
Cross-Sectional Survey of Antibiotic Resistance in Extended Spectrum β-Lactamase-Producing Enterobacteriaceae Isolated from Pigs in Greece

 , , , , and
, , , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Study Design
2.3. Sample Collection
2.4. Isolation and Identification of Extended Spectrum Cephalosporin Resistant (ESCR) Strains
2.5. Isolation and Identification of Salmonella spp.
2.6. Antimicrobial Susceptibility Testing
2.7. Phenotypic Confirmation of ESBL Production
2.8. Antibacterial Resistance Genes of ESBL-Producing Enterobacteriaceae
2.9. Statistical Analysis
3. Results
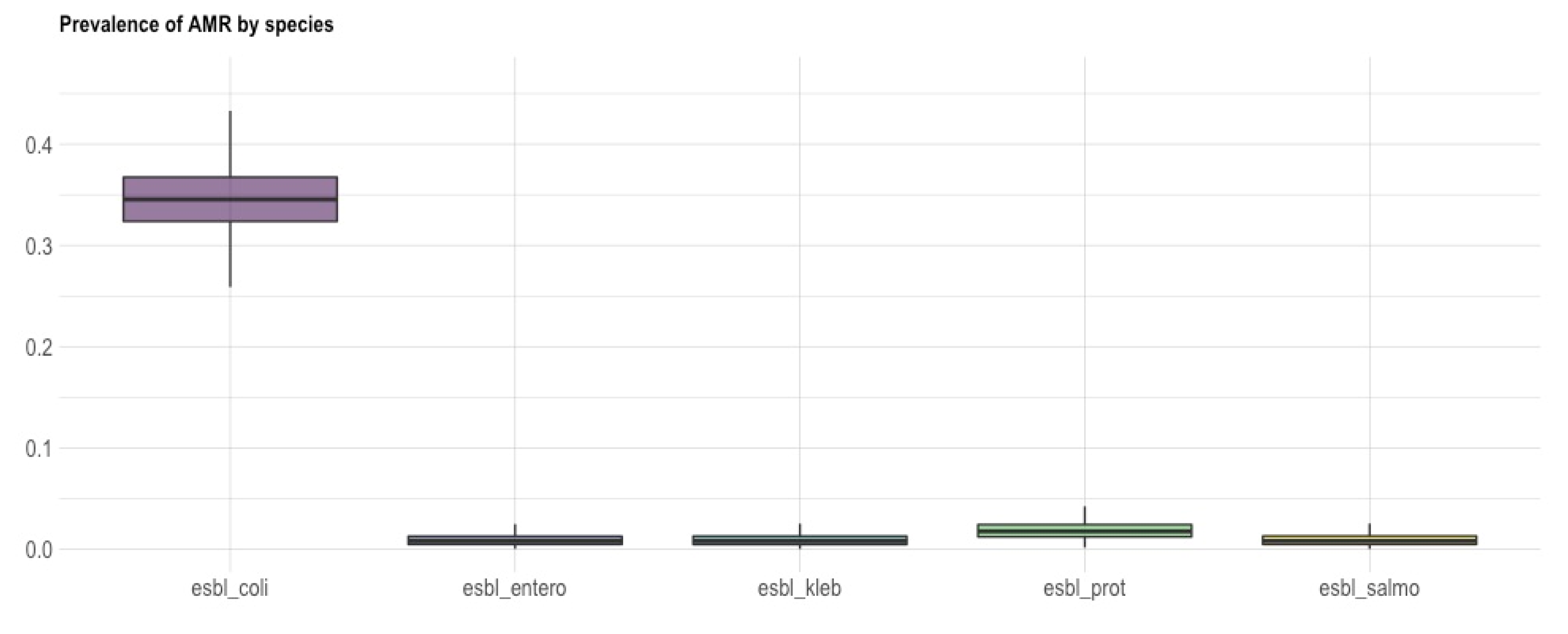
3.1. Isolation and Identification of ESBL-Producing Enterobacteriaceae
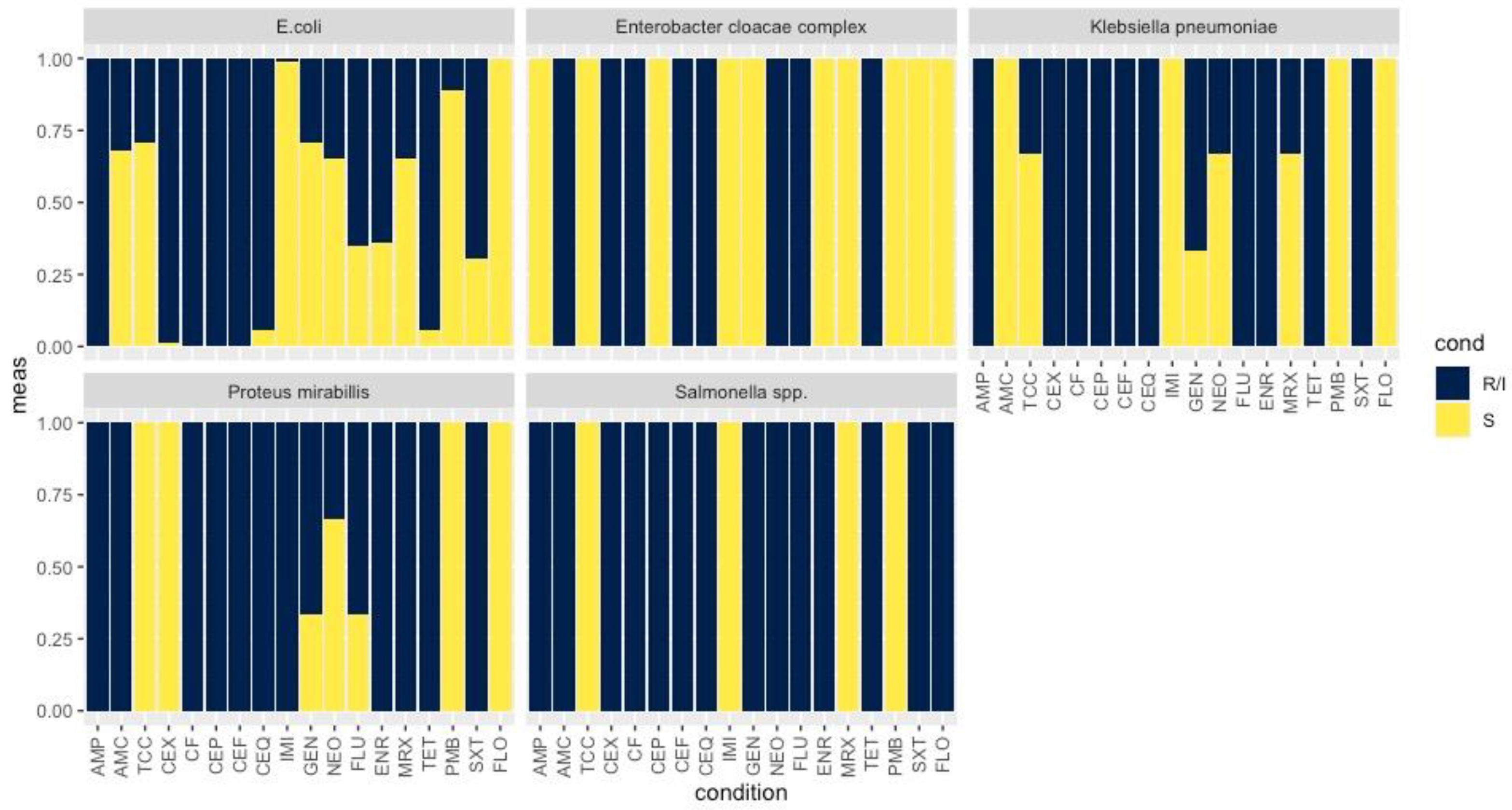
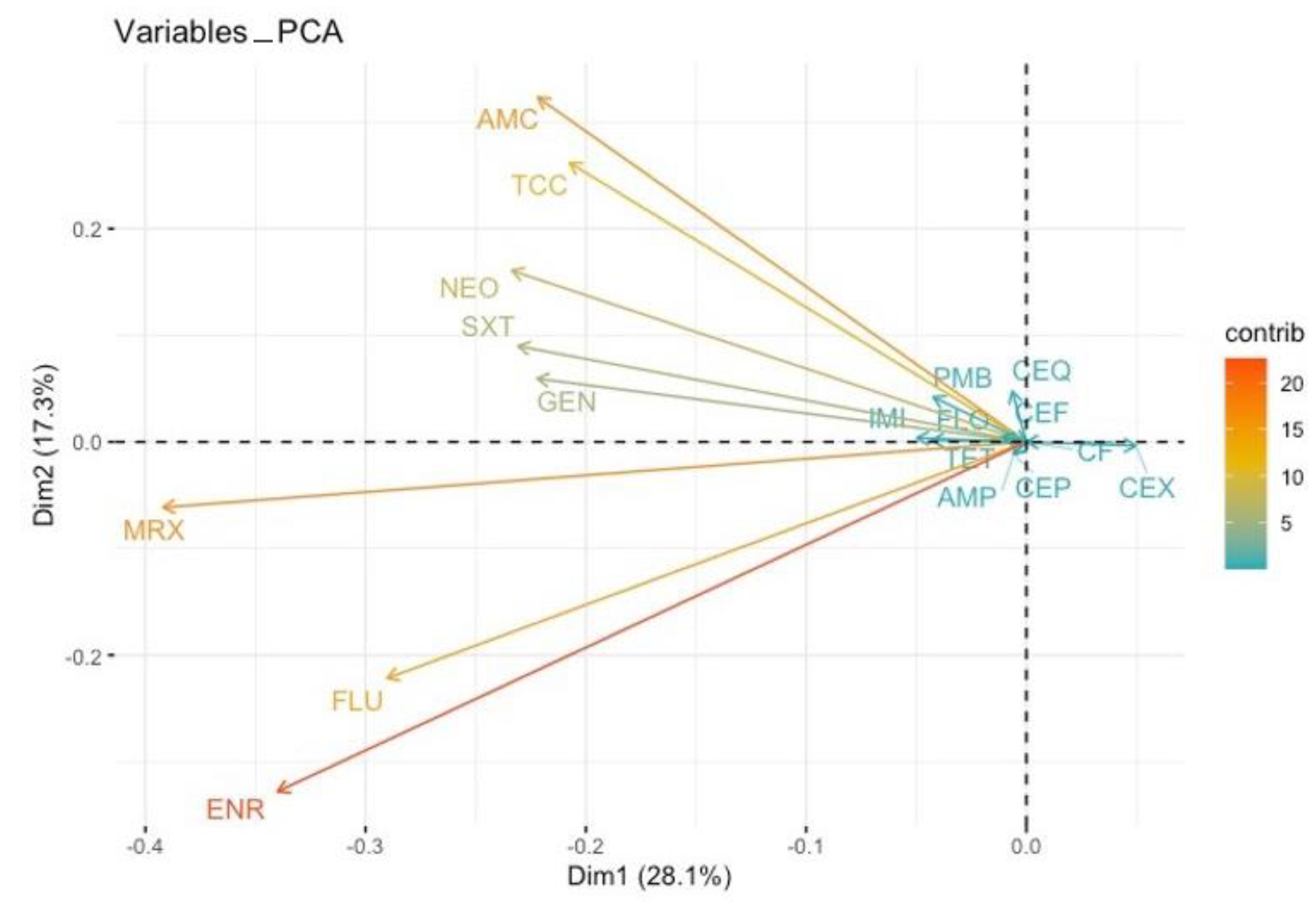
3.2. Antimicrobial Resistance Phenotype and Genotype of the ESBL-Producing Enterobacteriaceae
3.3. Antimicrobial Resistance Phenotype and Genotype of ESBL-Producing Enterobacteriaceae to non β-lactam Antibiotics
3.4. Logistic Regression Analysis Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lekagul, A.; Tangcharoensathien, V.; Yeung, S. Patterns of Antibiotic Use in Global Pig Production: A Systematic Review. Vet. Anim. Sci. 2019, 7, 100058. [Google Scholar] [CrossRef]
- Aarestrup, F.M. Veterinary Drug Usage and Antimicrobial Resistance in Bacteria of Animal Origin. Basic Clin. Pharmacol. Toxicol. 2005, 96, 271–281. [Google Scholar] [CrossRef]
- Landers, T.F.; Cohen, B.; Wittum, T.E.; Larson, E.L. A Review of Antibiotic Use in Food Animals: Perspective, Policy, and Potential. Public Health Rep. 2012, 127, 4–22. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. WHO Guidelines on Use of Medically Important Antimicrobials in Food-Producing Animals; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Burch, D.G.S.; Sperling, D. Amoxicillin—Current Use in Swine Medicine. J. Vet. Pharmacol. Ther. 2018, 41, 356–368. [Google Scholar] [CrossRef] [Green Version]
- Papatsiros, V.G.; Alexopoulos, C.; Kyriakis, S.C. Latest Information in Relation to Postpartum Dysgalactia Syndrome of Sows. J. Hell. Vet. Med. Soc. 2007, 58, 61–75. [Google Scholar] [CrossRef] [Green Version]
- ECDC/EFSA/EMA Second Joint Report on the Integrated Analysis of the Consumption of Antimicrobial Agents and Occurrence of Antimicrobial Resistance in Bacteria from Humans and Food-Producing Animals|EFSA. Available online: https://www.efsa.europa.eu/en/efsajournal/pub/4872 (accessed on 4 May 2022).
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Humans, Animals and Food in 2018/2019. EFSA J. 2021, 19, e06490. [Google Scholar] [CrossRef]
- Bengtsson, B.; Greko, C. Antibiotic Resistance—Consequences for Animal Health, Welfare, and Food Production. Ups. J. Med. Sci. 2014, 119, 96–102. [Google Scholar] [CrossRef]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global Trends in Antimicrobial Use in Food Animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [Green Version]
- Nhung, N.T.; Cuong, N.V.; Thwaites, G.; Carrique-Mas, J. Antimicrobial Usage and Antimicrobial Resistance in Animal Production in Southeast Asia: A Review. Antibiotics 2016, 5, 37. [Google Scholar] [CrossRef] [Green Version]
- Pollock, J.; Muwonge, A.; Hutchings, M.R.; Mainda, G.; Bronsvoort, B.M.; Gally, D.L.; Corbishley, A. Resistance to Change: AMR Gene Dynamics on a Commercial Pig Farm with High Antimicrobial Usage. Sci. Rep. 2020, 10, 1708. [Google Scholar] [CrossRef] [Green Version]
- Pholwat, S.; Pongpan, T.; Chinli, R.; Rogawski McQuade, E.T.; Thaipisuttikul, I.; Ratanakorn, P.; Liu, J.; Taniuchi, M.; Houpt, E.R.; Foongladda, S. Antimicrobial Resistance in Swine Fecal Specimens Across Different Farm Management Systems. Front. Microbiol. 2020, 11, 1238. [Google Scholar] [CrossRef]
- Bertelloni, F.; Cilia, G.; Bogi, S.; Ebani, V.V.; Turini, L.; Nuvoloni, R.; Cerri, D.; Fratini, F.; Turchi, B. Pathotypes and Antimicrobial Susceptibility of Escherichia Coli Isolated from Wild Boar (Sus Scrofa) in Tuscany. Animals 2020, 10, 744. [Google Scholar] [CrossRef]
- Cilia, G.; Turchi, B.; Fratini, F.; Bilei, S.; Bossù, T.; De Marchis, M.L.; Cerri, D.; Pacini, M.I.; Bertelloni, F. Prevalence, Virulence and Antimicrobial Susceptibility of Salmonella Spp., Yersinia Enterocolitica and Listeria Monocytogenes in European Wild Boar (Sus Scrofa) Hunted in Tuscany (Central Italy). Pathogens 2021, 10, 93. [Google Scholar] [CrossRef]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4, 2–4. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.-C.; Yeh, K.-S. Characteristics of Extended-Spectrum β-Lactamase–Producing Escherichia Coli Isolated from Fecal Samples of Piglets with Diarrhea in Central and Southern Taiwan in 2015. BMC Vet. Res. 2017, 13, 66. [Google Scholar] [CrossRef] [Green Version]
- Livermore, D.M.; Woodford, N. The β-Lactamase Threat in Enterobacteriaceae, Pseudomonas and Acinetobacter. Trends Microbiol. 2006, 14, 413–420. [Google Scholar] [CrossRef]
- Peirano, G.; Pitout, J.D.D. Extended-Spectrum β-Lactamase-Producing Enterobacteriaceae: Update on Molecular Epidemiology and Treatment Options. Drugs 2019, 79, 1529–1541. [Google Scholar] [CrossRef]
- Bergšpica, I.; Kaprou, G.; Alexa, E.A.; Prieto, M.; Alvarez-Ordóñez, A. Extended Spectrum β-Lactamase (ESBL) Producing Escherichia Coli in Pigs and Pork Meat in the European Union. Antibiotics 2020, 9, 678. [Google Scholar] [CrossRef]
- Pitout, J.D.; Laupland, K.B. Extended-Spectrum β-Lactamase-Producing Enterobacteriaceae: An Emerging Public-Health Concern. Lancet Infect. Dis. 2008, 8, 159–166. [Google Scholar] [CrossRef]
- Smet, A.; Martel, A.; Persoons, D.; Dewulf, J.; Heyndrickx, M.; Herman, L.; Haesebrouck, F.; Butaye, P. Broad-Spectrum β-Lactamases among Enterobacteriaceae of Animal Origin: Molecular Aspects, Mobility and Impact on Public Health. FEMS Microbiol. Rev. 2010, 34, 295–316. [Google Scholar] [CrossRef] [Green Version]
- Ewers, C.; Bethe, A.; Semmler, T.; Guenther, S.; Wieler, L.H. Extended-Spectrum β-Lactamase-Producing and AmpC-Producing Escherichia Coli from Livestock and Companion Animals, and Their Putative Impact on Public Health: A Global Perspective. Clin. Microbiol. Infect. 2012, 18, 646–655. [Google Scholar] [CrossRef] [Green Version]
- Jouini, A.; Vinué, L.; Slama, K.B.; Sáenz, Y.; Klibi, N.; Hammami, S.; Boudabous, A.; Torres, C. Characterization of CTX-M and SHV Extended-Spectrum β-Lactamases and Associated Resistance Genes in Escherichia Coli Strains of Food Samples in Tunisia. J. Antimicrob. Chemother. 2007, 60, 1137–1141. [Google Scholar] [CrossRef] [Green Version]
- Overdevest, I.; Willemsen, I.; Rijnsburger, M.; Eustace, A.; Xu, L.; Hawkey, P.; Heck, M.; Savelkoul, P.; Vandenbroucke-Grauls, C.; van der Zwaluw, K.; et al. Extended-Spectrum β-Lactamase Genes of Escherichia Coli in Chicken Meat and Humans, The Netherlands. Emerg. Infect. Dis. 2011, 17, 1216–1222. [Google Scholar] [CrossRef]
- Leverstein-van Hall, M.A.; Dierikx, C.M.; Cohen Stuart, J.; Voets, G.M.; van den Munckhof, M.P.; van Essen-Zandbergen, A.; Platteel, T.; Fluit, A.C.; van de Sande-Bruinsma, N.; Scharinga, J.; et al. Dutch Patients, Retail Chicken Meat and Poultry Share the Same ESBL Genes, Plasmids and Strains. Clin. Microbiol. Infect. 2011, 17, 873–880. [Google Scholar] [CrossRef] [Green Version]
- Kola, A.; Kohler, C.; Pfeifer, Y.; Schwab, F.; Kühn, K.; Schulz, K.; Balau, V.; Breitbach, K.; Bast, A.; Witte, W.; et al. High Prevalence of Extended-Spectrum-β-Lactamase-Producing Enterobacteriaceae in Organic and Conventional Retail Chicken Meat, Germany. J. Antimicrob. Chemother. 2012, 67, 2631–2634. [Google Scholar] [CrossRef] [Green Version]
- Dohmen, W.; Bonten, M.J.M.; Bos, M.E.H.; van Marm, S.; Scharringa, J.; Wagenaar, J.A.; Heederik, D.J.J. Carriage of Extended-Spectrum β-Lactamases in Pig Farmers Is Associated with Occurrence in Pigs. Clin. Microbiol. Infect. 2015, 21, 917–923. [Google Scholar] [CrossRef] [Green Version]
- National Research Council. Nutrient Requirements of Swine (NRC): Eleventh Revised Edition; The National Academies Press (NAP): Washington, DC, USA, 2012. [Google Scholar]
- Breakpoint Tables for Interpretation of MICs and Zone Diameters, Version 10.0; The European Committee on Antimicrobial Susceptibility Testing: Växjö, Sweden, 2020; 77.
- Braun, S.D.; Jamil, B.; Syed, M.A.; Abbasi, S.A.; Weiß, D.; Slickers, P.; Monecke, S.; Engelmann, I.; Ehricht, R. Prevalence of Carbapenemase-Producing Organisms at the Kidney Center of Rawalpindi (Pakistan) and Evaluation of an Advanced Molecular Microarray-Based Carbapenemase Assay. Future Microbiol. 2018, 13, 1225–1246. [Google Scholar] [CrossRef]
- Butiuc-Keul, A.; Carpa, R.; Podar, D.; Szekeres, E.; Muntean, V.; Iordache, D.; Farkas, A. Antibiotic Resistance in Pseudomonas spp. Through the Urban Water Cycle. Curr. Microbiol. 2021, 78, 1227–1237. [Google Scholar] [CrossRef]
- Gelman, A.; Carlin, J.B.; Stern, H.S.; Rubin, D.B. Bayesian Data Analysis; Chapman and Hall/CRC: New York, NY, USA, 1995; ISBN 978-0-429-25841-1. [Google Scholar]
- R Core Team (2020). European Environment Agency. Available online: https://www.eea.europa.eu/data-and-maps/indicators/oxygen-consuming-substances-in-rivers/r-development-core-team-2006 (accessed on 4 May 2022).
- Denwood, M.J. Runjags: An R Package Providing Interface Utilities, Model Templates, Parallel Computing Methods and Additional Distributions for MCMC Models in JAGS. J. Stat. Softw. 2016, 71, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. Ggplot2. WIREs Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]
- Lee, W.; Grimm, K.J. Generalized Linear Mixed-Effects Modeling Programs in R for Binary Outcomes. Struct. Equ. Modeling A Multidiscip. J. 2018, 25, 824–828. [Google Scholar] [CrossRef]
- Burow, E.; Rostalski, A.; Harlizius, J.; Gangl, A.; Simoneit, C.; Grobbel, M.; Kollas, C.; Tenhagen, B.-A.; Käsbohrer, A. Antibiotic Resistance in Escherichia Coli from Pigs from Birth to Slaughter and Its Association with Antibiotic Treatment. Prev. Vet. Med. 2019, 165, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, H.; Wang, L.; Peng, Q.; Li, Y.; Zhou, H.; Li, Q. Molecular Characterization of Extended-Spectrum β-Lactamase-Producing Multidrug Resistant Escherichia Coli from Swine in Northwest China. Front. Microbiol. 2018, 9, 1756. [Google Scholar] [CrossRef] [Green Version]
- Aguirre, L.; Vidal, A.; Seminati, C.; Tello, M.; Redondo, N.; Darwich, L.; Martín, M. Antimicrobial Resistance Profile and Prevalence of Extended-Spectrum Beta-Lactamases (ESBL), AmpC Beta-Lactamases and Colistin Resistance (Mcr) Genes in Escherichia Coli from Swine between 1999 and 2018. Porc. Health Manag. 2020, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Tamta, S.; Kumar, O.R.V.; Singh, S.V.; Pruthvishree, B.S.; Karthikeyan, R.; Rupner, R.; Sinha, D.K.; Singh, B.R. Antimicrobial Resistance Pattern of Extended-Spectrum β-Lactamase-Producing Escherichia Coli Isolated from Fecal Samples of Piglets and Pig Farm Workers of Selected Organized Farms of India. Vet. World 2020, 13, 360–363. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; An, J.-U.; Guk, J.-H.; Song, H.; Yi, S.; Kim, W.-H.; Cho, S. Prevalence, Characteristics and Clonal Distribution of Extended-Spectrum β-Lactamase- and AmpC β-Lactamase-Producing Escherichia Coli Following the Swine Production Stages, and Potential Risks to Humans. Front. Microbiol. 2021, 12, 710747. [Google Scholar] [CrossRef]
- Valiakos, G.; Vontas, A.; Tsokana, C.N.; Giannakopoulos, A.; Chatzopoulos, D.; Billinis, C. Resistance in Escherichia Coli Strains Isolated from Pig Faecal Samples and Pig Farm Workers, Greece. Am. J. Anim. Vet. Sci. 2016, 11, 142–144. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulos, D.; Papadopoulos, T.; Papageorgiou, K.; Sergelidis, D.; Adamopoulou, M.; Kritas, S.K.; Petridou, E. Antimicrobial Resistance Rates in Commensal Escherichia Coli Isolates from Healthy Pigs in Greek Swine Farms. J. Hell. Vet. Med. Soc. 2021, 72, 2909–2916. [Google Scholar] [CrossRef]
- Athanasakopoulou, Z.; Reinicke, M.; Diezel, C.; Sofia, M.; Chatzopoulos, D.C.; Braun, S.D.; Reissig, A.; Spyrou, V.; Monecke, S.; Ehricht, R.; et al. Antimicrobial Resistance Genes in ESBL-Producing Escherichia Coli Isolates from Animals in Greece. Antibiotics 2021, 10, 389. [Google Scholar] [CrossRef]
- Poirel, L.; Stephan, R.; Perreten, V.; Nordmann, P. The Carbapenemase Threat in the Animal World: The Wrong Culprit. J. Antimicrob. Chemother. 2014, 69, 2007–2008. [Google Scholar] [CrossRef] [Green Version]
- Scientific Opinion on Carbapenem Resistance in Food Animal Ecosystems|EFSA. Available online: https://www.efsa.europa.eu/en/efsajournal/pub/3501 (accessed on 4 May 2022).
- Yang, P.; Chen, Y.; Jiang, S.; Shen, P.; Lu, X.; Xiao, Y. Association between Antibiotic Consumption and the Rate of Carbapenem-Resistant Gram-Negative Bacteria from China Based on 153 Tertiary Hospitals Data in 2014. Antimicrob. Resist. Infect. Control. 2018, 7, 137. [Google Scholar] [CrossRef]
- Villar, H.E.; Danel, F.; Livermore, D.M. Permeability to Carbapenems of Proteus Mirabilis Mutants Selected for Resistance to Imipenem or Other Beta-Lactams. J. Antimicrob. Chemother. 1997, 40, 365–370. [Google Scholar] [CrossRef] [Green Version]
- Politi, L.; Tassios, P.T.; Lambiri, M.; Kansouzidou, A.; Pasiotou, M.; Vatopoulos, A.C.; Mellou, K.; Legakis, N.J.; Tzouvelekis, L.S. Repeated Occurrence of Diverse Extended-Spectrum β-Lactamases in Minor Serotypes of Food-Borne Salmonella Enterica Subsp. Enterica. J. Clin. Microbiol. 2005, 43, 3453–3456. [Google Scholar] [CrossRef] [Green Version]
- Rubira, I.; Figueras, L.P.; Jiménez, J.C.; de Arcaute, M.R.; Ruiz, H.; Ventura, J.A.; Lacasta, D. Salmonella Enterica Subsp. Diarizonae Serotype 61:K:1:5:(7) a Host Adapted to Sheep; IntechOpen: London, UK, 2021; ISBN ISBN 978-1-83969-018-1. [Google Scholar]
- Bernreiter-Hofer, T.; Schwarz, L.; Müller, E.; Cabal-Rosel, A.; Korus, M.; Misic, D.; Frankenfeld, K.; Abraham, K.; Grünzweil, O.; Weiss, A.; et al. The Pheno- and Genotypic Characterization of Porcine Escherichia Coli Isolates. Microorganisms 2021, 9, 1676. [Google Scholar] [CrossRef]
- Balázs, B.; Nagy, J.B.; Tóth, Z.; Nagy, F.; Károlyi, S.; Turcsányi, I.; Bistyák, A.; Kálmán, A.; Sárközi, R.; Kardos, G. Occurrence of Escherichia Coli Producing Extended Spectrum β-Lactamases in Food-Producing Animals. Acta Vet. Hung. 2021, 69, 211–215. [Google Scholar] [CrossRef]
- Giufrè, M.; Mazzolini, E.; Cerquetti, M.; Brusaferro, S.; Accogli, M.; Agnoletti, F.; Agodi, A.; Alborali, G.L.; Arghittu, M.; Auxilia, F.; et al. Extended-Spectrum β-Lactamase-Producing Escherichia Coli from Extraintestinal Infections in Humans and from Food-Producing Animals in Italy: A ‘One Health’ Study. Int. J. Antimicrob. Agents 2021, 58, 106433. [Google Scholar] [CrossRef]
- Madec, J.Y.; Haenni, M.; Nordmann, P.; Poirel, L. Extended-Spectrum β-Lactamase/AmpC- and Carbapenemase-Producing Enterobacteriaceae in Animals: A Threat for Humans? Clin. Microbiol. Infect. 2017, 23, 826–833. [Google Scholar] [CrossRef] [Green Version]
- Lekagul, A.; Tangcharoensathien, V.; Mills, A.; Rushton, J.; Yeung, S. How antibiotics are used in pig farming: A mixed-methods study of pig farmers, feed mills and veterinarians in Thailand. BMJ Glob. Health 2020, 5, e001918. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Filioussis, G.; Kachrimanidou, M.; Christodoulopoulos, G.; Kyritsi, M.; Hadjichristodoulou, C.; Adamopoulou, M.; Tzivara, A.; Kritas, S.K.; Grinberg, A. Short communication: Bovine mastitis caused by a multidrug-resistant, mcr-1-positive (colistin-resistant), extended-spectrum β-lactamase-producing Escherichia coli clone on a Greek dairy farm. J. Dairy Sci. 2020, 103, 852–857. [Google Scholar] [CrossRef]
- van der Fels-Klerx, H.J.; Puister-Jansen, L.F.; van Asselt, E.D.; Burgers, S.L.G.E. Farm Factors Associated with the Use of Antibiotics in Pig Production. J. Anim. Sci. 2011, 89, 1922–1929. [Google Scholar] [CrossRef] [Green Version]
- Ström, G.; Halje, M.; Karlsson, D.; Jiwakanon, J.; Pringle, M.; Fernström, L.-L.; Magnusson, U. Antimicrobial Use and Antimicrobial Susceptibility in Escherichia Coli on Small- and Medium-Scale Pig Farms in North-Eastern Thailand. Antimicrob. Resist. Infect. Control. 2017, 6, 75. [Google Scholar] [CrossRef] [Green Version]
- Chantziaras, I.; Boyen, F.; Callens, B.; Dewulf, J. Correlation between Veterinary Antimicrobial Use and Antimicrobial Resistance in Food-Producing Animals: A Report on Seven Countries. J. Antimicrob. Chemother. 2014, 69, 827–834. [Google Scholar] [CrossRef] [Green Version]
- Ricker, N.; Trachsel, J.; Colgan, P.; Jones, J.; Choi, J.; Lee, J.; Coetzee, J.F.; Howe, A.; Brockmeier, S.L.; Loving, C.L.; et al. Toward Antibiotic Stewardship: Route of Antibiotic Administration Impacts the Microbiota and Resistance Gene Diversity in Swine Feces. Front. Vet. Sci. 2020, 7, 255. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Region | Number of Farms | Total Capacity | Capacity per Farm | Number of Samples per Age | |||||
| 50–100 | 101–250 | 251–450 | 451–700 | 701–2100 | Suckling Piglets | Weaning Piglets | |||
| Northern Greece | 4 | 1640 | 0 | 2 | 1 | 1 | 0 | 8 | 19 |
| Central Greece | 13 | 3710 | 4 | 2 | 3 | 4 | 0 | 24 | 44 |
| Western Greece | 10 | 5790 | 0 | 4 | 2 | 2 | 2 | 22 | 42 |
| Southern Greece | 7 | 3190 | 0 | 4 | 2 | 0 | 1 | 19 | 36 |
| Total | 34 | 14,300 | 4 | 12 | 8 | 7 | 3 | 73 | 141 |
| Criteria | Farms | Sows | Weaners |
|---|---|---|---|
| Capacity | Minimum 50 sows | ||
| Type of farm | Exclusive farrow-to-finish | ||
| Antiparasitic treatment | Antiparasitic treatment (IM) before farrowing | ||
| Vaccination | Aujeszky’s disease virus Porcine parvovirus Atrophic rhinitis Erysipelas Porcine Reproductive and Respiratory Virus E. coli Clostridium spp. | Porcine Circovirus type 2 Mycoplasma hyopneumoniae | |
| Diet | Home-made diets (mixed corn/barley/wheat–soybean-based meal) balanced in dietary nutrients (essential amino acids, minerals and vitamins) 1 | ||
| Toxin binders | Systematically used in the feed during gestation and lactation | Systematically used in the feed | |
| Routine program for metaphylaxis of PPDS | Amoxicillin (IM) after farrowing | ||
| Routine program of metaphylaxis for PWD | Amoxicillin via feed for the first 10 days after weaning | ||
| Bacterial Species | Percentage of Isolates (n = 214) 1 | Percentage of ESBL-Producing Isolates (n = 78) 2 |
|---|---|---|
| Escherichia coli | 32.2 (n = 69) | 88.5 (n = 69) |
| Klebsiella pneumoniae spp. pneumoniae | 1.4 (n = 3) | 3.8 (n = 3) |
| Proteus mirabilis | 1.9 (n = 4) | 5.1 (n = 4) |
| Enterobacter cloacae | 0.5 (n = 1) | 1.3 (n = 1) |
| Salmonella enterica subsp. diarizonae | 0.5 (n = 1) | 1.3 (n = 1) |
| Total | 36.5 (n = 78) | 100 (n = 78) |
| Resistance Genes | Number of Isolates | ||||
|---|---|---|---|---|---|
| E. coli | K. pneumoniae | P. mirabilis | S. enterica ssp. diarizonae | E. cloacae Complex | |
| blaCTX-M1/15 | 52 | 2 | - | - | - |
| blaCTX-M9 | 6 | 1 | 3 | - | - |
| blaCTX-M8 | 6 | - | 1 | - | - |
| blaSHV | 3 | 2 | - | - | - |
| blaTEM | 38 | 1 | 3 | 1 | - |
| blaACT | 18 | - | - | - | - |
| blaCMY | 1 | - | - | - | - |
| blaOXA-1 | 17 | - | 4 | - | - |
| blaOXA-60 | 1 | - | - | - | - |
| aadA1 | 36 | 1 | 3 | 1 | - |
| aadA2 | 21 | - | 1 | 1 | - |
| aadA4 | 14 | - | - | 1 | - |
| aphA | 21 | 1 | 1 | 1 | - |
| rmtA | 27 | - | - | - | - |
| rmtC | 7 | - | - | - | - |
| aac(6′)-Ib | 2 | - | 3 | - | - |
| aac(3′)-Iva | 1 | - | 3 | - | - |
| qnrS | 29 | 3 | - | 1 | - |
| qnrA | 2 | - | - | - | - |
| qnrB | 4 | - | - | - | - |
| sul1 | 18 | 2 | 3 | - | - |
| sul2 | 38 | 3 | 4 | 1 | - |
| sul3 | 17 | - | - | 1 | - |
| dfrA1 | 13 | 1 | 3 | - | - |
| dfrA5 | 42 | - | 2 | - | 1 |
| dfrA7 | 4 | 1 | - | - | - |
| dfrA12 | 12 | - | - | 1 | - |
| dfrA13 | 2 | - | - | - | - |
| dfrA14 | 12 | 1 | - | - | - |
| dfrA15 | 1 | - | - | - | - |
| dfrA17 | 15 | 1 | 1 | 1 | - |
| dfrA19 | 4 | - | - | - | - |
| mcr-1/mcr-2 | 6 | - | - | 1 | - |
| mcr-4 | 1 | - | - | - | - |
| mcr-8 | 3 | - | - | 1 | - |
| mph | 12 | - | - | - | - |
| mrx | 16 | - | - | - | - |
| intl1 | 20 | 2 | - | - | - |
| intl2 | 7 | 1 | 3 | - | - |
| tnpISEcp1 | 30 | 3 | - | 1 | - |
| oqxA | 1 | 3 | - | - | - |
| oqxB | 1 | 3 | - | - | - |
| ESBL-Producing | Logistic Regression | |||
|---|---|---|---|---|
| Parameter | Category | Estimate (95% CI) | p Value | |
| E. coli | PMLs | 0 | 1 | |
| 1 | 0.25 (0.07; 0.84) | 0.0215 | ||
| P. mirabilis | Size | - | 85.86 (4.02; 6620.06) | 0.00648 |
| Antibiotic | Logistic Regression | |||
|---|---|---|---|---|
| Parameter | Category | Estimate (95% CI) | p Value (Random Effect) | |
| AMP | Size | 48.72 (3.21; 1955.14) | 0.0171 | |
| AMC | Size | 31.38 (5.3; 290.81) | <0.005 | |
| IMI | Size | 33.09 (2.16; 1454.48) | 0.008 | |
| GEN | PMLs | 0 | 1 | |
| 1 | 0.15 (0.02; 0.58) | 0.016 | ||
| Quinolones | 0 | 1 | ||
| 1 | 13.14 (2.46; 243.71) | 0.015 | ||
| NEO | PMLs | 0 | 1 | |
| 1 | 0.19 (0.03; 1.05) | 0.028 | ||
| TETs | 0 | 1 | ||
| 1 | 7.7 (1.62; 59.44) | 0.009 | ||
| MRX | TETs | 0 | 1 | |
| 1 | 14.45 (0.96; 5.59) | 0.011 | ||
| ENR | Cephalosporins | 0 | 1 | |
| 1 | 3.49 (1.07; 14.84) | 0.045 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsekouras, N.; Athanasakopoulou, Z.; Diezel, C.; Kostoulas, P.; Braun, S.D.; Sofia, M.; Monecke, S.; Ehricht, R.; Chatzopoulos, D.C.; Gary, D.; et al. Cross-Sectional Survey of Antibiotic Resistance in Extended Spectrum β-Lactamase-Producing Enterobacteriaceae Isolated from Pigs in Greece. Animals 2022, 12, 1560. https://doi.org/10.3390/ani12121560
Tsekouras N, Athanasakopoulou Z, Diezel C, Kostoulas P, Braun SD, Sofia M, Monecke S, Ehricht R, Chatzopoulos DC, Gary D, et al. Cross-Sectional Survey of Antibiotic Resistance in Extended Spectrum β-Lactamase-Producing Enterobacteriaceae Isolated from Pigs in Greece. Animals. 2022; 12(12):1560. https://doi.org/10.3390/ani12121560
Chicago/Turabian StyleTsekouras, Nikolaos, Zoi Athanasakopoulou, Celia Diezel, Polychronis Kostoulas, Sascha D. Braun, Marina Sofia, Stefan Monecke, Ralf Ehricht, Dimitris C. Chatzopoulos, Dominik Gary, and et al. 2022. "Cross-Sectional Survey of Antibiotic Resistance in Extended Spectrum β-Lactamase-Producing Enterobacteriaceae Isolated from Pigs in Greece" Animals 12, no. 12: 1560. https://doi.org/10.3390/ani12121560