicalho, M.L.; Machado, V.S.; Bicalho, R.C. Fecal microbial diversity in pre-weaned dairy calves as described by pyrosequencing of metagenomic 16S rDNA. Associations of Faecalibacterium species with health and growth. PLoS ONE 2017, 8, e63157. [Google Scholar]
- Brulc, J.M.; Antonopoulosi, D.A.; Berg Miller, M.E.; Wilson, M.K. Gene-centric metagenomics of the fiber-adherent bovine rumen microbiome reveals forage specific glycoside hydrolases. Proc. Natl. Acad. Sci. USA 2009, 106, 1948–1953. [Google Scholar] [CrossRef] [Green Version]
- Pichel, M.G.; Binsztein, N.; Qadri, F.; Giron, J.A. Type IV Longus Pilus of Enterotoxigenic Escherichia coli: Occurrence and Association with Toxin Types and Colonization Factors among Strains Isolated in Argentina. J. Clin. Microbiol. 2002, 40, 694–697. [Google Scholar] [CrossRef] [Green Version]
- Gu, S.; Chen, D.; Zhang, J.N.; Lv, X.; Wang, K.; Duan, L.P.; Nie, Y.; Wu, X.L. Bacterial Community Mapping of the Mouse Gastrointestinal Tract. PLoS ONE 2013, 8, e74957. [Google Scholar] [CrossRef] [Green Version]
- Suen, G.; Stevenson, D.M.; Bruce, D.C.; Chertkov, O.; Copeland, A.; Cheng, J.F.; Detter, C.; Detter, J.C.; Goodwin, L.A.; Han, C.S.; et al. Complete genome of the cellulolytic ruminal bacterium Ruminococcus albus 7. J. Bacteriol. 2011, 193, 5574–5575. [Google Scholar] [CrossRef] [Green Version]
- Mo, S.; Kim, B.S.; Yun, S.J.; Lee, J.J.; Yoon, S.H.; Oh, C.H. Genome sequencing of Clostridium butyricum DKU-01, isolated from infant feces. Gut Pathog. 2015, 7, 8. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Content |
|---|---|
| Ingredients (%) | |
| Pennisetum purpureum | 35.00 |
| Corn | 40.89 |
| Soybean meal | 13.98 |
| Wheat bran | 7.15 |
| Salt | 0.65 |
| CaHPO4 | 0.84 |
| Limestone | 0.84 |
| Premix 1 | 0.65 |
| Total | 100 |
| Nutrient level | |
| DM (%) | 88.86 |
| ME (MJ/kg) | 10.43 |
| CP (%) | 12.06 |
| NDF (%) | 30.06 |
| ADF (%) | 16.39 |
| Ca (%) | 0.76 |
| p (%) | 0.54 |
| Gene Symbols | Product Length, bp | Accession No. | Nucleotide Sequence of Primers (5′→3′) |
|---|---|---|---|
| ZO-1 | 111 | XM_018066118.1 | F: CGTCCTGATCCTGAACCTGTGTCT |
| R: GCTCTTCTCGCTCCTCCTGTGT | |||
| Occludin | 157 | XM_018065681.1 | F: CCTGTGTTGCCTCCACTCTT |
| R: TCCGTATAGCCTGTTCCATAGC | |||
| Claudin-1 | 95 | XM_005675123.3 | F: CCTGCTGGGACTAATAGCCA |
| R: CAGCCATTCGCATCTTCTGT | |||
| β-actin | 91 | NM_001314342.1 | F: TCCTTCCTGGGCATGGAATC |
| R: CGTAAAGGTCCTTGCGGATG |
| Item 1 | CON | F1 | F2 | F3 | SEM 2 | p-Value |
|---|---|---|---|---|---|---|
| 15 d | ||||||
| ALT (U/L) | 2.87 | 2.17 | 2.90 | 3.30 | 0.66 | 0.40 |
| AST (U/L) | 13.97 | 13.24 | 11.42 | 9.76 | 2.90 | 0.48 |
| LDH (U/L) | 4359.98 a | 3434.89 b | 3505.28 b | 3656.11 b | 371.55 | 0.03 |
| TG (mmol/L) | 1.00 | 0.93 | 0.80 | 0.97 | 0.12 | 0.37 |
| T-CHO (mmol/L) | 1.92 | 1.32 | 1.57 | 1.77 | 0.22 | 0.05 |
| GLU (mmol/L) | 0.73 a | 0.69 b | 0.60 c | 0.62 c | 0.04 | 0.01 |
| TP (mg/mL) | 27.69 b | 30.15 b | 30.40 b | 35.84 a | 2.00 | 0.01 |
| 30 d | ||||||
| ALT (U/L) | 2.74 | 4.19 | 3.38 | 3.05 | 0.75 | 0.26 |
| AST (U/L) | 18.78 a | 11.85 b | 10.34 b | 9.10 b | 2.55 | 0.02 |
| LDH (U/L) | 5174.24 a | 5851.01 a | 4848.49 b | 4093.43 b | 584.36 | 0.04 |
| TG (mmol/L) | 0.81 | 0.98 | 0.97 | 0.68 | 0.14 | 0.14 |
| T-CHO (mmol/L) | 1.49 | 1.38 | 1.64 | 1.76 | 0.15 | 0.09 |
| GLU (mmol/L) | 0.67 | 0.64 | 0.64 | 0.57 | 0.38 | 0.10 |
| TP (mg/mL) | 88.49 c | 107.08 b | 110.92 a | 119.06 a | 5.11 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, W.; Guo, G.; Chen, J.; Wang, S.; Gao, Z.; Zhao, Z.; Yin, F. Effects of Dietary Fucoidan Supplementation on Serum Biochemical Parameters, Small Intestinal Barrier Function, and Cecal Microbiota of Weaned Goat Kids. Animals 2022, 12, 1591. https://doi.org/10.3390/ani12121591
Yang W, Guo G, Chen J, Wang S, Gao Z, Zhao Z, Yin F. Effects of Dietary Fucoidan Supplementation on Serum Biochemical Parameters, Small Intestinal Barrier Function, and Cecal Microbiota of Weaned Goat Kids. Animals. 2022; 12(12):1591. https://doi.org/10.3390/ani12121591
Chicago/Turabian StyleYang, Weiguang, Guangzhen Guo, Jiayi Chen, Shengnan Wang, Zhenhua Gao, Zhihui Zhao, and Fuquan Yin. 2022. "Effects of Dietary Fucoidan Supplementation on Serum Biochemical Parameters, Small Intestinal Barrier Function, and Cecal Microbiota of Weaned Goat Kids" Animals 12, no. 12: 1591. https://doi.org/10.3390/ani12121591