
Organ-Specific Metabolome Deciphering Cell Pathways to Cope with Mercury in Wild Fish (Golden Grey Mullet Chelon auratus)

 ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Fish Sampling
2.3. Chemicals
2.4. 1H NMR-Based Metabolomics
2.4.1. Gills and Liver Metabolite Extraction
2.4.2. 1H NMR Metabolomics and Spectral Pre-Processing
2.5. Statistical Analysis
3. Results
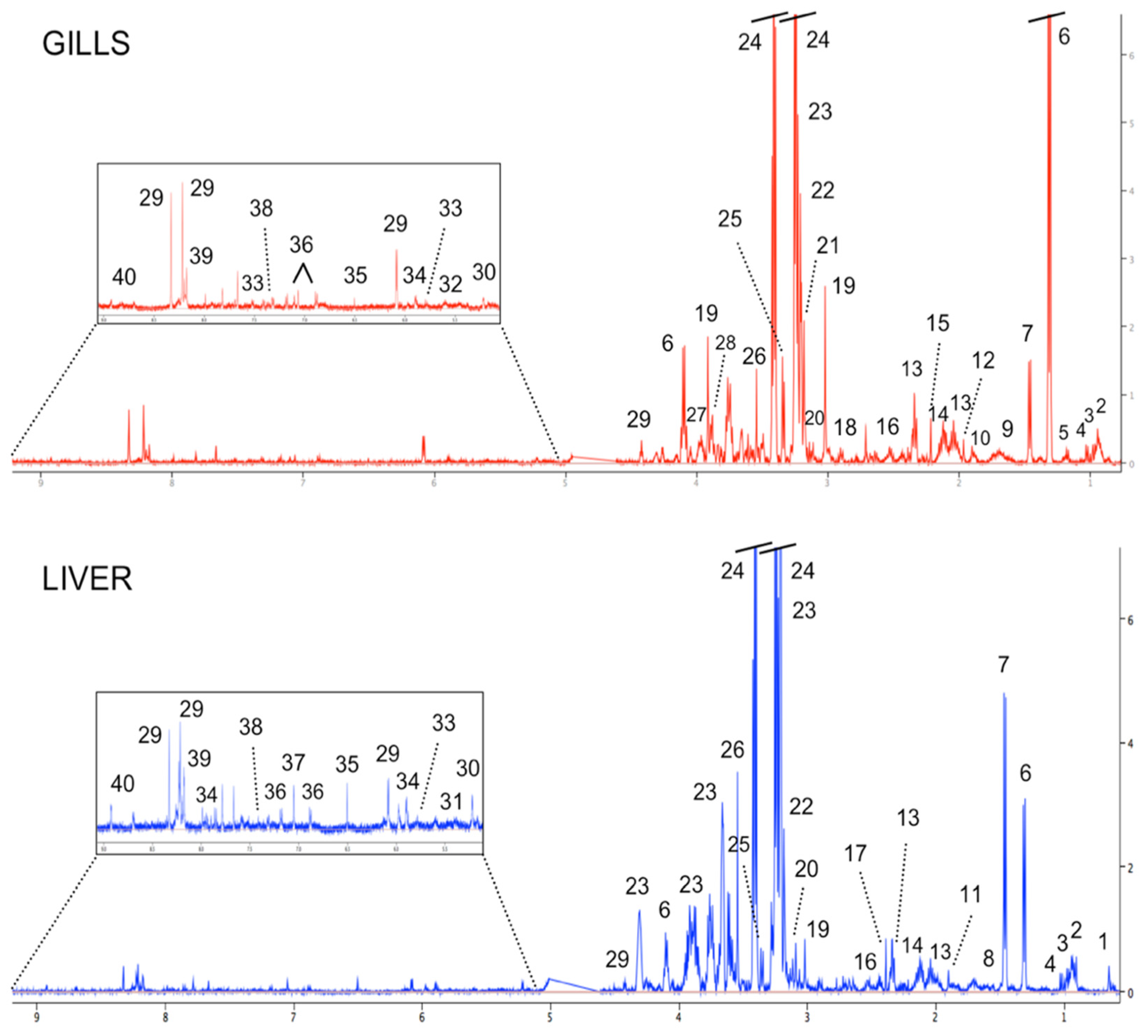
3.1. Metabolite Profiling of Fish Gills and Liver from the Reference Area
3.2. Metabolome Comparison between Fish Gills and Liver from the Reference Area
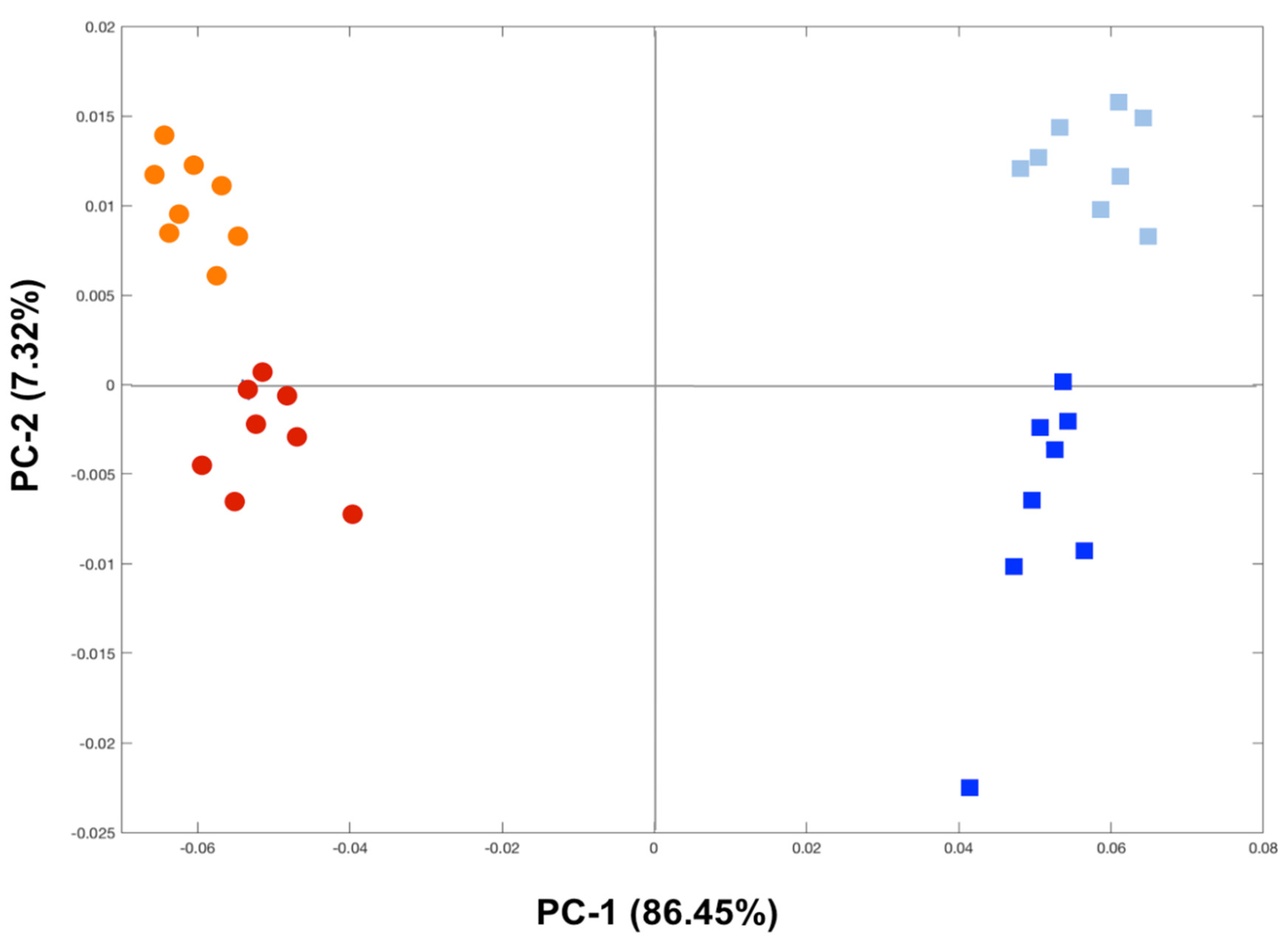
3.3. Metabolome Changes Induced by Hg in Fish Gills and Liver
4. Discussion
4.1. Metabolome of Fish Gills and Liver from the Reference Area
4.2. Tissue-Specific Metabolic Changes Induced by Hg in Fish Gills and Liver
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lankadurai, B.P.; Nagato, E.G.; Simpson, M.J. Environmental metabolomics: An emerging approach to study organism responses to environmental stressors. Environ. Ver. 2013, 21, 180–205. [Google Scholar] [CrossRef]
- Brandão, F.; Cappello, T.; Raimundo, J.; Santos, M.A.; Maisano, M.; Mauceri, A.; Pacheco, M.; Pereira, P. Unravelling the mechanisms of mercury hepatotoxicity in wild fish (Liza aurata) through a triad approach: Bioaccumulation, metabolomic profiles and oxidative stress. Metallomics 2015, 7, 1352–1363. [Google Scholar] [CrossRef] [PubMed]
- Cappello, T.; Brandao, F.; Guilherme, S.; Santos, M.A.; Maisano, M.; Mauceri, A.; Canario, J.; Pacheco, M.; Pereira, P. Insights into the mechanisms underlying mercury-induced oxidative stress in gills of wild fish (Liza aurata) combining 1H NMR metabolomics and conventional biochemical assays. Sci. Total Environ. 2016, 548–549, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Kang, J.S. Metabolomic studies for the evaluation of toxicity induced by environmental toxicants on model organisms. Metabolites 2021, 11, 485. [Google Scholar] [CrossRef] [PubMed]
- Cappello, T. NMR-based metabolomics of aquatic organisms. EMagRes 2020, 9, 81–100. [Google Scholar]
- Cappello, T.; Pereira, P.; Maisano, M.; Mauceri, A.; Pacheco, M.; Fasulo, S. Advances in understanding the mechanisms of mercury toxicity in wild golden grey mullet (Liza aurata) by 1H NMR-based metabolomics. Environ. Pollut. 2016, 219, 139–148. [Google Scholar] [CrossRef]
- Bridges, K.N.; Zhang, Y.; Curran, T.E.; Magnuson, J.T.; Venables, B.J.; Durrer, K.E.; Allen, M.S.; Roberts, A.P. Alterations to the intestinal microbiome and metabolome of Pimephales promelas and Mus musculus following exposure to dietary methylmercury. Environ. Sci. Technol. 2018, 52, 8774–8784. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, C.; Gao, X.; Zhu, J.; Wang, L.; Cao, S.; Wu, Q.; Qiao, S.; Zhang, Z.; Li, L. Comparative effects of mercury chloride and methylmercury exposure on early neurodevelopment in zebrafish larvae. RSC Adv. 2019, 9, 10766–10775. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, D.A.; Rantin, F.T.; Kalinin, A.L. Inorganic mercury exposure: Toxicological effects, oxidative stress biomarkers and bioaccumulation in the tropical freshwater fish matrinxa, Brycon amazonicus (Spix and Agassiz, 1829). Ecotoxicology 2010, 19, 105–123. [Google Scholar] [CrossRef]
- Mieiro, C.L.; Duarte, A.C.; Pereira, M.E.; Pacheco, M. Mercury accumulation patterns and biochemical endpoints in wild fish (Liza aurata): A multi-organ approach. Ecotoxicol. Environ. Saf. 2011, 74, 2225–2232. [Google Scholar] [CrossRef]
- Pereira, P.; Raimundo, J.; Barata, M.; Araújo, O.; Pousão-Ferreira, P.; Canário, J.; Almeida, A.; Pacheco, M. A new page on the road book of inorganic mercury in fish body–tissue distribution and elimination following waterborne exposure and post-exposure periods. Metallomics 2015, 7, 525–535. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.-W.; Lee, S.; Lee, B.-J.; Hur, S.-W.; Son, M.-H.; Kim, K.-W.; Kim, K.-D.; Han, H.-S. A comparative study of effects of dietary mercuric chloride and methylmercury chloride on growth performance, tissue accumulation, stress and immune responses, and plasma measurements in Korean rockfish, Sebastes schlegeli. Chemosphere 2020, 260, 127611. [Google Scholar] [CrossRef] [PubMed]
- Amiard, J.-C.; Amiard-Triquet, C.; Barka, S.; Pellerin, J.; Rainbow, P. Metallothioneins in aquatic invertebrates: Their role in metal detoxification and their use as biomarkers. Aquat. Toxicol. 2006, 76, 160–202. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, O.; Puga, S.; Brandão, F.; Canário, J.; O’Driscoll, N.J.; Santos, M.A.; Pacheco, M.; Pereira, P. Oxidative stress profiles in brain point out a higher susceptibility of fish to waterborne divalent mercury compared to dietary organic mercury. Mar. Pollut. Bull. 2017, 122, 110–121. [Google Scholar] [CrossRef] [Green Version]
- Cappello, T.; Giannetto, A.; Parrino, V.; Maisano, M.; Oliva, S.; De Marco, G.; Guerriero, G.; Mauceri, A.; Fasulo, S. Baseline levels of metabolites in different tissues of mussel Mytilus galloprovincialis (Bivalvia: Mytilidae). Comp. Biochem. Physiol. D 2018, 26, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Tsentalovich, Y.P.; Zelentsova, E.A.; Yanshole, L.V.; Yanshole, V.V.; Odud, I.M. Most abundant metabolites in tissues of freshwater fish pike-perch (Sander lucioperca). Sci. Rep. 2020, 10, 17128. [Google Scholar] [CrossRef]
- Guilherme, S.; Válega, M.; Pereira, M.; Santos, M.; Pacheco, M. Erythrocytic nuclear abnormalities in wild and caged fish (Liza aurata) along an environmental mercury contamination gradient. Ecotoxicol. Environ. Saf. 2008, 70, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Mieiro, C.; Ahmad, I.; Pereira, M.; Duarte, A.; Pacheco, M. Antioxidant system breakdown in brain of feral golden grey mullet (Liza aurata) as an effect of mercury exposure. Ecotoxicology 2010, 19, 1034–1045. [Google Scholar] [CrossRef]
- Pacheco, M.; Santos, M.A.; Teles, M.; Oliveira, M.; Lopes, C.; Rebelo, J.E. Assessment of biotransformation and genotoxicity biomarkers in feral sand-smelt (Atherina presbyter) collected in a contaminated coastal lagoon (Ria de Aveiro, Portugal). Fresenius Environ. Bull. 2004, 13, 1433–1438. [Google Scholar]
- Wu, H.; Southam, A.D.; Hines, A.; Viant, M.R. High-throughput tissue extraction protocol for NMR-and MS-based metabolomics. Anal. Biochem. 2008, 372, 204–212. [Google Scholar] [CrossRef]
- Cappello, T.; De Marco, G.; Conti, G.O.; Giannetto, A.; Ferrante, M.; Mauceri, A.; Maisano, M. Time-dependent metabolic disorders induced by short-term exposure to polystyrene microplastics in the Mediterranean mussel Mytilus galloprovincialis. Ecotoxicol. Environ. Saf. 2021, 209, 111780. [Google Scholar] [CrossRef]
- Cappello, T.; Vitale, V.; Oliva, S.; Villari, V.; Mauceri, A.; Fasulo, S.; Maisano, M. Alteration of neurotransmission and skeletogenesis in sea urchin Arbacia lixula embryos exposed to copper oxide nanoparticles. Comp. Biochem. Physiol. C 2017, 199, 20–27. [Google Scholar] [CrossRef]
- Wishart, D.S.; Knox, C.; Guo, A.C.; Eisner, R.; Young, N.; Gautam, B.; Hau, D.D.; Psychogios, N.; Dong, E.; Bouatra, S.; et al. HMDB: A knowledgebase for the human metabolome. Nucleic Acids Res. 2009, 37, D603–D610. [Google Scholar] [CrossRef]
- Pirooznia, M.; Nagarajan, V.; Deng, Y. GeneVenn—A web application for comparing gene lists using Venn diagrams. Bioinformation 2007, 1, 420–422. [Google Scholar] [CrossRef] [Green Version]
- Fasulo, S.; Maisano, M.; Sperone, E.; Mauceri, A.; Bernabò, I.; Cappello, T.; D’Agata, A.; Tripepi, S.; Brunelli, E. Toxicity of Foroozan crude oil to ornate wrasse (Thalassoma pavo, Osteichthyes, Labridae): Últrastructure and cellular biomarkers. Ital. J. Zool. 2012, 79, 182–199. [Google Scholar] [CrossRef]
- De Domenico, E.; Mauceri, A.; Giordano, D.; Maisano, M.; Giannetto, A.; Parrino, V.; Natalotto, A.; D’Agata, A.; Cappello, T.; Fasulo, S. Biological responses of juvenile European sea bass (Dicentrarchus labrax) exposed to contaminated sediments. Ecotoxicol. Environ. Saf. 2013, 97, 114–123. [Google Scholar] [CrossRef]
- Evans, D.H. The fish gill: Site of action and model for toxic effects of environmental pollutants. Environ. Health Perspect. 1987, 71, 47–58. [Google Scholar] [CrossRef]
- Maisano, M.; Cappello, T.; Oliva, S.; Natalotto, A.; Giannetto, A.; Parrino, V.; Battaglia, P.; Romeo, T.; Salvo, A.; Spanò, N.; et al. PCB and OCP accumulation and evidence of hepatic alteration in the Atlantic bluefin tuna, Thunnus thynnus, from the Mediterranean Sea. Mar. Environ. Res. 2016, 121, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Cappello, T.; Giannetto, A.; Parrino, V.; De Marco, G.; Mauceri, A.; Maisano, M. Food safety using NMR-based metabolomics: Assessment of the Atlantic bluefin tuna, Thunnus thynnus, from the Mediterranean Sea. Food Chem. Toxicol. 2018, 115, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Mieiro, C.L.; Dolbeth, M.; Marques, T.A.; Duarte, A.C.; Pereira, M.E.; Pacheco, M. Mercury accumulation and tissue-specific antioxidant efficiency in the wild European sea bass (Dicentrarchus labrax) with emphasis on seasonality. Environ. Sci. Pollut. Res. 2014, 21, 10638–10651. [Google Scholar] [CrossRef] [PubMed]
- Ung, C.Y.; Lam, S.H.; Hlaing, M.M.; Winata, C.L.; Korzh, S.; Mathavan, S.; Gong, Z. Mercury-induced hepatotoxicity in zebrafish: In vivo mechanistic insights from transcriptome analysis, phenotype anchoring and targeted gene expression validation. BMC Genom. 2010, 11, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fugelli, K.; Thoroed, S.M. Taurine and volume regulation in fish cells. Prog. Clin. Biol. Res. 1990, 351, 481–488. [Google Scholar] [PubMed]
- Avella, M.; Ducoudret, O.; Pisani, D.F.; Poujeol, P. Swelling-activated transport of taurine in cultured gill cells of sea bass: Physiological adaptation and pavement cell plasticity. Am. J. Psyhiol.-Regul. Integr. Comp. Physiol. 2009, 296, R1149–R1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, M.; Manna, P.; Sil, P.C. Induction of necrosis in cadmium-induced hepatic oxidative stress and its prevention by the prophylactic properties of taurine. J. Trace Elem. Med. Biol. 2009, 23, 300–313. [Google Scholar] [CrossRef]
- Rohlfs, E.M.; Garner, S.C.; Mar, M.H.; Zeisel, S.H. Glycerophosphocholine and phosphocholine are the major choline metabolites in rat milk. J. Nutr. 1993, 123, 1762–1768. [Google Scholar] [CrossRef]
- Calder, P.C. Branched-chain amino acids and immunity. J. Nutr. 2006, 136, 288S–293S. [Google Scholar] [CrossRef]
- Maisano, M.; Natalotto, A.; Cappello, T.; Giannetto, A.; Oliva, S.; Parrino, V.; Sanfilippo, M.; Mauceri, A. Influences of environmental variables on neurotransmission, oxidative system and hypoxia signaling on two clam species from a Mediterranean coastal lagoon. J. Shellfish Res. 2016, 35, 41–49. [Google Scholar] [CrossRef]
- Kokushi, E.; Uno, S.; Harada, T.; Koyama, J. 1H NMR-based metabolomics approach to assess toxicity of bunker a heavy oil to freshwater carp, Cyprinus carpio. Environ. Toxicol. 2012, 27, 404–414. [Google Scholar] [CrossRef]
- Postic, C.; Dentin, R.; Girard, J. Role of the liver in the control of carbohydrate and lipid homeostasis. Diabetes Metab. 2004, 30, 398–408. [Google Scholar] [CrossRef]
- Qin, X.; Wang, X. Quantification of nucleotides and their sugar conjugates in biological samples: Purposes, instruments and applications. J. Pharm. Biomed. Anal. 2018, 158, 280–287. [Google Scholar] [CrossRef]
- Elsheashaey, A.; Obada, M.; Abdelsameea, E.; Bayomy, M.F.F.; El-Said, H. The role of serum bile acid profile in differentiation between non-alcoholic fatty liver disease and chronic viral hepatitis. Egypt. Liver J. 2020, 10, 50. [Google Scholar] [CrossRef]
- Baliou, S.; Adamaki, M.; Ioannou, P.; Pappa, A.; Panayiotidis, M.I.; Spandidos, D.A.; Christodoulou, I.; Kyriakopoulos, A.M.; Zoumpourlis, V. Protective role of taurine against oxidative stress (review). Mol. Med. Rep. 2021, 24, 605. [Google Scholar] [CrossRef]
- Sonkar, K.; Ayyappan, V.; Tressler, C.M.; Adelaja, O.; Cai, R.; Cheng, M.; Glunde, K. Focus on the glycerophosphocholine pathway in choline phospholipid metabolism of cancer. NMR Biomed. 2019, 32, e4112. [Google Scholar] [CrossRef] [PubMed]
- Aquilano, K.; Baldelli, S.; Ciriolo, M.R. Glutathione: New roles in redox signalling for an old antioxidant. Front. Pharmacol. 2014, 5, 196. [Google Scholar] [CrossRef] [Green Version]
- Borchel, A.; Verleih, M.; Rebl, A.; Kühn, C.; Goldammer, T. Creatine metabolism differs between mammals and rainbow trout (Oncorhynchus mykiss). SpringerPlus 2014, 3, 510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kültz, D.; Somero, G.N. Ion transport in gills of the euryhaline fish Gillichthys mirabilis is facilitated by a phosphocreatine circuit. Am. J. Physiol. 1995, 268, R1003–R1012. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites Involved to: | Chemical Shift and Peak Shape, ppm | GILLS | LIVER |
|---|---|---|---|
| Amino Acid Metabolism | |||
| Isoleucine | 0.92 (t), 1.00 (d), 1.26 (m), 1.44 (m), 1.96 (m), 3.66 (d) | 0.33 ± 0.03 | 0.69 ± 0.15 |
| Leucine | 0.94 (d), 0.96 (d), 1.66 (m), 3.71 (t) | 0.87 ± 0.09 | 1.11 ± 0.24 |
| Valine | 0.98 (d), 1.03 (d), 2.25 (m), 3.59 (d) | 0.62 ± 0.06 | 0.92 ± 0.18 |
| Isobutyrate | 1.19 (d), 2.59 (m) | 0.51 ± 0.11 | - |
| Alanine | 1.46 (d), 3.76 (m) | 4.28 ± 0.46 | 11.25 ± 2.88 |
| Arginine | 1.68 (m), 1.90 (m), 3.23 (t), 3.74 (t) | 1.64 ± 0.22 | - |
| Lysine | 1.48 (m), 1.73 (m), 1.91 (m), 3.03 (t), 3.76 (t) | 0.58 ± 0.13 | - |
| N6-acetyllysine | 1.41 (m), 1.56 (m), 1.87 (m), 1.98 (s), 3.19 (q), 3.74 (t) | 0.46 ± 0.04 | - |
| Glutamate | 2.08 (m), 2.34 (m), 3.74 (t) | 7.61 ± 0.93 | 10.76 ± 2.23 |
| Glutamine | 2.12 (m), 2.44 (m), 3.75 (t) | 1.16 ± 0.16 | 2.17 ± 0.32 |
| Aspartate | 2.66 (dd), 2.79 (dd), 3.87 (dd) | 1.15 ± 0.17 | - |
| Creatine | 3.02 (s), 3.91 (s) | 2.16 ± 0.62 | 1.27 ± 0.13 |
| Serine | 3.84 (dd), 3.95 (m) | 2.76 ± 0.19 | - |
| Glycine | 3.54 (s) | 3.23 ± 0.19 | 6.66 ± 1.56 |
| Tyrosine | 6.89 (d), 7.19 (d) | 0.48 ± 0.08 | 0.87 ± 0.13 |
| Phenylalanine | 3.13 (m), 3.28 (m), 3.98 (m), 7.31 (d), 7.36 (t), 7.41 (m) | 0.49 ± 0.07 | 0.99 ± 0.12 |
| Energy metabolism | |||
| Acetate | 1.91 (s) | - | 0.67 ± 0.12 |
| Succinate | 2.41 (s) | - | 0.95 ± 0.16 |
| Malonate | 3.13 (s) | 0.79 ± 0.07 | 1.62 ± 0.41 |
| Glucose | 3.23 (m), 3.40 (m), 3.45 (m), 3.52 (dd), 3.73 (m), 3.82 (m), 3.88 (dd), 4.63 (d), 5.22 (d) | 2.01 ± 0.25 | 6.87 ± 1.51 |
| Glycogen | 3.40 (m), 3.60 (m), 3.80 (m), 3.96 (s), 5.40 (s) | - | 1.35 ± 0.27 |
| Lactate | 1.33 (d), 4.12 (q) | 19.84 ± 3.82 | 10.09 ± 1.68 |
| Fumarate | 6.51 (s) | 0.17 ± 0.01 | 0.54 ± 0.06 |
| UDP-glucose | 3.49 (m), 3.72 (d), 4.12 (m), 4.27 (m), 4.37 (m), 5.63 (q), 5.96 (d), 6.71 (d), 8.11 (d) | 0.36 ± 0.04 | - |
| Osmoregulation | |||
| Betaine | 3.25 (s), 3.89 (s) | 1.04 ± 0.21 | - |
| Taurine | 3.25 (s), 3.41 (t) | 45.84 ± 6.12 | 41.83 ± 11.21 |
| Glycerophosphocholine | 3.21 (s), 3.60 (dd), 3.67 (m), 3.90 (m), 4.31 (m) | 1.96 ± 0.53 | 14.54 ± 2.61 |
| Nucleotide and their derivative metabolism | |||
| Uracil | 5.81 (d), 7.54 (d) | 0.47 ± 0.05 | 1.28 ± 0.36 |
| Uridine | 3.8 (dd), 3.9 (dd), 4.1 (q), 4.2 (t), 4.3 (t), 5.9 (dd), 7.9 (d) | 0.45 ± 0.06 | 1.22 ± 0.29 |
| Hypoxanthine | 8.17 (s), 8.20 (s) | 1.07 ± 0.15 | 1.42 ± 0.25 |
| Inosine | 3.83 (dd), 3.90 (dd), 4.27 (dd), 4.25 (t), 4.76 (t), 6.08 (d), 8.21 (s), 8.33 (s) | 3.02 ± 0.27 | 1.83 ± 0.31 |
| Bile acid metabolism | |||
| Taurocholic acid | 0.65 (s), 0.91 (s), 0.98 (m), 1.36 (m), 1.42 (m), 1.61 (m), 1.96 (m), 2.13 (d), 2.20 (m), 2.53 (t), 3.18 (m), 3.28 (dd), 3.61 (s), 3.78 (s) | - | 1.59 ± 0.76 |
| Miscellaneous metabolic pathways | |||
| Choline | 3.21 (s), 3.52 (s), 4.07 (m) | 0.89 ± 0.15 | - |
| Glutathione | 2.13 (m), 2.54 (m), 2.97 (dd), 3.75 (m), 4.53 (m) | 1.17 ± 0.23 | 1.72 ± 0.22 |
| Niacinamide | 7.58 (dq), 8.24 (dt), 8.70 (dd), 8.92 (d) | 0.52 ± 0.04 | 2.33 ± 0.42 |
| Acetone | 2.22 (s) | 0.83 ± 0.22 | - |
| Phosphocholine | 3.21 (s), 3.57 (t), 4.16 (m) | 1.06 ± 0.15 | 0.95 ± 0.38 |
| Unknown resonances | |||
| Unknown resonance #1 | 1.52 (m) | - | 1.82 ± 0.68 |
| Unknown resonance #2 | 3.36 (d) | 2.88 ± 0.36 | 0.76 ± 0.11 |
| Unknown resonance #3 | 7.05 (s) | - | 2.55 ± 0.58 |
| Description | Metabolites |
|---|---|
| Both in fish gills and liver | Leucine; Isoleucine; Valine; Lactate; Alanine; Glutamate; Glutamine; Glutathione; Creatine; Malonate; Phosphocholine; Glycerophosphocholine; Taurine; Glycine; Inosine; Glucose; Uracil; Uridine; Fumarate; Tyrosine; Phenylalanine; Niacinamide; Hypoxanthine; Unknown resonance #2 |
| Only in fish gills | Acetone; Arginine; Aspartate; Betaine; Choline; Isobutyrate; Lysine; N6-acetyllysine; Serine; UDP-glucose |
| Only in fish liver | Acetate; Glycogen; Succinate; Taurocholic acid; Unknown resonance #1; Unknown resonance #3 |
| Metabolites Involved to: | GILLS | LIVER |
|---|---|---|
| Amino Acid Metabolism | ||
| Isoleucine | +34% | −15% |
| Leucine | +27% | −21% |
| Valine | +21% | −17% |
| Isobutyrate | +42% | Not found |
| Alanine | +24% a | +37% a |
| Glutamate | −22% | +24% |
| Creatine | +43% a | n.c. |
| Glycine | −26% | n.c. |
| Serine | −18% | Not found |
| Tyrosine | n.c. | −28% a |
| Phenylalanine | n.c. | −25% a |
| Energy metabolism | ||
| Lactate | +38% a | −44% |
| Fumarate | +35% | n.c. |
| Succinate | Not found | +12% |
| Glucose | n.c. | +52% a |
| Glycogen | Not found | n.c. |
| Osmoregulation | ||
| Taurine | −52% a | −38% a |
| Glycerophosphocholine | −32% a | +23% |
| Nucleoside and their derivative metabolism | ||
| Inosine | −18% | −16% |
| Uracil | +25% a | −32% |
| Hypoxanthine | n.c. | −45% a |
| Miscellaneous metabolic pathways | ||
| Glutathione | −38% a | +56% a |
| Choline | +48% a | Not found |
| Phosphocholine | −19% | +93% a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Marco, G.; Brandão, F.; Pereira, P.; Pacheco, M.; Cappello, T. Organ-Specific Metabolome Deciphering Cell Pathways to Cope with Mercury in Wild Fish (Golden Grey Mullet Chelon auratus). Animals 2022, 12, 79. https://doi.org/10.3390/ani12010079
De Marco G, Brandão F, Pereira P, Pacheco M, Cappello T. Organ-Specific Metabolome Deciphering Cell Pathways to Cope with Mercury in Wild Fish (Golden Grey Mullet Chelon auratus). Animals. 2022; 12(1):79. https://doi.org/10.3390/ani12010079
Chicago/Turabian StyleDe Marco, Giuseppe, Fátima Brandão, Patrícia Pereira, Mário Pacheco, and Tiziana Cappello. 2022. "Organ-Specific Metabolome Deciphering Cell Pathways to Cope with Mercury in Wild Fish (Golden Grey Mullet Chelon auratus)" Animals 12, no. 1: 79. https://doi.org/10.3390/ani12010079