
Adaptive Responses of Thyroid Hormones, Insulin, and Glucose during Pregnancy and Lactation in Dairy Cows

 , , ,
, , , {kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Breeding
2.2. Samples
2.3. Statistical Analyses
3. Results
3.1. Pregnant and Nonpregnant Cows
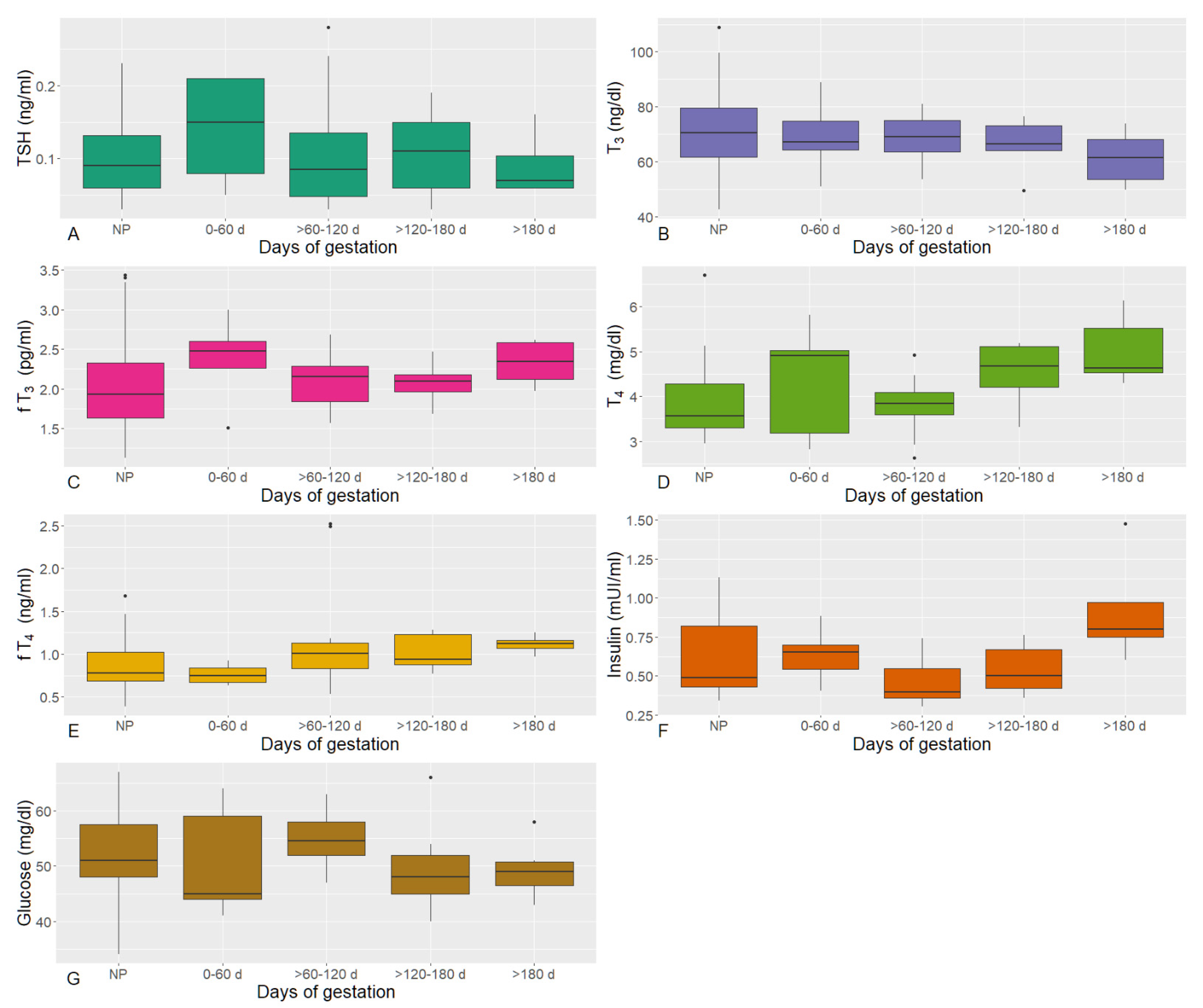
3.2. Lactating Phases
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsujii, H. Cystic ovarian disease and thyroid hormones in dairy cattle. Approaches Poult. Dairy Vet. Sci. 2017, 1, 256–261. [Google Scholar] [CrossRef]
- Nixon, D.A.; Akasha, M.A.; Anderson, R.R. Free and total thyroid hormones in serum of Holstein cows. J. Dairy Sci. 1988, 71, 1152–1160. [Google Scholar] [CrossRef]
- Tiirats, T. Thyroxine, triiodothyronine and reverse-triiodothyronine concentrations in blood plasma in relation to lactational stage, milk yield, energy and dietary protein intake in Estonian dairy cows. Acta Vet. Scand. 1997, 38, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Aceves, C.; Ruiz, A.; Romero, C.; Valverde, C. Homeorhesis during early lactation. Euthyroid sick-like syndrome in lactating cows. Eur. J. Endocrinol. 1985, 110, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Mutinati, M.; Rizzo, A.; Sciorsci, R.L. Cystic ovarian follicles and thyroid activity in the dairy cow. Anim. Reprod. Sci. 2013, 138, 150–154. [Google Scholar] [CrossRef]
- Spicer, L.J.; Alonso, J.; Chamberlain, C.S. Effects of thyroid hormones on bovine granulosa and thecal cell function in vitro: Dependence on insulin and gonadotropins. J. Dairy Sci. 2001, 84, 1069–1076. [Google Scholar] [CrossRef]
- Gregoraszczuk, E.L.; Galas, J. In Vitro Effect of Triiodothyronine on the Cyclic AMP, Progesterone and Testosterone Level in Porcine Theca, Granulosa and Luteal Cells. Endocr. Regul. 1998, 32, 93–98. [Google Scholar]
- Huszenicza, G.Y.; Kulcsar, M.; Rudas, P. Clinical endocrinology of thyroid gland function in ruminants. Vet. Med. Praha 2002, 47, 199–210. [Google Scholar]
- Stojic, V.; Gvozdic, D.; Kirovski, D.; Judith, N.A.; Huszenicza, G.Y.; Samanc, H.; Ivanov, I. Serum thyroxine and triiodothyronine concentrations prior to and after delivery in primiparous Holstein cows. Acta Vet. Beogr. 2001, 51, 3–8. [Google Scholar]
- Huszenicza, G.Y.; Kulcsar, M.; Nikolic, J.A.; Schmidt, J.; Korodi, P.; Katai, L.; Dieleman, S.; Ribiczei-Szabo, P.; Rudas, P. Plasma leptin concentration and its interrelation with some blood metabolites, metabolic hormones and the resumption of cyclic ovarian function in postpartum dairy cows supplemented with monensin or inert fat in feed. BSAP Occas. Publ. 2001, 26, 405–409. [Google Scholar] [CrossRef]
- Sinka, K.; Illek, J.; Kumprechtová, D.; Novák, P. Changes in T3 and T4 plasma concentrations in dairy cows during lactation. Jubil. World Buiatrics Congr. 2008, 283. [Google Scholar]
- Kirovski, D.; Sladojevic, Z.; Stojic, V.; Vujanac, I.; Lazarevic, M.; Radovanovic, A.; Savic, D.; Nedic, O. Effect of peripartum dietary energy supplementation on thyroid hormones, insulin-like growth factor-i and its binding proteins in early lactation dairy cows. Acta Vet. 2012, 62, 403–419. [Google Scholar] [CrossRef] [Green Version]
- Capuco, A.V.; Wood, D.L.; Elsasser, T.H.; Kahl, S.; Erdman, R.A.; Van Tassell, C.P.; Lefcourt, A.; Piperova, L.S. Effect of somatotropin on thyroid hormones and cytokines in lactating dairy cows during ad libitum and restricted feed intake. J. Dairy Sci. 2001, 84, 2430–2439. [Google Scholar] [CrossRef]
- Kunz, P.L.; Blum, J.W.; Hart, I.C.; Bickel, H.; Landis, J. Effects of different energy intakes before and after calving on food intake, performance and blood hormones and metabolites in dairy cows. Anim. Sci. 1985, 40, 219–231. [Google Scholar] [CrossRef] [Green Version]
- Nikolič, J.; Šamanc, H.; Begovic, J.; Damjanovic, Z.; Dokovic, R.; Kostic, G.; Kostic, G.; Krsmanovic, J.; Resanovic, V. Low peripheral serum thyroid hormone status independently affects the hormone profiles of healthy and ketotic cows during the first post partum. Acta Vet. 1997, 47, 3–13. [Google Scholar]
- Pucci, E.; Chiovato, L.; Pinchera, A. Thyroid and lipid metabolism. Int. J. Obes. Relat. Metab. Disord. 2000, 24, S109–S112. [Google Scholar] [CrossRef] [Green Version]
- Pethes, G.; Bokori, J.; Rudas, P.; Frenyó, V.L.; Fekete, S. Thyroxine, triiodothyronine, reverse-triiodothyronine, and other physiological characteristics of periparturient cows fed restricted energy. J. Dairy Sci. 1985, 68, 1148–1154. [Google Scholar] [CrossRef]
- Ronge, H.; Blum, J.; Clement, C.; Jans, F.; Leuenberger, H.; Binder, H. Somatomedin C in dairy cows related to energy and protein supply and to milk production. Anim. Sci. 1988, 47, 165–183. [Google Scholar] [CrossRef] [Green Version]
- McGuire, M.A.; Vicini, J.L.; Bauman, D.E.; Veenhuizen, J.J. Insulin-like growth factors and binding proteins in ruminants and their nutritional regulation. J. Anim. Sci. 1992, 70, 2901–2910. [Google Scholar] [CrossRef] [Green Version]
- Drackley, J.K. ADSA Foundation Scholar Award. Biology of dairy cows during the transition period: The final frontier? J. Dairy Sci. 1999, 82, 2259–2273. [Google Scholar] [CrossRef]
- Gross, J.; van Dorland, H.A.; Bruckmaier, R.M.; Schwarz, F.J. Performance and metabolic profile of dairy cows during a lactational and deliberately induced negative energy balance with subsequent realimentation. J. Dairy Sci. 2011, 94, 1820–1830. [Google Scholar] [CrossRef] [PubMed]
- Hayirli, A. The role of exogenous insulin in the complex of hepatic lipidosis and ketosis associated with insulin resistance phenomenon in postpartum dairy cattle. Vet. Res. Commun. 2006, 30, 749–774. [Google Scholar] [CrossRef] [PubMed]
- National Research Council (U.S.). Committee on Physiological Effects of Environmental Factors on Animals Guide to Environmental Research on Animals; National Academy of Sciences: Washington, DC, USA, 1971; ISBN 0309018692. [Google Scholar]
- Anderson, R.R.; Nixon, D.A.; Akasha, M.A. Total and free thyroxine and triiodothyronine in blood serum of mammals. Comp. Biochem. Physiol. A Comp. Physiol. 1988, 89, 401–404. [Google Scholar] [CrossRef]
- De Moraes, G.V.; Vera-Avila, H.R.; Lewis, A.W.; Koch, J.W.; Neuendorff, D.A.; Hallford, D.M.; Reeves, J.J.; Randel, R.D. Influence of hypo- or hyperthyroidism on ovarian function in Brahman cows. J. Anim. Sci. 1998, 76, 871–879. [Google Scholar] [CrossRef]
- Razavi, S.M.; Moghaddas, B.; Rakhshande, E.; Nazifi, S. Bovine Theileriosis: Effects on the Status of Thyroid Hormones, Homocystein, Serum Lipids and Lipoproteins. Res. J. Parasitol. 2015, 10, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Paulíková, I.; Seidel, H.; Nagy, O.; Tóthová, C.; Konvičná, J.; Kadaši, M.; Kováč, G. Thyroid Hormones, Insulin, Body Fat, and Blood Biochemistry Indices in Dairy Cows During the Reproduction/Production Cycle. Folia Vet. 2017, 61, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Du, M.; Tong, J.; Zhao, J.; Underwood, K.R.; Zhu, M.; Ford, S.P.; Nathanielsz, P.W. Fetal programming of skeletal muscle development in ruminant animals. J. Anim. Sci. 2010, 88, E51–E60. [Google Scholar] [CrossRef] [Green Version]
- Robinson, J.J.; McDonald, I.; Fraser, C.; Crofts, R.M.J. Studies on reproduction in prolific ewes. I. Growth of the products of conception. J. Agric. Sci. 1977, 88, 539–552. [Google Scholar] [CrossRef]
- Forhead, A.J.; Fowden, A.L. Thyroid hormones in fetal growth and prepartum maturation. J. Endocrinol. 2014, 221, R87–R103. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Carey, L.C.; Valego, N.K.; Liu, J.; Rose, J.C. Thyroid hormone modulates renin and ANG II receptor expression in fetal sheep. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R1006–R1014. [Google Scholar] [CrossRef]
- Lanham, S.A.; Fowden, A.L.; Roberts, C.; Cooper, C.; Oreffo, R.O.C.; Forhead, A.J. Effects of hypothyroidism on the structure and mechanical properties of bone in the ovine fetus. J. Endocrinol. 2011, 210, 189–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segar, J.L.; Volk, K.A.; Lipman, M.H.B.; Scholz, T.D. Thyroid hormone is required for growth adaptation to pressure load in the ovine fetal heart. Exp. Physiol. 2013, 98, 722–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, P.S.; Thorburn, G.D. Placental permeability to maternal thyroxine in the sheep. J. Endocrinol. 1971, 49, 549–550. [Google Scholar] [CrossRef]
- Dussault, J.H.; Hobel, C.J.; Distefano, J.J.; Erenberg, A.; Fisher, D.A. Triiodothyronine turnover in maternal and fetal sheep. Endocrinology 1972, 90, 1301–1308. [Google Scholar] [CrossRef] [PubMed]
- Steinhauser, C.B.; Askelson, K.; Hobbs, K.C.; Bazer, F.W.; Satterfield, M.C. Maternal nutrient restriction alters thyroid hormone dynamics in placentae of sheep having small for gestational age fetuses. Domest. Anim. Endocrinol. 2021, 77, 106632. [Google Scholar] [CrossRef] [PubMed]
- Liotta, L.; Bionda, A.; Quartuccio, M.; De Nardo, F.; Visalli, R.; Fazio, E. Thyroid and Lipidic Profiles in Nicastrese Goats (Capra hircus) during Pregnancy and Postpartum Period. Animals 2021, 11, 2386. [Google Scholar] [CrossRef] [PubMed]
- Fazio, E.; Medica, P.; Cravana, C.; Bruschetta, G.; Ferlazzo, A. Seasonal thyroid and lipid profiles in Thoroughbred pregnant and nonpregnant mares (Equus caballus). Theriogenology 2016, 85, 1582–1589. [Google Scholar] [CrossRef]
- Fazio, E.; Medica, P.; Trifiletti, C.; Ferlazzo, A. The outcome of the first stages of pregnancy on mares’ bloodstream thyroid hormones. Theriogenology 2016, 86, 1036–1041. [Google Scholar] [CrossRef]
- Fazio, E.; Medica, P.; Cravana, C.; Ferlazzo, A. Total and free iodothyronines profile in the donkey (Equus asinus) over a 12-month period. Acta Vet. Brno 2012, 81, 225–230. [Google Scholar] [CrossRef] [Green Version]
- Kelly, G.S. Peripheral metabolism of thyroid hormones: A review. Altern. Med. Rev. 2000, 5, 306–333. [Google Scholar]
- Riis, P.M.; Madsen, A. Thyroxine concentration and secretion rates in relation to pregnancy, lactation and energy balance in goats. J. Endocrinol. 1985, 107, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Koch, F.; Lamp, O.; Eslamizad, M.; Weitzel, J.; Kuhla, B. Metabolic Response to Heat Stress in Late-Pregnant and Early Lactation Dairy Cows: Implications to Liver-Muscle Crosstalk. PLoS ONE 2016, 11, e0160912. [Google Scholar] [CrossRef] [PubMed]
- Ramos, C.F.; Teixeira, C.V.; Passos, M.C.; Pazos Moura, C.C.; Lisboa, P.C.; Curty, F.H.; Moura, E.G. Low-protein diet changes thyroid function in lactating rats. Proc. Soc. Exp. Biol. Med. 2000, 224, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Brito, M.A.; González, F.D.; Ribeiro, L.A.; Campos, R.; Lacerda, L.; Barbosa, P.R.; Bergmann, G. Composição do sangue e do leite em ovinos leiteiros do sul do Brasil: Variações na gestação e na lactação. Ciênc. Rural 2006, 36, 942–948. [Google Scholar] [CrossRef] [Green Version]
- Fazio, E.; Medica, P.; Cravana, C.; Messineo, C.; Ferlazzo, A. Total and free iodothyronine levels of growing thoroughbred foals: Effects of weaning and gender. Livest. Sci. 2007, 110, 207–213. [Google Scholar] [CrossRef]
- Lucy, M.C. Mechanisms linking nutrition and reproduction in postpartum cows. Reprod. Suppl. 2003, 61, 415–427. [Google Scholar] [CrossRef]
- De Koster, J.D.; Opsomer, G. Insulin Resistance in Dairy Cows. Vet. Clin. Food Anim. Pract. 2013, 29, 299–322. [Google Scholar] [CrossRef]
- Bossaert, P.; Leroy, J.L.M.R.; De Vliegher, S.; Opsomer, G. Interrelations between glucose-induced insulin response, metabolic indicators, and time of first ovulation in high-yielding dairy cows. J. Dairy Sci. 2008, 91, 3363–3371. [Google Scholar] [CrossRef]
- Gross, J.J.; Bruckmaier, R.M. Invited review: Metabolic challenges and adaptation during different functional stages of the mammary gland in dairy cows: Perspectives for sustainable milk production. J. Dairy Sci. 2019, 102, 2828–2843. [Google Scholar] [CrossRef] [Green Version]
- Karis, P.; Jaakson, H.; Ling, K.; Bruckmaier, R.M.; Gross, J.J.; Pärn, P.; Kaart, T.; Ots, M. Body condition and insulin resistance interactions with periparturient gene expression in adipose tissue and lipid metabolism in dairy cows. J. Dairy Sci. 2020, 103, 3708–3718. [Google Scholar] [CrossRef]
- Collier, R.J.; Collier, J.L.; Rhoads, R.P.; Baumgard, L.H. Invited review: Genes involved in the bovine heat stress response. J. Dairy Sci. 2008, 91, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Magdub, A.; Johnson, H.D.; Belyea, R.L. Effect of environmental heat and dietary fiber on thyroid physiology of lactating cows. J. Dairy Sci. 1982, 65, 2323–2331. [Google Scholar] [CrossRef]
- Nardone, A.; Lacetera, N.; Bernabucci, U.; Ronchi, B. Composition of colostrum from dairy heifers exposed to high air temperatures during late pregnancy and the early postpartum period. J. Dairy Sci. 1997, 80, 838–844. [Google Scholar] [CrossRef]
- Kahl, S.; Elsasser, T.H.; Rhoads, R.P.; Collier, R.J.; Baumgard, L.H. Environmental heat stress modulates thyroid status and its response to repeated endotoxin challenge in steers. Domest. Anim. Endocrinol. 2015, 52, 43–50. [Google Scholar] [CrossRef]
- Li, G.; Ali, I.S.; Currie, R.W. Insulin induces myocardial protection and Hsp70 localization to plasma membranes in rat hearts. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H1709–H1721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhoads, R.P.; Baumgard, L.H.; Suagee, J.K.; Sanders, S.R. Nutritional Interventions to Alleviate the Negative Consequences of Heat Stress. Adv. Nutr. 2013, 4, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Min, L.; Cheng, J.; Shi, B.; Yang, H.; Zheng, N.; Wang, J. Effects of heat stress on serum insulin, adipokines, AMP-activated protein kinase, and heat shock signal molecules in dairy cows. J. Zhejiang Univ. Sci. B 2015, 16, 541–548. [Google Scholar] [CrossRef] [Green Version]
- De Rensis, F.; Scaramuzzi, R.J. Heat stress and seasonal effects on reproduction in the dairy cow—a review. Theriogenology 2003, 60, 1139–1151. [Google Scholar] [CrossRef]
- Marai, I.F.M.; El-Darawany, A.A.; Fadiel, A.; Abdel-Hafez, M.A.M. Physiological traits as affected by heat stress in sheep—A review. Small Rumin. Res. 2007, 71, 1–12. [Google Scholar] [CrossRef]
- Brenta, G. Why can insulin resistance be a natural consequence of thyroid dysfunction? J. Thyroid Res. 2011, 2011, 152850. [Google Scholar] [CrossRef] [Green Version]
- Fiore, E.; Gianesella, M.; Arfuso, F.; Giudice, E.; Piccione, G.; Lora, M.; Stefani, A.; Morgante, M. Glucose infusion response on some metabolic parameters in dairy cows during transition period. Arch. Anim. Breed. 2014, 57, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Fiore, E.; Piccione, G.; Perillo, L.; Barberio, A.; Manuali, E.; Morgante, M.; Gianesella, M.; Fiore, E.; Piccione, G.; Perillo, L.; et al. Hepatic lipidosis in high-yielding dairy cows during the transition period: Haematochemical and histopathological findings. Anim. Prod. Sci. 2015, 57, 74–80. [Google Scholar] [CrossRef]
- Ospina, P.A.; Nydam, D.V.; Stokol, T.; Overton, T.R. Evaluation of nonesterified fatty acids and β-hydroxybutyrate in transition dairy cattle in the northeastern United States: Critical thresholds for prediction of clinical diseases. J. Dairy Sci. 2010, 93, 546–554. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fazio, E.; Bionda, A.; Chiofalo, V.; Crepaldi, P.; Lopreiato, V.; Medica, P.; Liotta, L. Adaptive Responses of Thyroid Hormones, Insulin, and Glucose during Pregnancy and Lactation in Dairy Cows. Animals 2022, 12, 1395. https://doi.org/10.3390/ani12111395
Fazio E, Bionda A, Chiofalo V, Crepaldi P, Lopreiato V, Medica P, Liotta L. Adaptive Responses of Thyroid Hormones, Insulin, and Glucose during Pregnancy and Lactation in Dairy Cows. Animals. 2022; 12(11):1395. https://doi.org/10.3390/ani12111395
Chicago/Turabian StyleFazio, Esterina, Arianna Bionda, Vincenzo Chiofalo, Paola Crepaldi, Vincenzo Lopreiato, Pietro Medica, and Luigi Liotta. 2022. "Adaptive Responses of Thyroid Hormones, Insulin, and Glucose during Pregnancy and Lactation in Dairy Cows" Animals 12, no. 11: 1395. https://doi.org/10.3390/ani12111395