
Prevalence and Virulence Characteristics of Enterococcus faecalis and Enterococcus faecium in Bovine Mastitis Milk Compared to Bovine Normal Raw Milk in South Korea

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation of E. faecalis and E. faecium
2.2. DNA Extraction and Screening for Virulence Genes
2.3. Phenotype of Virulence Characteristics
2.3.1. Biofilm Formation
2.3.2. Gelatinase Production
2.3.3. Hemolysin Production
2.4. Antimicrobial Resistance
2.5. Statistical Analysis
3. Results
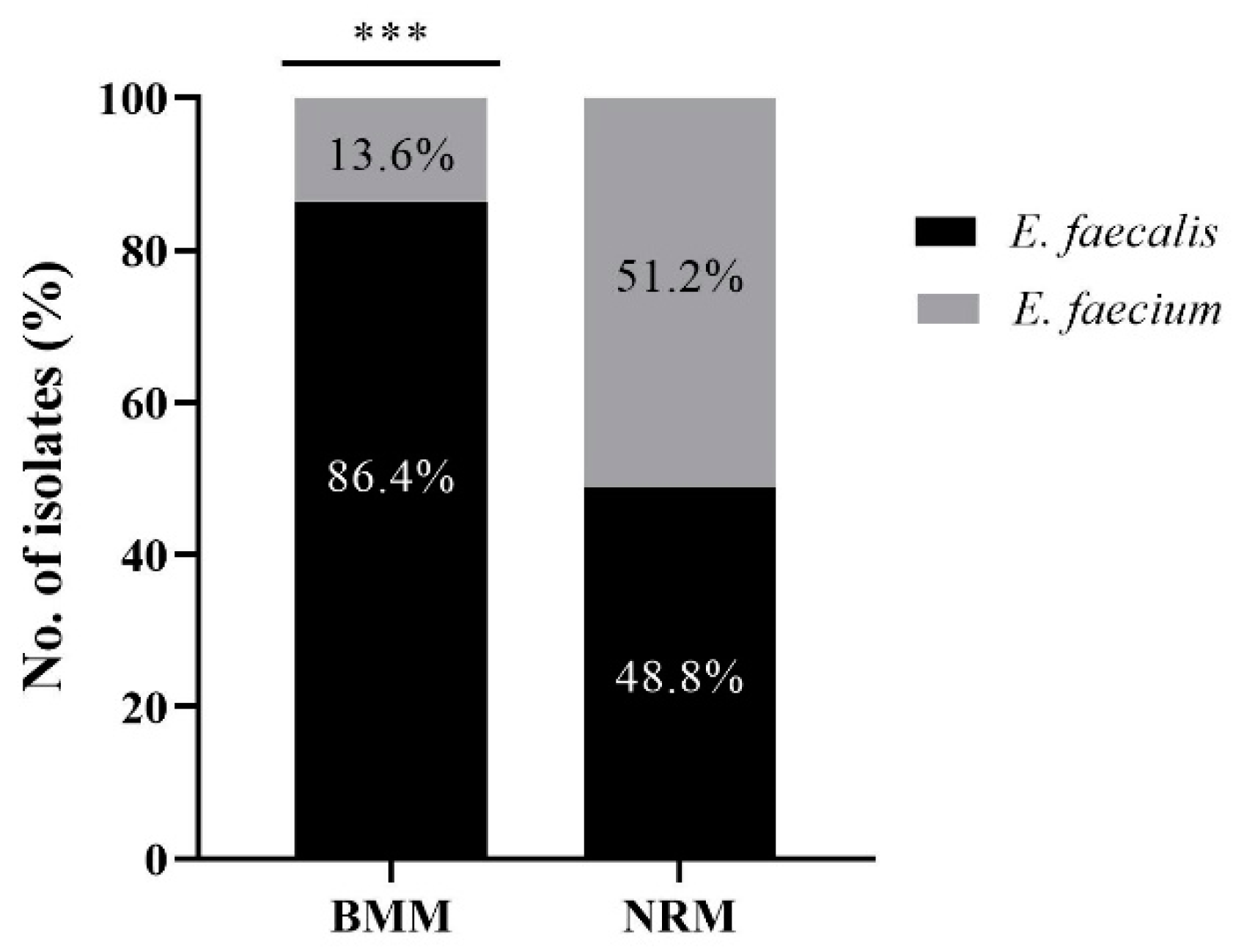
3.1. Identification of E. faecalis and E. faecium from BMM and NRM
3.2. Detection of Virulence Genes
3.3. Biofilm, Gelatinase, and Hemolysin Production
3.4. Correlation between Virulence Genes and Corresponding Phenotype
3.5. Antimicrobial Resistance of E. faecalis and E. faecium
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hogeveen, H.; Huijps, K.; Lam, T.J.G.M. Economic aspects of mastitis: New developments. N. Z. Vet. J. 2011, 59, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Riekerink, R.G.O.; Barkema, H.W.; Veenstra, S.; Poole, D.E.; Dingwell, R.T.; Keefe, G.P. Prevalence of contagious mastitis pathogens in bulk tank milk in Prince Edward Island. Can. Vet. J. 2006, 47, 567. [Google Scholar]
- Różańska, H.; Lewtak-Piłat, A.; Kubajka, M.; Weiner, M. Occurrence of enterococci in mastitic cow’s milk and their antimicrobial resistance. J. Vet. Res. 2019, 63, 93–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiruthiga, A.; Padmavathy, K.; Shabana, P.; Naveenkumar, V.; Gnanadesikan, S.; Malaiyan, J. Improved detection of esp, hyl, asa1, gelE, cylA virulence genes among clinical isolates of Enterococci. BMC Res. Notes. 2020, 13, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, C.M.; Holzapfel, W.H.; Stiles, M.E. Enterococci at the crossroads of food safety? Int. J. Food Microbiol. 1999, 47, 1–24. [Google Scholar] [CrossRef]
- Nam, H.M.; Lim, S.K.; Kang, H.M.; Kim, J.M.; Moon, J.S.; Jang, K.C.; Jung, S.C. Prevalence and antimicrobial susceptibility of gram-negative bacteria isolated from bovine mastitis between 2003 and 2008 in Korea. Int. J. Dairy Sci. 2009, 92, 2020–2026. [Google Scholar] [CrossRef] [Green Version]
- Golob, M.; Pate, M.; Kušar, D.; Dermota, U.; Avberšek, J.; Papić, B.; Zdovc, I. Antimicrobial resistance and virulence genes in Enterococcus faecium and Enterococcus faecalis from humans and retail red meat. Biomed. Res. Int. 2019, 2019, 2815279. [Google Scholar] [CrossRef] [Green Version]
- Kang, I.B.; Kim, D.H.; Chon, J.W.; Seo, K.H. Effect of microbial control measures on farmstead cheesemaking and antimicrobial resistance of Staphylococcus aureus and Enterococcus spp. isolates. J. Food Saf. 2018, 38, e12432. [Google Scholar] [CrossRef]
- Murray, B.E.; Miller, W.R. Treatment of Enterococcal Infections; UpToDate: Waltham, MA, USA, 2018; Available online: https://www.uptodate.com/contents/treatment-of-enterococcal-infections (accessed on 31 May 2021).
- Kim, S.H.; Kim, D.H.; Lim, H.W.; Seo, K.H. High prevalence of non-faecalis and non-faecium Enterococcus spp. in farmstead cheesehouse and their applicability as hygiene indicators. LWT 2020, 126, 109271. [Google Scholar] [CrossRef]
- Eaton, T.J.; Gasson, M.J. Molecular screening of Enterococcus virulence determinants and potential for genetic exchange between food and medical isolates. Appl. Environ. Microbiol. 2001, 67, 1628–1635. [Google Scholar] [CrossRef] [Green Version]
- Mah, T.F. Biofilm-specific antibiotic resistance. Future Microbiol. 2012, 7, 1061–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, J.A.; Huang, D.B. Biofilm formation by enterococci. J. Med. Microbiol. 2007, 56, 1581–1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, J.; Teng, F.; Murray, B.E. Gelatinase is important for translocation of Enterococcus faecalis across polarized human enterocyte-like T84 cells. Infect. Immun. 2005, 73, 1606–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coburn, P.S.; Gilmore, M.S. The Enterococcus faecalis cytolysin: A novel toxin active against eukaryotic and prokaryotic cells. Cell. Microbiol. 2003, 5, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.; Lee, Y.J. Molecular Characteristics of Enterococcus faecalis and Enterococcus faecium from Bulk Tank Milk in Korea. Animals 2021, 11, 661. [Google Scholar] [CrossRef] [PubMed]
- Vankerckhoven, V.; Van Autgaerden, T.; Vael, C.; Lammens, C.; Chapelle, S.; Rossi, R.; Goossens, H. Development of a multiplex PCR for the detection of asa1, gelE, cylA, esp, and hyl genes in enterococci and survey for virulence determinants among European hospital isolates of Enterococcus faecium. J. Clin. Microbiol. 2004, 42, 4473–4479. [Google Scholar] [CrossRef] [Green Version]
- Stepanović, S.; Vuković, D.; Hola, V.; Bonaventura, G.D.; Djukić, S.; Ćirković, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. Apmis 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Clinical & Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Supplement M100-S3011; CLSI: Wayne, PA, USA, 2020. [Google Scholar]
- Yun, M.J.; Yoon, S.; Lee, Y.J. Monitoring and characteristics of major mastitis pathogens from bulk tank milk in Korea. Animals 2020, 10, 1562. [Google Scholar] [CrossRef]
- Lee, S.H.I.; Camargo, C.H.; Gonçalves, J.L.; Cruz, A.G.; Sartori, B.T.; Machado, M.B.; Oliveira, C.A.F. Characterization of Staphylococcus aureus isolates in milk and the milking environment from small-scale dairy farms of São Paulo, Brazil, using pulsed-field gel electrophoresis. J. Dairy Sci. 2012, 95, 7377–7383. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.N.; Han, S.G. Bovine mastitis: Risk factors, therapeutic strategies, and alternative treatments. Asian-Australas. J. Anim. Sci. 2020, 33, 1699. [Google Scholar] [CrossRef]
- Coque, T.M.; Patterson, J.E.; Steckelberg, J.M.; Murray, B.E. Incidence of hemolysin, gelatinase, and aggregation substance among enterococci isolated from patients with endocarditis and other infections and from feces of hospitalized and community-based persons. J. Infect. Dis. 1995, 171, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, A.W.; Pereira, R.I.; Oliveira, D.V.D.; Martins, P.D.; d’Azevedo, P.A.; Van der Sand, S.; Frazzon, A.P.G. Molecular detection of virulence factors among food and clinical Enterococcus faecalis strains in South Brazil. Braz. J. Microbiol. 2014, 45, 327–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maasjost, J.; Lüschow, D.; Kleine, A.; Hafez, H.M.; Mühldorfer, K. Presence of virulence genes in Enterococcus species isolated from meat turkeys in Germany does not correlate with chicken embryo lethality. Biomed. Res. Int. 2019, 2019, 6147695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Tyne, D.; Martin, M.J.; Gilmore, M.S. Structure, function, and biology of the Enterococcus faecalis cytolysin. Toxins 2013, 5, 895–911. [Google Scholar] [CrossRef] [Green Version]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control. 2019, 8, 76. [Google Scholar] [CrossRef]
- Kafil, H.S.; Mobarez, A.M. Assessment of biofilm formation by enterococci isolates from urinary tract infections with different virulence profiles. J. King Saud Univ. Sci. 2015, 27, 312–317. [Google Scholar] [CrossRef] [Green Version]
- Cui, P.; Feng, L.; Zhang, L.; He, J.; An, T.; Fu, X.; Yang, X. Antimicrobial Resistance, Virulence Genes, and Biofilm Formation Capacity Among Enterococcus species From Yaks in Aba Tibetan Autonomous Prefecture, China. Front. Microbiol. 2020, 11, 1250. [Google Scholar] [CrossRef]
- Soares, R.O.; Fedi, A.C.; Reiter, K.C.; Caierão, J.; d’Azevedo, P.A. Correlation between biofilm formation and gelE, esp, and agg genes in Enterococcus spp. clinical isolates. Virulence 2014, 5, 634–637. [Google Scholar] [CrossRef] [Green Version]
- Heikens, E.; Bonten, M.J.; Willems, R.J. Enterococcal surface protein Esp is important for biofilm formation of Enterococcus faecium E1162. J. Bacteriol. Res. 2007, 189, 8233–8240. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Dong, M.; Zheng, J.; Song, Q.; Yin, W.; Li, J.; Niu, W. Relationship of biofilm formation and gelE gene expression in Enterococcus faecalis recovered from root canals in patients requiring endodontic retreatment. J. Endod. 2011, 37, 631–636. [Google Scholar] [CrossRef]
- Kim, Y.B.; Seo, K.W.; Shim, J.B.; hyun Son, S.; Noh, E.B.; Lee, Y.J. Molecular characterization of antimicrobial-resistant Enterococcus faecalis and Enterococcus faecium isolated from layer parent stock. Poult. Sci. 2019, 98, 5892–5899. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Koo, M. Occurrence, Antimicrobial Resistance and Molecular Diversity of Enterococcus faecium in Processed Pork Meat Products in Korea. Foods 2020, 9, 1283. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lv, Y.; Cai, J.; Schwarz, S.; Cui, L.; Hu, Z.; Shen, J. A novel gene, optrA, that confers transferable resistance to oxazolidinones and phenicols and its presence in Enterococcus faecalis and Enterococcus faecium of human and animal origin. J. Antimicrob. Chemother. 2015, 70, 2182–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pankey, G.; Ashcraft, D.; Patel, N. In vitro synergy of daptomycin plus rifampin against Enterococcus faecium resistant to both linezolid and vancomycin. J. Antimicrob. Chemother. 2005, 49, 5166–5168. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Milk Type | No. of Isolates | No of Genes Encoding Virulence Factors (%) | |||
|---|---|---|---|---|---|
| esp *** | asa1 | gelE *** | cylA *** | ||
| BMM | 70 | 38 (54.3%) | 50 (71.4%) | 60 (85.7%) | 21 (30.0%) |
| NRM | 20 | 5 (25.0%) | 16 (80.0%) | 12 (60.0%) | 0 |
| Virulence Factor | Encoding Gene | Phenotype and Genotype of Isolates from BMM and NRM | |||||||
|---|---|---|---|---|---|---|---|---|---|
| P+G+ | P−G+ | P+G− | P−G− | ||||||
| BMM | NRM | BMM | NRM | BMM | NRM | BMM | NRM | ||
| Strong biofilm formation (=OD > 0.208) | esp * | 35 | 5 | 3 | 0 | 18 | 6 | 25 | 30 |
| asa1 | 37 | 5 | 14 | 11 | 16 | 6 | 14 | 19 | |
| gelE * | 43 | 6 | 18 | 6 | 10 | 5 | 10 | 24 | |
| Gelatinase | gelE * | 18 | 2 | 43 | 10 | 0 | 0 | 20 | 29 |
| Hemolysin | cylA | 2 | 0 | 19 | 0 | 0 | 0 | 60 | 41 |
| Antimicrobial Agents | No (%) of Antimicrobial Resistance Isolates from BMM and NRM | |||||
|---|---|---|---|---|---|---|
| BMM | NRM | |||||
| E. faecalis (n = 70) | E. faecium (n = 11) | Total (n = 81) | E. faecalis (n = 20) | E. faecium (n = 21) | Total (n = 41) | |
| AMP | 0 | 0 | 0 | 0 | 0 | 0 |
| C | 16 (22.9%) | 1 (9.1%) | 17 (21.0%) | 0 | 0 | 0 |
| CIP | 0 | 1 (9.1%) | 1 (1.2%) | 0 | 0 | 0 |
| DOX | 3 (4.3%) | 1 (9.1%) | 4 (4.9%) | 0 | 0 | 0 |
| TET | 45 (64.3%) | 3 (27.3%) | 48 (59.3%) | 4 (20%) | 3 (14.3%) | 7 (17.1%) |
| ERY | 19 (27.1%) | 3 (27.3%) | 22 (27.2%) | 1 (5%) | 13 (61.9%) | 14 (34.1%) |
| N | 0 | 1 (9.1%) | 1 (1.2%) | 0 | 0 | 0 |
| RIF | 11 (15.7%) | 4 (36.4%) | 15 (18.5%) | 1 (5%) | 5 (23.8%) | 6 (14.6%) |
| VAN | 0 | 0 | 0 | 0 | 0 | 0 |
| MDR | 12 (17.1%) | 1 (9.1%) | 13 (16.0%) | 0 | 2 (9.5%) | 2 (4.9%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-J.; Youn, H.-Y.; Kang, H.-J.; Moon, J.-S.; Jang, Y.-S.; Song, K.-Y.; Seo, K.-H. Prevalence and Virulence Characteristics of Enterococcus faecalis and Enterococcus faecium in Bovine Mastitis Milk Compared to Bovine Normal Raw Milk in South Korea. Animals 2022, 12, 1407. https://doi.org/10.3390/ani12111407
Kim H-J, Youn H-Y, Kang H-J, Moon J-S, Jang Y-S, Song K-Y, Seo K-H. Prevalence and Virulence Characteristics of Enterococcus faecalis and Enterococcus faecium in Bovine Mastitis Milk Compared to Bovine Normal Raw Milk in South Korea. Animals. 2022; 12(11):1407. https://doi.org/10.3390/ani12111407
Chicago/Turabian StyleKim, Hyeon-Jin, Hye-Young Youn, Hye-Jeong Kang, Jin-San Moon, Yong-Seok Jang, Kwang-Young Song, and Kun-Ho Seo. 2022. "Prevalence and Virulence Characteristics of Enterococcus faecalis and Enterococcus faecium in Bovine Mastitis Milk Compared to Bovine Normal Raw Milk in South Korea" Animals 12, no. 11: 1407. https://doi.org/10.3390/ani12111407