
How Epigenetics Can Enhance Pig Welfare?

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Livestock Demands
3. Housing Systems
4. Animal Welfare Indicators
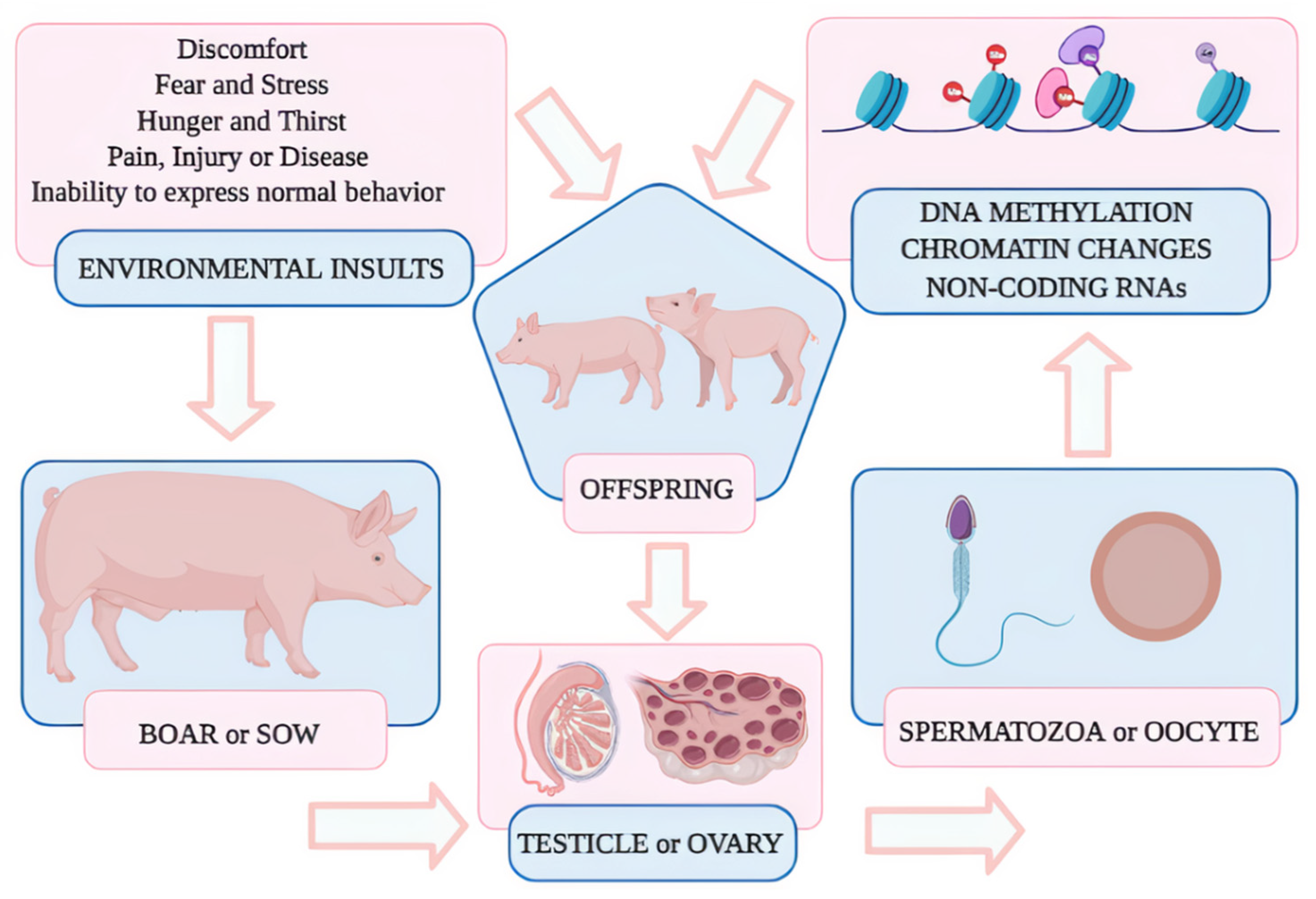
5. Epigenetics
6. Epigenetic Assessment in Mammals
7. Epigenetics Studies in the Porcine Model
8. Gene Network Analysis on Stress in Pigs
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cornish, A.; Raubenheimer, D.; McGreevy, P. What we know about the public’s level of concern for farm animal welfare in food production in developed countries. Animals 2016, 6, 74. [Google Scholar] [CrossRef] [Green Version]
- Sinclair, M.; Zhang, Y.; Descovich, K.; Phillips, C.J.C. Farm animal welfare science in China—A bibliometric review of chinese literature. Animals 2020, 10, 540. [Google Scholar] [CrossRef] [Green Version]
- de Queiroz, R.G.; de Faria Domingues, C.H.; Canozzi, M.E.A.; Garcia, R.G.; Ruviaro, C.F.; Barcellos, J.O.J.; Borges, J.A.R. How do Brazilian citizens perceive animal welfare conditions in poultry, beef, and dairy supply chains? PLoS ONE 2018, 13, e0202062. [Google Scholar] [CrossRef] [Green Version]
- Upton, M.; Otte, J. The impact of trade agreements on livestock producers. BSAP Occas. Publ. 2004, 33, 67–84. [Google Scholar] [CrossRef]
- European Commission. Study on the Impact of Animal Welfare International Activities; European Commission: Brussels, Belgium, 2017; Volume 1, pp. 212–213. [Google Scholar]
- European Community. Council directive 2001/88/ec amending directive 91/630/eec laying down minimum standards for the protection of pigs. Off. J. Eur. Community 2001, 316, 1–4. [Google Scholar] [CrossRef]
- MAPA. Estabelecer as Boas Práticas de Manejo e Bem-Estar Animal nas Granjas de Suínos de Criação Comercial; Diário Oficial da União: Brasilia, Brazil, 2020; p. 5. [Google Scholar]
- Dagg, P.J.; Butler, R.J.; Murray, J.G.; Biddle, R.R. Meeting the requirements of importing countries: Practice and policy for on-farm approaches to food safety. Rev. Sci. Tech. 2006, 25, 685–700. [Google Scholar] [CrossRef]
- Alonso, M.E.; González-Montaña, J.R.; Lomillos, J.M. Consumers’ concerns and perceptions of farm animal welfare. Animals 2020, 10, 385. [Google Scholar] [CrossRef] [Green Version]
- Trevisan, L.; Brum, J.S. Incidence of pale, soft and exudative (PSE) pork meat in reason of extrinsic stress factors. Agr. Sci. 2020, 92, e20190086. [Google Scholar] [CrossRef]
- Hyde, L.K.; Friso, S.; Choi, S.W. Introduction to Epigenetics; Elsevier Inc.: Amsterdam, The Netherlands, 2019; ISBN 9780128045725. [Google Scholar]
- Nilsson, E.; King, S.; McBirney, M.; Kubsad, D.; Pappalardo, M.; Beck, D.; Sadler-Riggleman, I.; Skinner, M.K. Vinclozolin induced epigenetic transgenerational inheritance of pathologies and sperm epimutation biomarkers for specific diseases. PLoS ONE 2018, 13, e0202662. [Google Scholar] [CrossRef]
- Lyko, F.; Foret, S.; Kucharski, R.; Wolf, S.; Falckenhayn, C.; Maleszka, R. The honey bee epigenomes: Differential methylation of brain DNA in queens and workers. PLoS Biol. 2010, 8, e1000506. [Google Scholar] [CrossRef] [Green Version]
- Pértille, F.; Ibelli, A.M.G.; El Sharif, M.; Poleti, M.D.; Fröhlich, A.S.; Rezaei, S.; Ledur, M.C.; Jensen, P.; Guerrero-Bosagna, C.; Coutinho, L.L. Putative Epigenetic Biomarkers of Stress in Red Blood Cells of Chickens Reared Across Different Biomes. Front. Genet. 2020, 11, 508809. [Google Scholar] [CrossRef]
- Hao, Y.; Cui, Y.; Gu, X. Genome-wide DNA methylation profiles changes associated with constant heat stress in pigs as measured by bisulfite sequencing. Sci. Rep. 2016, 6, 27507. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.; Liu, J.R.; Zhang, Y.; Yang, P.G.; Feng, Y.J.; Cui, Y.J.; Yang, C.H.; Gu, X.H. The microRNA expression profile in porcine skeletal muscle is changed by constant heat stress. Anim. Genet. 2016, 47, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Pértille, F.; Brantsæter, M.; Nordgreen, J.; Coutinho, L.L.; Janczak, A.M.; Jensen, P.; Guerrero-Bosagna, C. DNA methylation profiles in red blood cells of adult hens correlate with their rearing conditions. J. Exp. Biol. 2017, 220, 3579–3587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turk, J. Meeting projected food demands by 2050: Understanding and enhancing the role of grazing ruminants. J. Anim. Sci. 2016, 94, 53–62. [Google Scholar] [CrossRef]
- Kearney, J. Food consumption trends and drivers. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2793–2807. [Google Scholar] [CrossRef]
- United Nations. Transforming our World: The 2030 Agenda for Sustainable Development; New York, NY, USA, 2015. [Google Scholar]
- Keeling, L.; Tunón, H.; Olmos Antillón, G.; Berg, C.; Jones, M.; Stuardo, L.; Swanson, J.; Wallenbeck, A.; Winckler, C.; Blokhuis, H. Animal welfare and the United Nations sustainable development goals. Front. Vet. Sci. 2019, 6, 336. [Google Scholar] [CrossRef]
- Savulescu, J.; Vergunst, F. Five Ways the Meat on Your Plate Is Killing the Planet. 2017. Available online: https://theconversation.com/five-ways-the-meat-on-your-plate-is-killing-the-planet-76128 (accessed on 3 August 2021).
- Blokhuis, H.J.; Keeling, L.J.; Gavinelli, A.; Serratosa, J. Animal welfare’s impact on the food chain. Trends Food Sci. Technol. 2008, 19, S79–S87. [Google Scholar] [CrossRef]
- Buller, H.; Blokhuis, H.; Jensen, P.; Keeling, L. Towards farm animal welfare and sustainability. Animals 2018, 8, 81. [Google Scholar] [CrossRef] [Green Version]
- National Research Council. Guide for the care and use of laboratory animals; The National Academies Press: Washington, DC, USA, 1996; pp. 25–26. ISBN 9780309053778. [Google Scholar]
- Region, W.; Tracy, K. The effect of housing systems on the welfare of pigs in santa sub-division in the North West region of Cameroon. J. Environ. Issues Agric. Dev. Ctries. 2018, 10, 24–25. [Google Scholar]
- Li, Y.Z.; Gonyou, H.W. Comparison of management options for sows kept in pens with electronic feeding stations. Can. J. Anim. Sci. 2013, 93, 445–452. [Google Scholar] [CrossRef]
- McGlone, J.J. The future of pork production in the world: Towards sustainable, welfare-positive systems. Animals 2013, 3, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Jansen, J.; Kirkwood, R.N.; Zanella, A.J.; Tempelman, R.J. Influence of gestation housing on sow behavior and fertility. J. Swine Health Prod. 2007, 15, 132–136. [Google Scholar]
- Jang, J.C.; Hong, J.S.; Jin, S.S.; Kim, Y.Y. Comparing gestating sows housing between electronic sow feeding system and a conventional stall over three consecutive parities. Livest. Sci. 2017, 199, 37–45. [Google Scholar] [CrossRef]
- Van Epps, A.; Blaney, L. Antibiotic residues in animal waste: Occurrence and degradation in conventional agricultural waste management practices. Curr. Pollut. Rep. 2016, 2, 135–155. [Google Scholar] [CrossRef] [Green Version]
- Dourmad, J.Y.; Ryschawy, J.; Trousson, T.; Bonneau, M.; Gonzàlez, J.; Houwers, H.W.J.; Hviid, M.; Zimmer, C.; Nguyen, T.L.T.; Morgensen, L. Evaluating environmental impacts of contrasting pig farming systems with life cycle assessment. Animal 2014, 8, 2027–2037. [Google Scholar] [CrossRef] [Green Version]
- Harrison, R. Animal Machine; Vincent Stuart: London, UK, 1982; Volume 296, ISBN 9781780642840. [Google Scholar]
- The Humane Society of the United States. Welfare issues with gestation crates for pregnant sows. Impacts Farm Anim. 2013, 25, 3. [Google Scholar]
- Mellor, D.J. Updating animal welfare thinking: Moving beyond the “five freedoms” towards “A lifeworth living”. Animals 2016, 6, 21. [Google Scholar] [CrossRef]
- Browning, H.; Veit, W. Is humane slaughter possible? Animals 2020, 10, 799. [Google Scholar] [CrossRef] [PubMed]
- Parada Sarmiento, M.; Bernardino, T.; Tatemoto, P.; Polo, G.; Zanella, A.J. The in-utero experience of piglets born from sows with lameness shapes their life trajectory. Sci. Rep. 2021, 11, 13052. [Google Scholar] [CrossRef]
- Eastwood, L. Group Housing Systems for Dry Sows; Compassion in World Farming: Surrey, UK, 2017; Volume 17–37, pp. 1–4. [Google Scholar]
- Broom, D.M. Animal welfare: Concepts and measurement. J. Anim. Sci. 1991, 69, 4167–4175. [Google Scholar] [CrossRef] [PubMed]
- Poletto, R.; Hötzel, M.J. The five freedoms in the global animal agriculture market: Challenges and achievements as opportunities. Anim. Front. 2012, 2, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Broom, D.M. Indicators of poor welfare. Br. Vet. J. 1986, 142, 524–526. [Google Scholar] [CrossRef]
- Mouzo, D.; Rodríguez-Vázquez, R.; Lorenzo, J.M.; Franco, D.; Zapata, C.; López-Pedrouso, M. Proteomic application in predicting food quality relating to animal welfare. A review. Trends Food Sci. Technol. 2020, 99, 520–530. [Google Scholar] [CrossRef]
- Hunter, R.R.; Mitchell, M.A.; Carlisle, A.J.; Quinn, A.D.; Kettlewell, P.J.; Knowles, T.G.; Warriss, P.D. Physiological responses of broilers to pre-slaughter lairage: Effects of the thermal micro-environment? Br. Poult. Sci. 1998, 39, 53–54. [Google Scholar] [CrossRef] [PubMed]
- D’eath, R.B.; Turner, S.P.; Kurt, E.; Evans, G.; Thölking, L.; Looft, H.; Wimmers, K.; Murani, E.; Klont, R.; Foury, A.; et al. Pigs’ aggressive temperament affects pre-slaughter mixing aggression, stress and meat quality. Animal 2010, 4, 604–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fidan, E.D.; Türkyılmaz, M.K.; Nazlıgül, A.; Aypak, S.Ü.; Karaarslan, S. Effect of preslaughter shackling on stress, meat quality traits, and glycolytic potential in broilers. J. Agric. Sci. Technol. 2015, 17, 1141–1150. [Google Scholar]
- Loudon, K.M.W.; Tarr, G.; Lean, I.J.; Polkinghorne, R.; McGilchrist, P.; Dunshea, F.R.; Gardner, G.E.; Pethick, D.W. The impact of pre-slaughter stress on beef eating quality. Animals 2019, 9, 612. [Google Scholar] [CrossRef] [Green Version]
- Rocha, L.M.; Dionne, A.; Saucier, L.; Nannoni, E.; Faucitano, L. Hand-held lactate analyzer as a tool for the real-time measurement of physical fatigue before slaughter and pork quality prediction. Animal 2015, 24, 707–714. [Google Scholar] [CrossRef] [Green Version]
- Möstl, E.; Palme, R. Hormones as indicators of stress. Domest. Anim. Endocrinol. 2002, 23, 67–74. [Google Scholar] [CrossRef]
- Moberg, G.P. Influence of stress on reproduction: Measure on well-being. In Animal Stress; Moberg, G.P., Ed.; Springer: New York, NY, USA, 1985; Volume 53, pp. 245–267. ISBN 9788578110796. [Google Scholar]
- Einarsson, S.; Madej, A.; Tsuma, V. The influence of stress on early pregnancy in the pig. Anim. Reprod. Sci. 1996, 42, 165–172. [Google Scholar] [CrossRef]
- Berger, S.; Wikelski, M.; Romero, L.M.; Kalko, E.K.V.; Rödl, T. Behavioral and physiological adjustments to new predators in an endemic island species, the Galápagos marine iguana. Horm. Behav. 2007, 52, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Palme, R. Non-invasive measurement of glucocorticoids: Advances and problems. Physiol. Behav. 2019, 199, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Dickens, M.J.; Romero, L.M. A consensus endocrine profile for chronically stressed wild animals does not exist. Gen. Comp. Endocrinol. 2013, 191, 177–189. [Google Scholar] [CrossRef]
- Lee, D.Y.; Kim, E.; Choi, M.H. Technical and clinical aspects of cortisol as a biochemical marker of chronic stress. BMB Rep. 2015, 48, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Pollard, T.M. Use of cortisol as a stress marker: Practical and theoretical problems. Am. J. Hum. Biol. 1995, 7, 265–274. [Google Scholar] [CrossRef]
- Hunter, R.G. Epigenetic effects of stress and corticosteroids in the brain. Front. Cell. Neurosci. 2012, 6, 18. [Google Scholar] [CrossRef] [Green Version]
- Nätt, D.; Johansson, I.; Faresjö, T.; Ludvigsson, J.; Thorsell, A. High cortisol in 5-year-old children causes loss of DNA methylation in SINE retrotransposons: A possible role for ZNF263 in stress-related diseases. Clin. Epigenet. 2015, 7, 91. [Google Scholar] [CrossRef] [Green Version]
- Laplant, Q.; Vialou, V.; Covington, H.E.; Dumitriu, D.; Feng, J.; Warren, B.L.; Maze, I.; Dietz, D.M.; Watts, E.L.; Iñiguez, S.D.; et al. Dnmt3a regulates emotional behavior and spine plasticity in the nucleus accumbens. Nat. Neurosci. 2010, 13, 1137–1143. [Google Scholar] [CrossRef] [Green Version]
- Sailaja, B.S.; Cohen-Carmon, D.; Zimmerman, G.; Soreq, H.; Meshorer, E. Stress-induced epigenetic transcriptional memory of acetylcholinesterase by HDAC4. Proc. Natl. Acad. Sci. USA 2012, 109, E3687–E3695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasper, C.; Ribeiro, D.; de Almeida, A.M.; Larzul, C.; Liaubet, L.; Murani, E. Omics application in animal science—A special emphasis on stress response and damaging behaviour in pigs. Genes 2020, 11, 920. [Google Scholar] [CrossRef] [PubMed]
- Waddington, C.H. The pupal contraction as an epigenetic crisis in drosophila. J. Zool. 1942, A111, 188. [Google Scholar]
- Gibney, E.R.; Nolan, C.M. Epigenetics and gene expression. Heredity 2010, 105, 4–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lind, M.I.; Spagopoulou, F. Evolutionary consequences of epigenetic inheritance. Heredity 2018, 121, 205–209. [Google Scholar] [CrossRef] [Green Version]
- Sundman, A.S.; Pértille, F.; Coutinho, L.L.; Jazin, E.; Guerrero-Bosagna, C.; Jensen, P. DNA methylation in canine brains is related to domestication and dog-breed formation. PLoS ONE 2020, 15, e0240787. [Google Scholar] [CrossRef]
- Pértille, F.; Da Silva, V.H.; Johansson, A.M.; Lindström, T.; Wright, D.; Coutinho, L.L.; Jensen, P.; Guerrero-Bosagna, C. Mutation dynamics of CpG dinucleotides during a recent event of vertebrate diversification. Epigenetics 2019, 14, 685–707. [Google Scholar] [CrossRef] [Green Version]
- Zhou, N.; Cao, Z.; Wu, R.; Liu, X.; Tao, J.; Chen, Z.; Song, D.; Han, F.; Li, Y.; Fang, F.; et al. Dynamic changes of histone H3 lysine 27 acetylation in pre-implantational pig embryos derived from somatic cell nuclear transfer. Anim. Reprod. Sci. 2014, 148, 153–163. [Google Scholar] [CrossRef]
- Han, K.; Ren, R.; Cao, J.; Zhao, S.; Yu, M. Genome-wide identification of histone modifications involved in placental development in pigs. Front. Genet. 2019, 10, 277. [Google Scholar] [CrossRef] [Green Version]
- Morgan, A.; Legresley, S.; Fischer, C. Remodeler catalyzed nucleosome repositioning: Influence of structure and stability. Int. J. Mol. Sci. 2021, 22, 76. [Google Scholar] [CrossRef]
- Greenberg, M.V.C.; Bourc’his, D. The diverse roles of DNA methylation in mammalian development and disease. Nat. Rev. Mol. Cell Biol. 2019, 20, 590–607. [Google Scholar] [CrossRef] [PubMed]
- Lienhard, M.; Grimm, C.; Morkel, M.; Herwig, R.; Chavez, L. MEDIPS: Genome-wide differential coverage analysis of sequencing data derived from DNA enrichment experiments. Bioinformatics 2014, 30, 284–286. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, K.D.; Rutherford, K.M.D.; Wallace, J.M.; Brameld, J.M.; Stöger, R.; Alberio, R.; Sweetman, D.; Gardner, D.S.; Perry, V.E.A.; Adam, C.L.; et al. Epigenetics and developmental programming of welfare and production traits in farm animals. Reprod. Fertil. Dev. 2016, 28, 1443–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutherford, K.M.D.; Piastowska-Ciesielska, A.; Donald, R.D.; Robson, S.K.; Ison, S.H.; Jarvis, S.; Brunton, P.J.; Russell, J.A.; Lawrence, A.B. Prenatal stress produces anxiety prone female offspring and impaired maternal behaviour in the domestic pig. Physiol. Behav. 2014, 129, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Otten, W.; Kanitz, E.; Tuchscherer, M. The impact of pre-natal stress on offspring development in pigs. J. Agric. Sci. 2015, 153, 907–919. [Google Scholar] [CrossRef]
- Bernardino, T.; Tatemoto, P.; Morrone, B.; Rodrigues, P.H.M.; Zanella, A.J. Piglets born from sows fed high fibre diets during pregnancy are less aggressive prior to weaning. PLoS ONE 2016, 11, e0167363. [Google Scholar] [CrossRef] [Green Version]
- Tatemoto, P.; Bernardino, T.; Alves, L.; Zanella, A.J. Sham-chewing in sows is associated with decreased fear responses in their offspring. Front. Vet. Sci. 2019, 6, 390. [Google Scholar] [CrossRef]
- Tatemoto, P.; Bernardino, T.; Alves, L.; Cristina de Oliveira Souza, A.; Palme, R.; José Zanella, A. Environmental enrichment for pregnant sows modulates HPA-axis and behavior in the offspring. Appl. Anim. Behav. Sci. 2019, 220, 104854. [Google Scholar] [CrossRef]
- Sullivan, R.; Saez, F. Epididymosomes, prostasomes, and liposomes: Their roles in mammalian male reproductive physiology. Reproduction 2013, 146, R21–R35. [Google Scholar] [CrossRef] [Green Version]
- Weaver, I.C.G.; Cervoni, N.; Champagne, F.A.; D’Alessio, A.C.; Sharma, S.; Seckl, J.R.; Dymov, S.; Szyf, M.; Meaney, M.J. Epigenetic programming by maternal behavior. Nat. Neurosci. 2004, 7, 847–854. [Google Scholar] [CrossRef]
- Champagne, F.A. Epigenetic mechanisms and the transgenerational effects of maternal care. Front. Neuroendocrinol. 2008, 29, 386–397. [Google Scholar] [CrossRef] [Green Version]
- Lacal, I.; Ventura, R. Epigenetic inheritance: Concepts, mechanisms and perspectives. Front. Mol. Neurosci. 2018, 11, 292. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.; Canning, J.; Kaneko, T.; Pru, J.K.; Tilly, J.L. Germline stem cells and follicular renewal in the postnatal mammalian ovary. Nature 2004, 430, 1062. [Google Scholar] [CrossRef]
- Johnson, J.; Bagley, J.; Skaznik-Wikiel, M.; Lee, H.J.; Adams, G.B.; Niikura, Y.; Tschudy, K.S.; Tilly, J.C.; Cortes, M.L.; Forkert, R.; et al. Oocyte generation in adult mammalian ovaries by putative germ cells in bone marrow and peripheral blood. Cell 2005, 122, 303–315. [Google Scholar] [CrossRef] [Green Version]
- Kyriazakis, I.; Whittemore, C.T. Development of the reproductive system during intra-uterine life. In Whittemore’s Science and Practice of Pig Production; Wiley-Blackwell: Hoboken, NJ, USA, 2006; p. 105. ISBN 9781405124485. [Google Scholar]
- Skvortsova, K.; Iovino, N.; Bogdanović, O. Functions and mechanisms of epigenetic inheritance in animals. Nat. Rev. Mol. Cell Biol. 2018, 19, 774–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuscher, J.J.; Day, J.J. Multigenerational epigenetic inheritance: One step forward, two generations back. Neurobiol. Dis. 2019, 132, 104591. [Google Scholar] [CrossRef]
- Guerrero-Bosagna, C.; Settles, M.; Lucker, B.; Skinner, M.K. Epigenetic transgenerational actions of vinclozolin on promoter regions of the sperm epigenome. PLoS ONE 2010, 5, e13100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manikkam, M.; Guerrero-Bosagna, C.; Tracey, R.; Haque, M.M.; Skinner, M.K. Transgenerational actions of environmental compounds on reproductive disease and identification of epigenetic biomarkers of ancestral exposures. PLoS ONE 2012, 7, e31901. [Google Scholar] [CrossRef] [Green Version]
- Pistek, V.L.; Fürst, R.W.; Kliem, H.; Bauersachs, S.; Meyer, H.H.D.; Ulbrich, S.E. HOXA10 mRNA expression and promoter DNA methylation in female pig offspring after in utero estradiol-17β exposure. J. Steroid Biochem. Mol. Biol. 2013, 138, 435–444. [Google Scholar] [CrossRef]
- Posner, R.; Toker, I.A.; Antonova, O.; Star, E.; Anava, S.; Azmon, E.; Hendricks, M.; Bracha, S.; Gingold, H.; Rechavi, O. Neuronal small RNAs control behavior transgenerationally. Cell 2019, 177, 1814–1826.e15. [Google Scholar] [CrossRef] [Green Version]
- Tarleton, B.J.; Wiley, A.A.; Bartol, F.F. Neonatal estradiol exposure alters uterine morphology and endometrial transcriptional activity in prepubertal gilts. Domest. Anim. Endocrinol. 2001, 21, 111–125. [Google Scholar] [CrossRef]
- Niculescu, M.D.; Zeisel, S.H. Diet, methyl donors and DNA methylation: Interactions between dietary folate, methionine and choline. J. Nutr. 2002, 132, 2333–2335. [Google Scholar] [CrossRef]
- Liu, X.; Wang, J.; Li, R.; Yang, X.; Sun, Q.; Albrecht, E.; Zhao, R. Maternal dietary protein affects transcriptional regulation of myostatin gene distinctively at weaning and finishing stages in skeletal muscle of Meishan pigs. Epigenetics 2011, 6, 899–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altmann, S.; Murani, E.; Schwerin, M.; Metges, C.C.; Wimmers, K.; Ponsuksili, S. Maternal dietary protein restriction and excess affects offspring gene expression and methylation of non-SMC subunits of condensin i in liver and skeletal muscle. Epigenetics 2012, 7, 239–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braunschweig, M.; Jagannathan, V.; Gutzwiller, A.; Bee, G. Investigations on transgenerational epigenetic response down the male line in F2 pigs. PLoS ONE 2012, 7, e30583. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Hu, L.; Gong, D.; Lu, H.; Xuan, Y.; Wang, R.; Wu, D.; Chen, D.; Zhang, K.; Gao, F.; et al. Genome-wide DNA methylation analysis in jejunum of Sus scrofa with intrauterine growth restriction. Mol. Genet. Genom. 2018, 293, 807–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponsuksili, S.; Trakooljul, N.; Basavaraj, S.; Hadlich, F.; Murani, E.; Wimmers, K. Epigenome-wide skeletal muscle DNA methylation profiles at the background of distinct metabolic types and ryanodine receptor variation in pigs. BMC Genom. 2019, 20, 492. [Google Scholar] [CrossRef]
- Wang, M.; Ibeagha-Awemu, E.M. Impacts of epigenetic processes on the health and productivity of livestock. Front. Genet. 2021, 11, 613636. [Google Scholar] [CrossRef]
- Yu, X.X.; Liu, Y.H.; Liu, X.M.; Wang, P.C.; Liu, S.; Miao, J.K.; Du, Z.Q.; Yang, C.X. Ascorbic acid induces global epigenetic reprogramming to promote meiotic maturation and developmental competence of porcine oocytes. Sci. Rep. 2018, 8, 6132. [Google Scholar] [CrossRef]
- Taweechaipaisankul, A.; Jin, J.; Lee, S.; Kim, G.; Suh, Y.; Ahn, M.; Park, S.; Lee, B.; Lee, B. Improved early development of porcine cloned embryos by treatment with quisinostat, a potent histone deacetylase inhibitor. J. Reprod. Dev. 2019, 65, 2–6. [Google Scholar] [CrossRef] [Green Version]
- Rempel, L.A.; Parrish, J.J.; Miles, J.R. Genes associated with chromatin modification within the swine placenta are differentially expressed due to factors associated with season. Front. Genet. 2020, 11, 1019. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Zhang, J.; Jiang, P.; Gong, D.; Wang, J.W.; Xia, Y.; Østergaard, M.V.; Wang, J.; Sangild, P.T. Marked methylation changes in intestinal genes during the perinatal period of preterm neonates. BMC Genom. 2014, 15, 716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khezri, A.; Narud, B.; Stenseth, E.B.; Johannisson, A.; Myromslien, F.D.; Gaustad, A.H.; Wilson, R.C.; Lyle, R.; Morrell, J.M.; Kommisrud, E.; et al. DNA methylation patterns vary in boar sperm cells with different levels of DNA fragmentation. BMC Genom. 2019, 20, 897. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Kadarmideen, H.N. Characterization of global DNA methylation in different gene regions reveals candidate biomarkers in pigs with high and low levels of boar taint. Vet. Sci. 2020, 7, 77. [Google Scholar] [CrossRef]
- Pértille, F.; Alvarez-rodriguez, M.; da Silva, A.N.; Barranco, I.; Roca, J.; Guerrero-bosagna, C.; Rodriguez-martinez, H. Sperm methylome profiling can discern fertility levels in the porcine biomedical model. Int. J. Mol. Sci. 2021, 22, 2679. [Google Scholar] [CrossRef]
- Tatemoto, P.; Bernardino, T.; Morrone, B.; Queiroz, M.R. Stereotypic behavior in sows is related to emotionality changes in the offspring. Front. Vet. Sci. 2020, 7, 79. [Google Scholar] [CrossRef] [Green Version]
- Kuzmuk, K.N.; Schook, L.B. Pigs as a model for biomedical sciences. In The Genetics of the Pig; Rothschild, M.F., Ruvinsky, A., Eds.; CABI: Urbana, IL, USA, 2011; pp. 426–444. ISBN 9781845937560. [Google Scholar]
- Martínez-Miró, S.; Tecles, F.; Ramón, M.; Escribano, D.; Hernández, F.; Madrid, J.; Orengo, J.; Martínez-Subiela, S.; Manteca, X.; Cerón, J.J. Causes, consequences and biomarkers of stress in swine: An update. BMC Vet. Res. 2016, 12, 171. [Google Scholar] [CrossRef] [Green Version]
- Fagerberg, L.; Hallstrom, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Mol. Cell. Proteom. 2014, 13, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Lunney, J.K.; Van Goor, A.; Walker, K.E.; Hailstock, T.; Franklin, J.; Dai, C. Importance of the pig as a human biomedical model. Science 2021, 13, 5758. [Google Scholar] [CrossRef]
- Collier, C.T.; Williams, P.N.; Carroll, J.A.; Welsh, T.H.; Laurenz, J.C. Effect of maternal restraint stress during gestation on temporal lipopolysaccharide-induced neuroendocrine and immune responses of progeny. Domest. Anim. Endocrinol. 2011, 40, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Schachtschneider, K.M.; Welge, M.E.; Auvil, L.S.; Chaki, S.; Rund, L.A.; Madsen, O.; Elmore, M.R.P.; Johnson, R.W.; Groenen, M.A.M.; Schook, L.B. Altered hippocampal epigenetic regulation underlying reduced cognitive development in response to early life environmental insults. Genes 2020, 11, 162. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.L.; Zhang, Z.; Li, B.; Gao, N.; Zhang, H.; Sangild, P.T.; Li, J.Q. Genome-wide DNA methylation analysis of the porcine hypothalamus-pituitary-ovary axis. Sci. Rep. 2017, 7, 4277. [Google Scholar] [CrossRef] [Green Version]
- Yuan, X.; Zhou, X.; Chen, Z.; He, Y.; Kong, Y.; Ye, S.; Gao, N.; Zhang, Z.; Zhang, H.; Li, J. Genome-wide DNA methylation analysis of hypothalamus during the onset of puberty in gilts. Front. Genet. 2019, 10, 1–10. [Google Scholar] [CrossRef]
- Witek, P.; Grzesiak, M.; Kotula-Balak, M.; Koziorowski, M.; Slomczynska, M.; Knapczyk-Stwora, K. Effect of neonatal exposure to endocrine-active compounds on epigenetic regulation of gene expression in corpus luteum of gilts. Theriogenology 2020, 159, 45–52. [Google Scholar] [CrossRef]
- Yuan, X.; Ye, S.; Chen, Z.; Pan, X.; Huang, S.; Li, Z.; Zhong, Y.; Gao, N.; Zhang, H.; Li, J.; et al. Dynamic DNA methylation of ovaries during pubertal transition in gilts. BMC Genom. 2019, 20, 228. [Google Scholar] [CrossRef]
- Sajjanar, B.; Trakooljul, N.; Wimmers, K.; Ponsuksili, S. DNA methylation analysis of porcine mammary epithelial cells reveals differentially methylated loci associated with immune response against Escherichia coli challenge. BMC Genom. 2019, 20, 623. [Google Scholar] [CrossRef]
- Simões, M.; Rino, J.; Pinheiro, I.; Martins, C.; Ferreira, F. Alterations of nuclear architecture and epigenetic signatures during African swine fever virus infection. Viruses 2015, 7, 4978–4996. [Google Scholar] [CrossRef] [PubMed]
- Knipe, D.M.; Lieberman, P.M.; Jung, J.U.; McBride, A.A.; Morris, K.V.; Ott, M.; Margolis, D.; Nieto, A.; Nevels, M.; Parks, R.J.; et al. Snapshots: Chromatin control of viral infection. Virology 2013, 435, 141–156. [Google Scholar] [CrossRef]
- John, R.M.; Rougeulle, C. Developmental epigenetics: Phenotype and the flexible epigenome. Front. Cell Dev. Biol. 2018, 6, 130. [Google Scholar] [CrossRef] [PubMed]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Iwayama, Y.; Toyota, T.; Ohnishi, T.; Ohba, H.; Maekawa, M.; Yoshikawa, T. Association study of the KCNJ3 gene as a susceptibility candidate for schizophrenia in the Chinese population. Hum. Genet. 2012, 131, 443–451. [Google Scholar] [CrossRef] [Green Version]
- Zarrei, M.; Fehlings, D.L.; Mawjee, K.; Switzer, L.; Thiruvahindrapuram, B.; Walker, S.; Merico, D.; Casallo, G.; Uddin, M.; MacDonald, J.R.; et al. De novo and rare inherited copy-number variations in the hemiplegic form of cerebral palsy. Genet. Med. 2018, 20, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Capera, J.; Serrano-Novillo, C.; Navarro-Pérez, M.; Cassinelli, S.; Felipe, A. The potassium channel odyssey: Mechanisms of traffic and membrane arrangement. Int. J. Mol. Sci. 2019, 20, 734. [Google Scholar] [CrossRef] [Green Version]
- Tyson, J.R.; Snutch, T.P. Molecular nature of voltage-gated calcium channels: Structure and species comparison. Wiley Interdiscip. Rev. Membr. Transp. Signal. 2013, 2, 181–206. [Google Scholar] [CrossRef]
- Svoboda, L.K.; Reddie, K.G.; Zhang, L.; Vesely, E.D.; Williams, E.S.; Schumacher, S.M.; O’Connell, R.P.; Shaw, R.; Day, S.M.; Anumonwo, J.M.; et al. Redox-sensitive sulfenic acid modification regulates surface expression of the cardiovascular voltage-gated potassium channel Kv1.5. Circ. Res. 2012, 111, 842–853. [Google Scholar] [CrossRef] [Green Version]
- Jeevaratnam, K.; Chadda, K.R.; Huang, C.L.H.; Camm, A.J. Cardiac potassium channels: Physiological insights for targeted therapy. J. Cardiovasc. Pharmacol. Ther. 2018, 23, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashcroft, F.M.; Rorsman, P. KATP channels and islet hormone secretion: New insights and controversies. Nat. Rev. Endocrinol. 2013, 9, 660–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comes, N.; Serrano-Albarrás, A.; Capera, J.; Serrano-Novillo, C.; Condom, E.; Ramón, Y.; Cajal, S.; Ferreres, J.C.; Felipe, A. Involvement of potassium channels in the progression of cancer to a more malignant phenotype. Biochim. Biophys. Acta Biomembr. 2015, 1848, 2477–2492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabò, I.; Zoratti, M.; Gulbins, E. Contribution of voltage-gated potassium channels to the regulation of apoptosis. FEBS Lett. 2010, 584, 2049–2056. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.; Zhang, Q.; Zhang, Y.; Wang, F.; Wang, L.; Yamamoto, R.; Sugai, T.; Kato, N. Fear conditioning suppresses large-conductance calcium-activated potassium channels in lateral amygdala neurons. Physiol. Behav. 2015, 138, 279–284. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Liu, S.B.; Cui, G.-B.; Ma, L.; Feng, B.; Xing, J.H.; Yang, Q.; Li, X.Q.; Wu, Y.; Xiong, L.Z.; et al. Acute stress induces down-regulation of large-conductance Ca 2+-activated potassium channels in the lateral amygdala. J. Physiol. 2012, 590, 875–886. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Mechanism | Generation | Swine Model | Context | Reference |
|---|---|---|---|---|
| Direct exposure | F0 | Boars | Investigation of methylation patterns of testis samples and their relationship with the boar taint flavour. | [104] |
| Boar’s semen | Correlation between different parameters of sperm DNA integrity and their methylation patterns. | [103] | ||
| DMRs are more efficient at discerning the fertility of boars’ ejaculate than single nucleotide polymorphisms (SNPs) using reduced representation of the methylated DNA. | [105] | |||
| Gilts | The epigenetic dynamic in hypothalamus-pituitary-ovary axis and its tissue-specific manner to establish the biological functions. | [113] | ||
| The dynamics of hypothalamic methylation at puberty. | [114] | |||
| Long-term effects of endocrine-active compounds on corpus luteum of swine females exposed during early life period. | [115] | |||
| Porcine embryos | Investigation of the effects of histone deacetylase inhibitors on the in vitro development of porcine embryos derived from somatic cell nuclear transfer. | [100] | ||
| Porcine oocytes | The effects of vitamin C in the regulation of global epigenetic modifications at DNA, RNA and histones levels and its potential for oocyte maturation and developmental competence. | [99] | ||
| Porcine ovary | Epigenetic mechanisms of ovarian development during the transition from puberty and sexual maturation. | [116] | ||
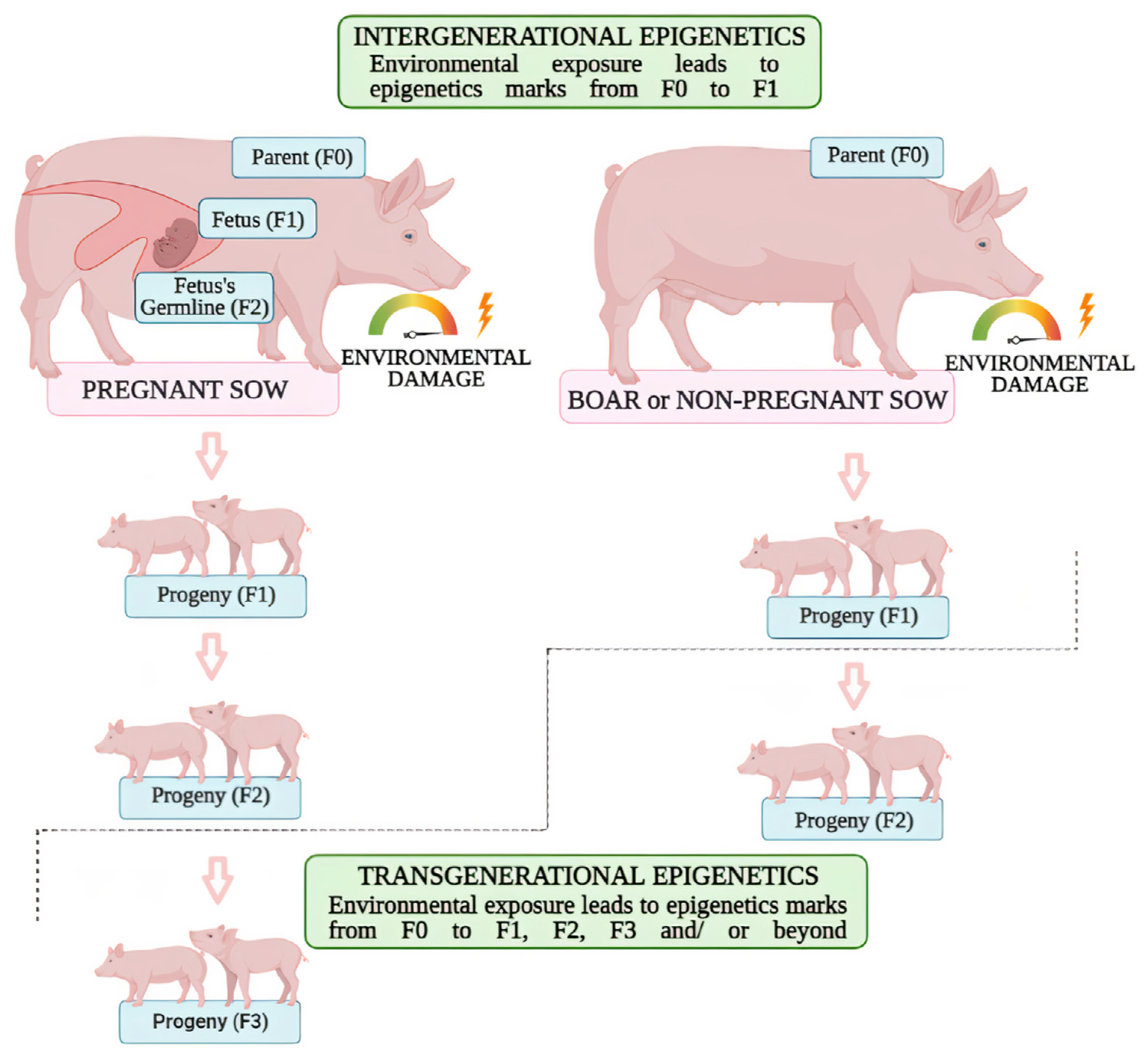
| Intergenerational epigenetics | F0–F1 | Pregnant sows and its offspring | Effects of exposure to low or high doses of estrogen during pregnancy and its role in female reproductive organs. | [89] |
| The immediate and long-term effects of maternal dietary protein affecting gene expression of offspring. | [93] | |||
| Restriction and excess dietary protein during pregnancy alters the offspring’s epigenetic marks and influences gene expression. | [94] | |||
| Boar’s semen and sow’s placenta | The role of breeding season in altering epigenetic components of the placenta and its consequences to foetal development. | [101] | ||
| Transgenerational epigenetics | F0–F2 | Boars | Transgenerational response of a methyl-enriched diet to boars and its responses on carcass traits, gene expression and DNA methylation. | [95] |
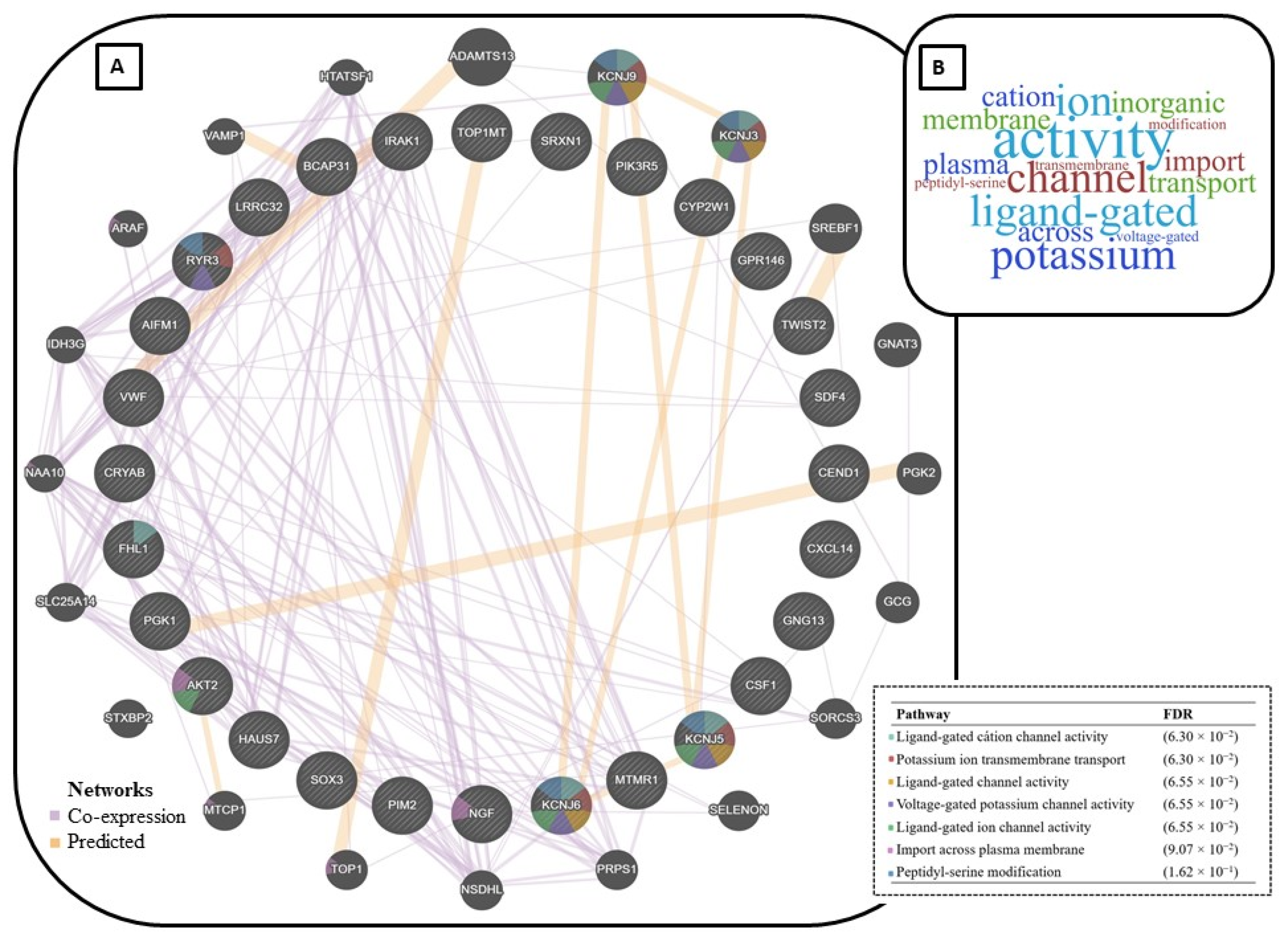
| Stress Source | Analysed Sample | Approach | Effect of Stress | Biomarker | Reference |
|---|---|---|---|---|---|
| Heat stress | Longissimus dorsi muscle | WGBS | DMRs in important genes involved in muscle development, metabolism, immunity, and stress response. | RYR3, PGK1, CRYAB and FHL1C | [15] |
| Intrauterine insult | Small intestine | RRBS | DMRs in several genes involved in cell development and immunity. | IRAK1, AIFM1, PIM2, BCAP31, MTMR1, SOX3, TWIST2 and HAUS7 | [96] |
| Sanitary challenge | Mammary epithelial cell | RRBS | DMRs in functional genes of the innate and adaptive immune response. | SDF4, SRXN1, CSF1 and CXCL14 | [117] |
| Mid intestine | RRBS | DMRS in genes involved in structural pathways of the cells with outcomes in the immature prenatal intestine. | CYP2W1, GPR146, TOP1MT and CEND1 | [102] | |
| Hippocampus | RRBS | DMRs in genes associated with blood brain barrier permeability and regulatory T-cell activation, which are reported to cause reductions in cognitive development. | VWF, LRRC32, NGF, GNG13, PIK3R5, KCNJ6, KCNJ5 and AKT2 | [112] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nery da Silva, A.; Silva Araujo, M.; Pértille, F.; Zanella, A.J. How Epigenetics Can Enhance Pig Welfare? Animals 2022, 12, 32. https://doi.org/10.3390/ani12010032
Nery da Silva A, Silva Araujo M, Pértille F, Zanella AJ. How Epigenetics Can Enhance Pig Welfare? Animals. 2022; 12(1):32. https://doi.org/10.3390/ani12010032
Chicago/Turabian StyleNery da Silva, Arthur, Michelle Silva Araujo, Fábio Pértille, and Adroaldo José Zanella. 2022. "How Epigenetics Can Enhance Pig Welfare?" Animals 12, no. 1: 32. https://doi.org/10.3390/ani12010032