
Molecular Characterization of 18S rDNA, ITS-1, ITS-2, and COI from Eimeria christenseni and E. arloingi in Goats from Shaanxi Province, Northwestern China

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Conventional Fecal Examination of Eimeria Species
2.3. Isolation of Oocysts for E. arloingi and E. christenseni
2.4. DNA Extraction
2.5. PCR Amplification
2.6. Sequence Analysis
2.7. Nucleotide Sequence Accession Numbers
3. Results
3.1. Conventional Fecal Examination of Eimeria Species
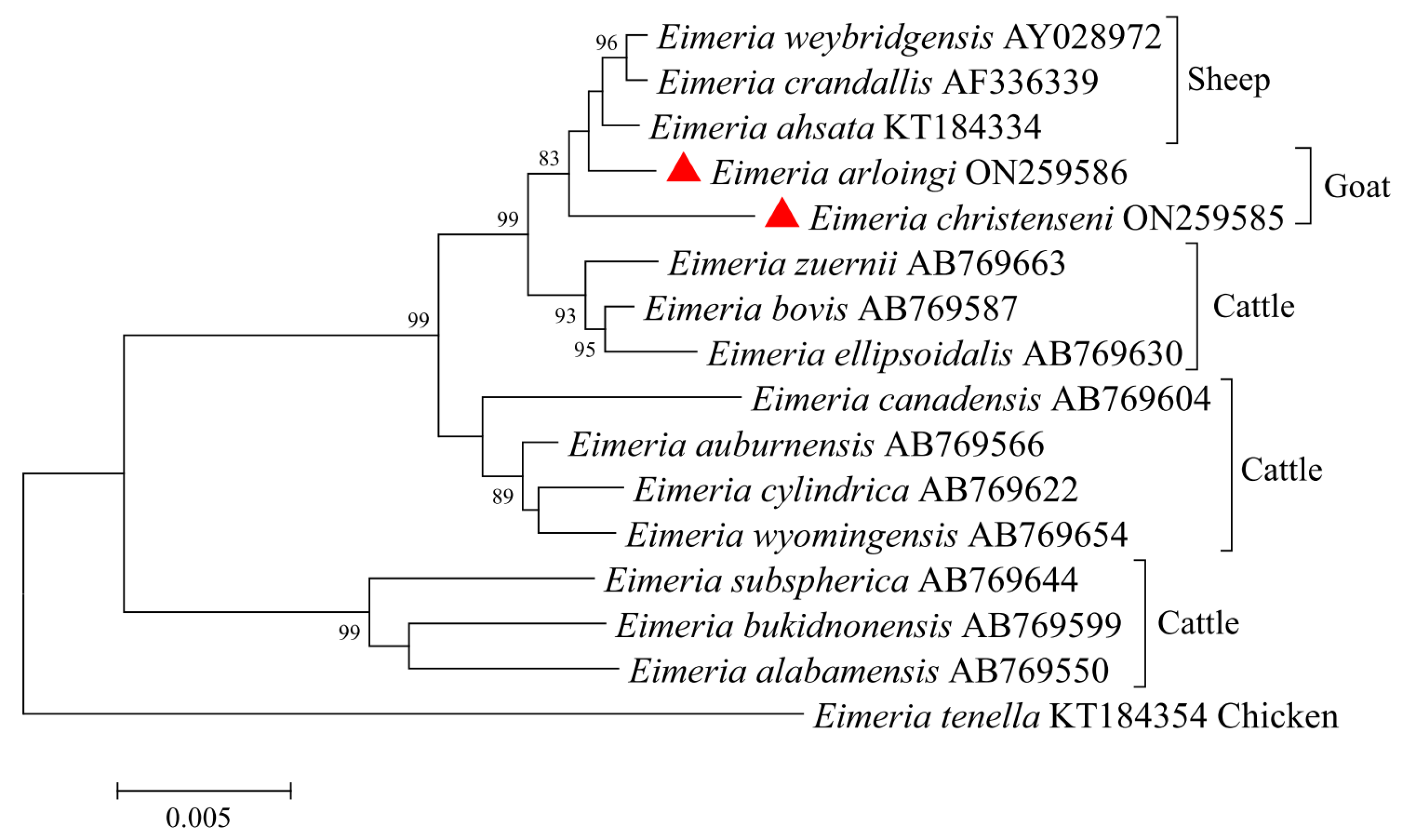
3.2. Characterization of 18S rDNA of E. christenseni and E. arloingi
3.3. Characterization of ITS-1 of E. christenseni and E. arloingi
3.4. Characterization of ITS-2 of E. christenseni and E. arloingi
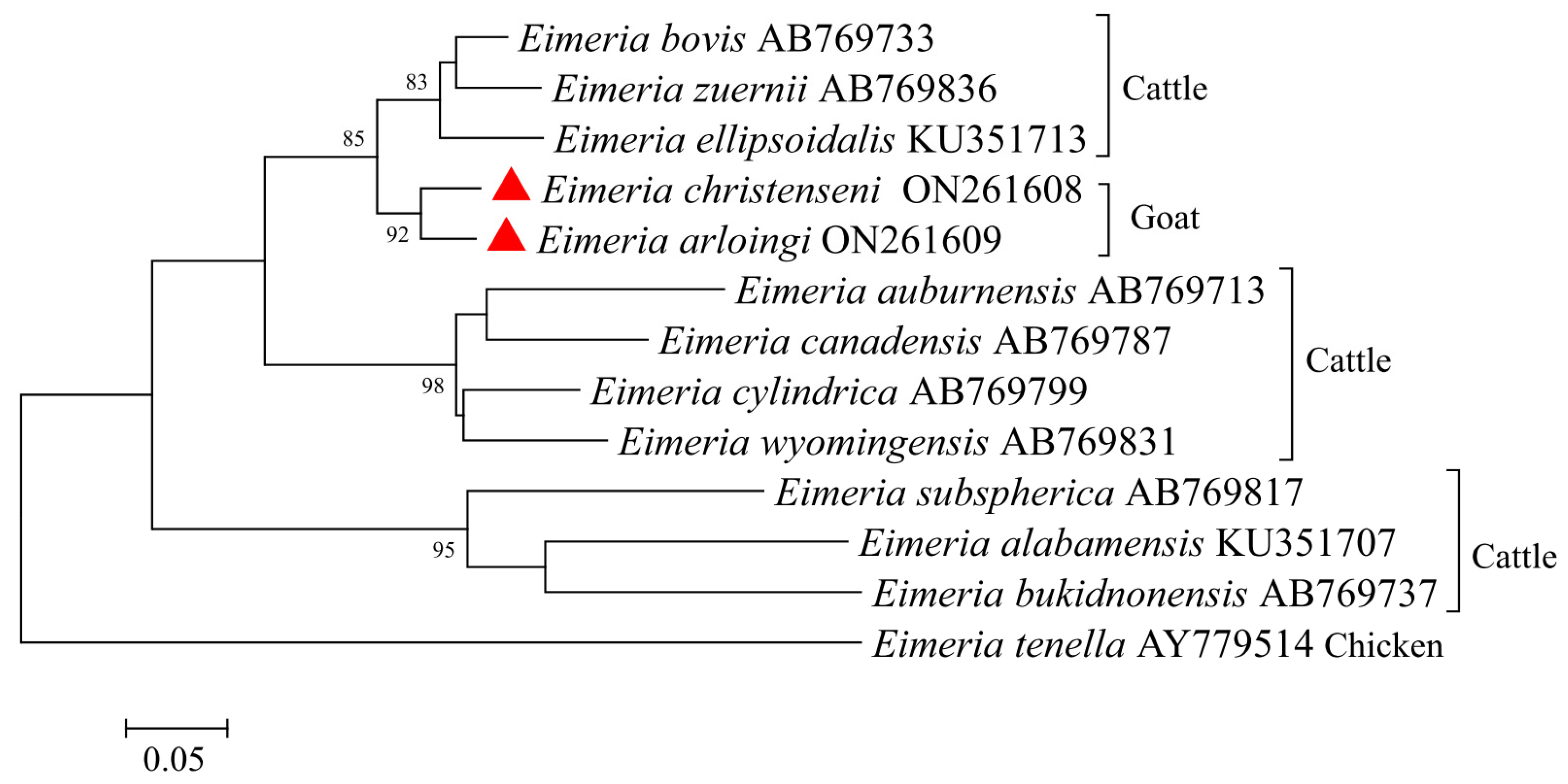
3.5. Characterization of COI of E. christenseni and E. arloingi
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blake, D.P.; Worthing, K.; Jenkins, M.C. Exploring Eimeria Genomes to Understand Population Biology: Recent Progress and Future Opportunities. Genes 2020, 11, 1103. [Google Scholar] [CrossRef] [PubMed]
- Sazmand, A.; Bahari, A.; Papi, S.; Otranto, D. Parasitic diseases of equids in Iran (1931–2020): A literature review. Parasites Vectors 2020, 13, 586. [Google Scholar] [CrossRef] [PubMed]
- Trejo-Huitrón, G.; Bautista-Gómez, L.G.; Martínez-Castañeda, J.S.; Romero-Núñez, C.; Trejo-Castro, L.; Espinosa-Ayala, E. Morphological characterization and first molecular identification of the eleven Eimeria species that infect sheep from Mexico. Parasitol. Res. 2020, 119, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.A.; Ferguson, D.J.; Miller, C.M.; Smith, N.C. Sex and Eimeria: A molecular perspective. Parasitology 2013, 140, 1701–1717. [Google Scholar] [CrossRef]
- Wang, R.J.; Wang, Z.Y.; Zhang, Y.J.; Su, R. Analysis on Key Factors Restricting Sustainable Development of Goat Industry. J. Domest. Anim. Ecol. 2020, 41, 77–81. (In Chinese) [Google Scholar]
- Wang, C.R.; Xiao, J.Y.; Chen, A.H.; Chen, J.; Wang, Y.; Gao, J.F.; Zhu, X.Q. Prevalence of coccidial infection in sheep and goats in northeastern China. Vet. Parasitol. 2010, 174, 213–217. [Google Scholar] [CrossRef]
- Zhao, G.H.; Lei, L.H.; Shang, C.C.; Gao, M.; Zhao, Y.Q.; Chen, C.X.; Chen, D.K. High prevalence of Eimeria infection in dairy goats in Shaanxi province, northwestern China. Trop. Anim. Health Prod. 2012, 44, 943–946. [Google Scholar] [CrossRef]
- Diao, N.C.; Zhao, B.; Chen, Y.; Wang, Q.; Chen, Z.Y.; Yang, Y.; Sun, Y.H.; Shi, J.F.; Li, J.M.; Shi, K.; et al. Prevalence of Eimeria spp. among goats in China: A systematic review and meta-analysis. Front. Cell. Infect. Microbiol. 2022, 12, 806085. [Google Scholar] [CrossRef]
- Keeton, S.T.N.; Navarre, C.B. Coccidiosis in Large and Small Ruminants. Vet. Clin. N. Am. Food Anim. Pract. 2018, 34, 201–208. [Google Scholar] [CrossRef]
- Bangoura, B.; Bardsley, K.D. Ruminant Coccidiosis. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 187–203. [Google Scholar] [CrossRef]
- Koudela, B.; Boková, A. Coccidiosis in goats in the Czech Republic. Vet. Parasitol. 1998, 76, 261–267. [Google Scholar] [CrossRef]
- Young, G.; Alley, M.L.; Foster, D.M.; Smith, G.W. Efficacy of amprolium for the treatment of pathogenic Eimeria species in Boer goat kids. Vet. Parasitol. 2011, 178, 346–349. [Google Scholar] [CrossRef] [PubMed]
- de-Macedo, L.O.; Bezerra-Santos, M.A.; de-Mendonça, C.L.; Alves, L.C.; Ramos, R.; de-Carvalho, G.A. Prevalence and risk factors associated with infection by Eimeria spp. in goats and sheep in Northeastern Brazil. J. Parasit. Dis. 2020, 44, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Joyner, L.P.; Long, P.L. The specific characters of the Eimeria, with special reference to the Coccidia of the fowl. Avian. Pathol. 1974, 3, 145–157. [Google Scholar] [CrossRef]
- Khodakaram-Tafti, A.; Hashemnia, M.; Razavi, S.M.; Sharifiyazdi, H.; Nazifi, S. Genetic characterization and phylogenetic analysis of Eimeria arloingi in Iranian native kids. Parasitol. Res. 2013, 112, 3187–3192. [Google Scholar] [CrossRef]
- Verma, R.; Sharma, D.K.; Gururaj, K.; Paul, S.; Banerjee, P.S.; Tiwari, J. Molecular epidemiology and point mutations in ITS1 and 18S rDNA genes of Eimeria ninakohlyakimovae and E. christenseni isolated from Indian goats. Vet. Parasitol. Reg. Stud. Rep. 2017, 9, 51–62. [Google Scholar] [CrossRef]
- Al-Habsi, K.; Ali, H.; Al-Kharousi, K.; Elshafie, E.I.; Al-Busaidi, R.; Muhiuddin, A.; Johnson, E.H. Vitamin B12 deficiency in newly weaned goat kids associated with clinical infection with Eimeria arloingi. Rev. Bras. Parasitol. Vet. 2020, 29, e005920. [Google Scholar] [CrossRef]
- Al-Habsi, K.; Yang, R.; Ryan, U.; Miller, D.W.; Jacobson, C. Morphological and molecular characterization of three Eimeria species from captured rangeland goats in Western Australia. Vet. Parasitol. Reg. Stud. Rep. 2017, 9, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Bawm, S.; Win, T.Z.B.; Win, S.Y.; Htun, L.L.; Nakao, R.; Katakura, K. First detection of Eimeria species in Myanmar domestic goats with both microscopic and molecular methods. Parasite 2020, 27, 38. [Google Scholar] [CrossRef]
- Ogedengbe, J.D.; Hanner, R.H.; Barta, J.R. DNA barcoding identifies Eimeria species and contributes to the phylogenetics of coccidian parasites (Eimeriorina, Apicomplexa, Alveolata). Int. J. Parasitol. 2011, 41, 843–850. [Google Scholar] [CrossRef]
- Oliveira, U.C.; Fraga, J.S.; Licois, D.; Pakandl, M.; Gruber, A. Development of molecular assays for the identification of the 11 Eimeria species of the domestic rabbit (Oryctolagus cuniculus). Vet. Parasitol. 2011, 176, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Lew, A.E.; Anderson, G.R.; Minchin, C.M.; Jeston, P.J.; Jorgensen, W.K. Inter-and intra-strain variation and PCR detection of the internal transcribed spacer 1 (ITS-1) sequences of Australian isolates of Eimeria species from chickens. Vet. Parasitol. 2003, 112, 33–50. [Google Scholar] [CrossRef]
- Hofmannová, L.; Jirků, M.; Mazánek, S.; Gremlicová, D.; Kvičerová, J. Eimeria melogale n. sp. (Apicomplexa: Eimeriidae) in the Javan ferret-badger (Melogale orientalis). Eur. J. Protistol. 2020, 73, 125668. [Google Scholar] [CrossRef] [PubMed]
- Imai, R.K.; Barta, J.R. Distribution and abundance of Eimeria species in commercial turkey flocks across Canada. Can. Vet. J. 2019, 60, 153–159. [Google Scholar]
- Yang, R.; Brice, B.; Bennett, M.D.; Eliott, A.; Ryan, U. Novel Eimeria sp. isolated from a King’s skink (Egernia kingii) in Western Australia. Exp. Parasitol. 2013, 133, 162–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eckert, J.; Braun, R.; Shirley, M.W.; Coudert, P. Guidelines on Techniques in Coccidiosis Research; OPOCE: Luxembourg, 1995; pp. 103–117. [Google Scholar]
- Schwarz, R.S.; Jenkins, M.C.; Klopp, S.; Miska, K.B. Genomic analysis of Eimeria spp. populations in relation to performance levels of broiler chicken farms in Arkansas and North Carolina. J. Parasitol. 2009, 95, 871–880. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | No. Tested | No. Positive (%) | OPG (Mean ± SD) | Species Composition (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ECH a | EAR b | ENI c | EAL d | EJO e | EHI f | ECI g | EAP h | ECO i | Others | ||||
| <1 month | 13 | 11 (84.62) | 8739 ± 18,778 | 6.11 | 40.46 | 28.24 | 5.34 | 0.00 | 16.79 | 1.53 | 1.53 | 0.00 | 0.00 |
| 1–2 months | 28 | 23 (82.14) | 7989 ± 13,430 | 41.98 | 26.72 | 8.40 | 1.53 | 0.76 | 15.27 | 3.05 | 5.34 | 1.53 | 0.76 |
| 2–3 months | 39 | 39 (100) | 162,080 ± 405,852 | 37.86 | 12.86 | 15.71 | 4.29 | 0.00 | 12.14 | 2.14 | 5.71 | 7.14 | 2.14 |
| 3–4 months | 76 | 73 (96.05) | 82,345 ± 341,874 | 17.07 | 28.46 | 17.89 | 4.88 | 1.63 | 17.89 | 2.44 | 2.44 | 6.50 | 0.81 |
| 4–5 months | 33 | 33 (100) | 86,497 ± 165,841 | 29.29 | 23.57 | 18.57 | 3.57 | 0.71 | 9.29 | 3.57 | 4.29 | 6.43 | 0.71 |
| 5–6 months | 12 | 12 (100) | 27,725 ± 26,902 | 36.43 | 10.00 | 12.14 | 2.14 | 0.71 | 5.71 | 2.14 | 5.00 | 5.00 | 2.14 |
| 6–7 months | 20 | 20 (100) | 11,290 ± 9967 | 24.29 | 23.57 | 11.43 | 4.29 | 0.00 | 3.57 | 1.43 | 2.14 | 4.29 | 0.71 |
| 7–8 months | 24 | 24 (100) | 7738 ± 6229 | 0.00 | 42.15 | 22.31 | 6.61 | 4.13 | 15.70 | 1.65 | 3.31 | 3.31 | 0.83 |
| Total | 245 | 235 (95.92) | 67,409 ± 260,945 | 24.13 | 25.97 | 16.84 | 4.08 | 0.99 | 12.05 | 2.24 | 3.72 | 4.27 | 1.01 |
| Target | Primer ID | Sequence (5′–3′) | Amplicon Size (bp) | Reference |
|---|---|---|---|---|
| 18S rDNA | ERIB1 | ACCTGGTTGATCCTGCCAG | ~1790 | [27] |
| ERIB10 | CTTCCGCAGGTTCACCTACGG | |||
| ITS1-ITS2 | ITS-1 | GGATGCAAAAGTCGTAACACGG | ~1010 | [27] |
| ITS-2 | TCCTCCGCTTAATAATATGC | |||
| COI | COI_UNI_199F | ATGATYTTCTTTGTAGTTATGCC | ~1272 | [24] |
| mtRNA20_UNI | GTATGGATTTCACGGTCAA |
| Sample Code | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | E. christenseni ON245513 | - | ||||||||
| 2 | E. arloingi ON245514 | 98.4 | - | |||||||
| 3 | E. ahsata KT184373 | 96.7 | 96.6 | - | ||||||
| 4 | E. christenseni KX857468 | 96.9 | 96.2 | 98.3 | - | |||||
| 5 | E. christenseni LC508121 | 96.9 | 96.7 | 97.8 | 98.9 | - | ||||
| 6 | E. arloingi LC508123 | 95.6 | 95.5 | 96.4 | 97.2 | 98.0 | - | |||
| 7 | E. arloingi KX857470 | 94.8 | 94.4 | 95.3 | 96.4 | 96.9 | 98.6 | - | ||
| 8 | E. hirci KX857469 | 95.3 | 94.8 | 95.8 | 97.2 | 97.3 | 98.4 | 97.7 | - | |
| 9 | E. hirci LC508122 | 96.6 | 96.4 | 96.9 | 98.0 | 98.8 | 98.4 | 97.3 | 98.4 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, G.; Yang, X.; Liu, D.; Li, Y.; Wang, J.; Chen, X.; Zhao, G.; Song, J. Molecular Characterization of 18S rDNA, ITS-1, ITS-2, and COI from Eimeria christenseni and E. arloingi in Goats from Shaanxi Province, Northwestern China. Animals 2022, 12, 1340. https://doi.org/10.3390/ani12111340
Liang G, Yang X, Liu D, Li Y, Wang J, Chen X, Zhao G, Song J. Molecular Characterization of 18S rDNA, ITS-1, ITS-2, and COI from Eimeria christenseni and E. arloingi in Goats from Shaanxi Province, Northwestern China. Animals. 2022; 12(11):1340. https://doi.org/10.3390/ani12111340
Chicago/Turabian StyleLiang, Gaoxing, Xin Yang, Ding Liu, Yuan Li, Junwei Wang, Xi Chen, Guanghui Zhao, and Junke Song. 2022. "Molecular Characterization of 18S rDNA, ITS-1, ITS-2, and COI from Eimeria christenseni and E. arloingi in Goats from Shaanxi Province, Northwestern China" Animals 12, no. 11: 1340. https://doi.org/10.3390/ani12111340