
Impact of Dietary Sodium Butyrate and Salinomycin on Performance and Intestinal Microbiota in a Broiler Gut Leakage Model

 ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Experimental Design
2.2. Sample and Data Collection
2.3. Bacterial Enumeration
2.4. Analysis of Short-Chain Fatty Acid and Lactic Acid
2.5. DNA Extraction
2.6. 16S rRNA Gene Amplicon Sequencing
2.7. Statistical Analyses
2.7.1. Statistical Analysis of Performance Data
2.7.2. 16S rRNA Sequence Read Processing and Quality Control
2.7.3. Statistical Analysis of 16S rRNA Sequencing Data
3. Results
3.1. Production Performance
3.2. Enumeration of Bacteria
3.3. Organic Acids
3.4. pH
3.5. 16S rRNA Gene Amplicons
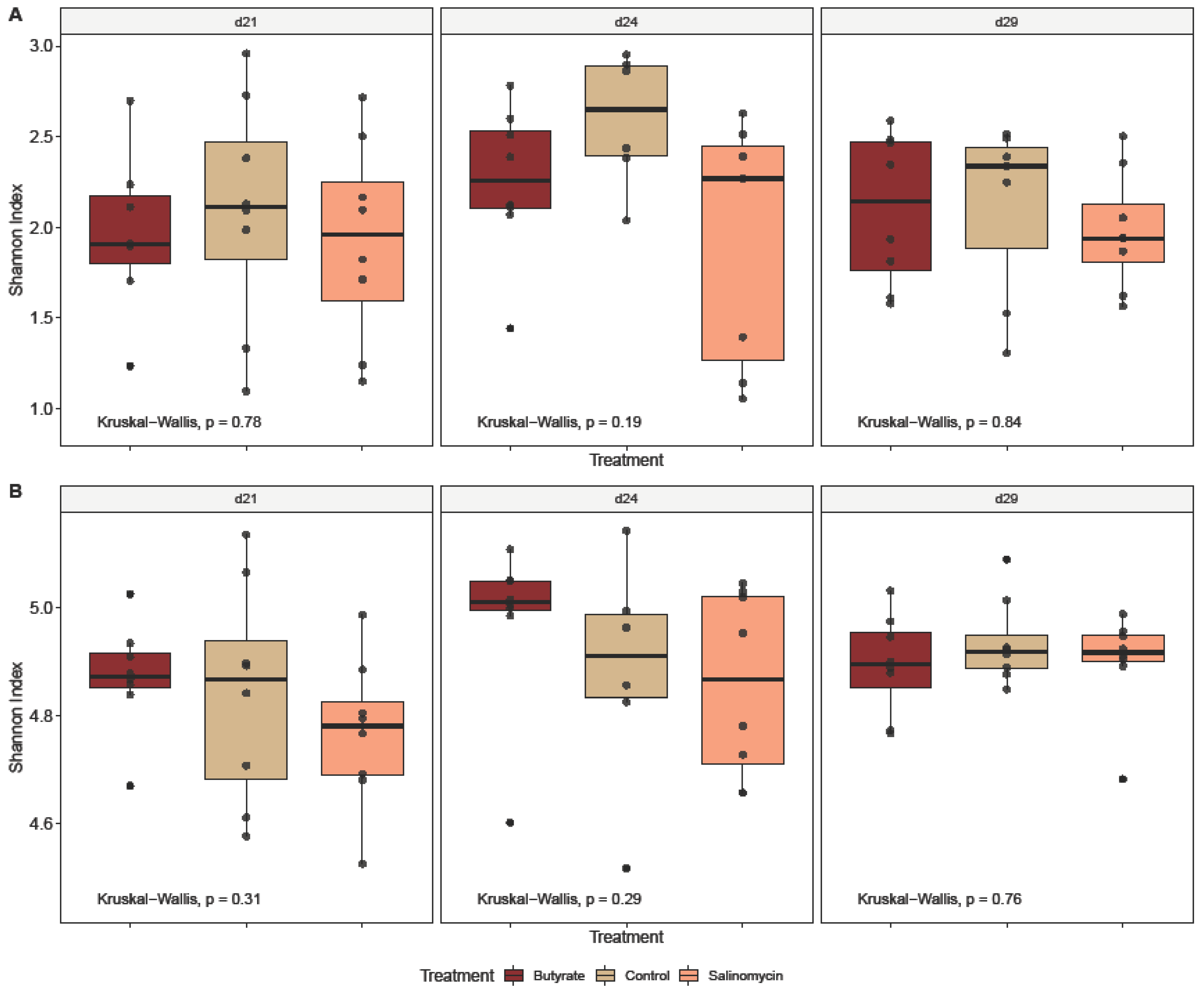
3.5.1. Dietary Effect on Microbial Alpha Diversity
3.5.2. Dietary Effect on Microbial Beta Diversity
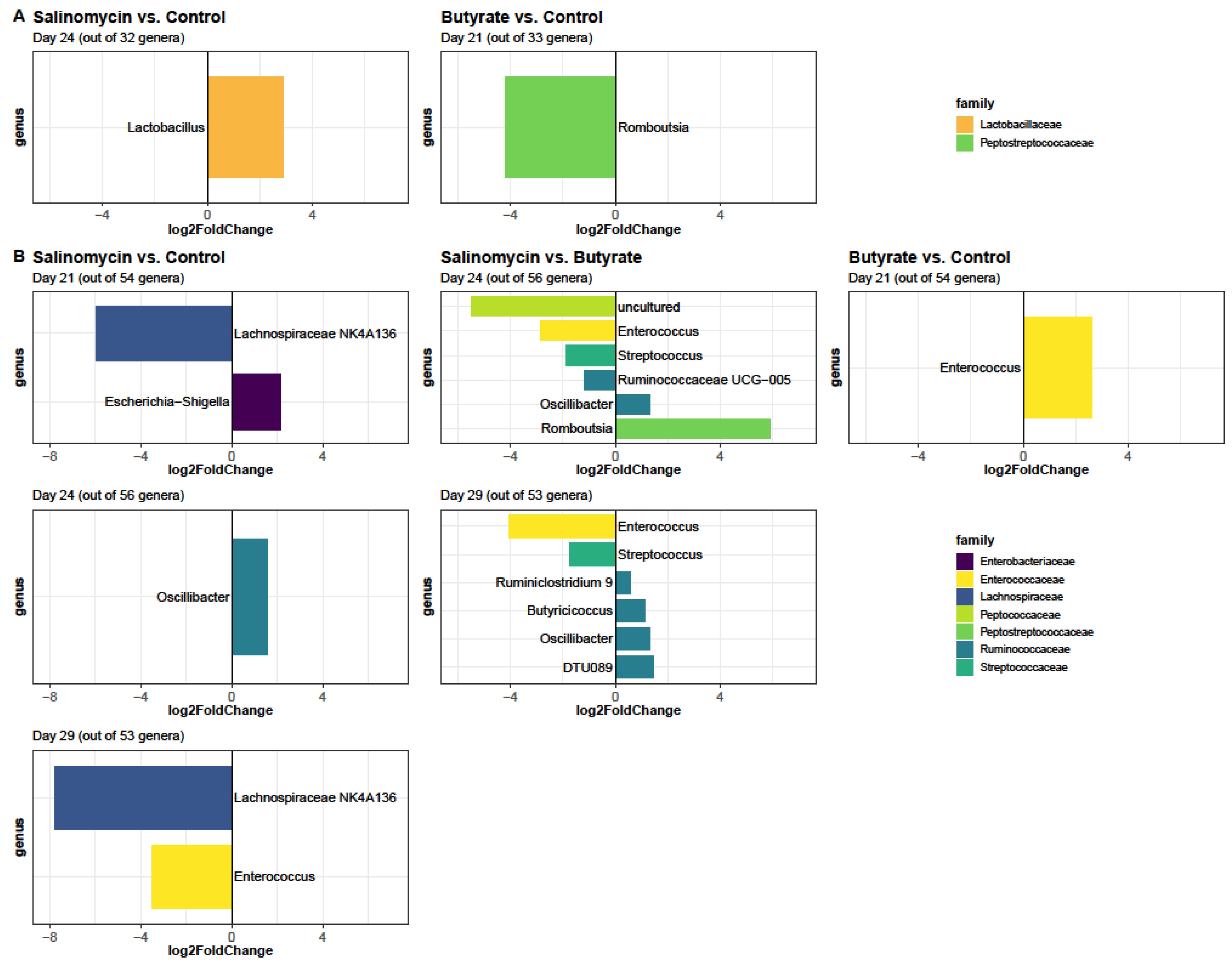
3.5.3. Dietary Effect on Microbiota in Different Segments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shang, Y.; Kumar, S.; Oakley, B.; Kim, W.K. Chicken gut microbiota: Importance and detection technology. Front. Vet. Sci. 2018, 5, 254. [Google Scholar] [CrossRef]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Kaunitz, J.D.; Akiba, Y. Control of Intestinal Epithelial Proliferation and Differentiation: The Microbiome, Enteroendocrine L Cells, Telocytes, Enteric Nerves, and GLP, Too. Dig. Dis. Sci. 2019, 64, 2709–2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carrasco, J.M.D.; Casanova, N.A.; Miyakawa, M.E.F. Microbiota, gut health and chicken productivity: What is the connection? Microorganisms 2019, 7, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crommen, S.; Simon, M.C. Microbial regulation of glucose metabolism and insulin resistance. Genes 2018, 9, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohrscheib, C.E.; Brownlie, J.C. Microorganisms that Manipulate Complex Animal Behaviours by Affecting the Host’s Nervous System. Springer Sci. Rev. 2013, 1, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Toor, D.; Wasson, M.K.; Kumar, P.; Karthikeyan, G.; Kaushik, N.K.; Goel, C.; Singh, S.; Kumar, A.; Prakash, H. Dysbiosis disrupts gut immune homeostasis and promotes gastric diseases. Int. J. Mol. Sci. 2019, 20, 2432. [Google Scholar] [CrossRef] [Green Version]
- Teirlynck, E.; Gussem, M.D.E.; Dewulf, J.; Haesebrouck, F.; Ducatelle, R.; van Immerseel, F. Morphometric evaluation of ‘dysbacteriosis’ in broilers. Avian Pathol. 2011, 40, 139–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venegas, D.P.; De La Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short chain fatty acids (SCFAs)mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Tellez, G.; Richards, J.D.; Escobar, J. Identification of Potential Biomarkers for Gut Barrier Failure in Broiler Chickens. Front. Vet. Sci. 2015, 2, 14. [Google Scholar] [CrossRef] [Green Version]
- Johansen, C.H.; Bjerrum, L.; Pedersen, K. Impact of salinomycin on the intestinal microflora of broiler chickens. Acta Vet. Scand. 2007, 49, 30. [Google Scholar] [CrossRef] [Green Version]
- Wong, A. Unknown Risk on the Farm: Does Agricultural Use of Ionophores Contribute to the Burden of Antimicrobial Resistance? mSphere 2019, 4, e00433-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Immerseel, F.; de Buck, J.; Pasmans, F.; Huyghebaert, G.; Haesebrouck, F.; Ducatelle, R. Clostridium perfringens in poultry: An emerging threat for animal and public health. Avian Pathol. 2004, 33, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Echeng, G.; Hao, H.; Exie, S.; Ewang, X.; Edai, M.; Ehuang, L.; Eyuan, Z.-H. Antibiotic alternatives: The substitution of antibiotics in animal husbandry? Front. Microbiol. 2014, 5, 217. [Google Scholar] [CrossRef] [Green Version]
- Mitic, L.L.; van Itallie, C.M.; Anderson, J.M. Molecular physiology and pathophysiology of tight junctions I. Tight junction structure and function: Lessons from mutant animals and proteins. Am. J. Physiol. Gastrointest. Liver Physiol. 2000, 279, G250–G254. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.; Ji, J.; Qu, H.; Wang, J.; Shu, D.; Wang, Y.; Liu, T.; Li, Y.; Luo, C. Effects of sodium butyrate on intestinal health and gut microbiota composition during intestinal inflammation progression in broilers. Poult. Sci. 2019, 98, 4449–4456. [Google Scholar] [CrossRef] [PubMed]
- Ahsan, U.; Cengiz, Ö.; Raza, I.; Kuter, E.; Chacher, M.; Iqbal, Z.; Umar, S.; Çakir, S. Sodium butyrate in chicken nutrition: The dynamics of performance, gut microbiota, gut morphology, and immunity. Worlds. Poult. Sci. J. 2018, 74, 156. [Google Scholar] [CrossRef] [Green Version]
- Tellez, G.; Latorre, J.D.; Kuttappan, V.A.; Hargis, B.M.; Hernandez-Velasco, X. Rye affects bacterial translocation, intestinal viscosity, microbiota composition and bone mineralization in Turkey poults. PLoS ONE 2015, 10, e0122390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tellez, G.; Latorre, J.D.; Kuttappan, V.A.; Kogut, M.H.; Wolfenden, A.; Hernandez-Velasco, X.; Hargis, B.M.; Bottje, W.G.; Bielke, L.R.; Faulkner, O.B. Utilization of rye as energy source affects bacterial translocation, intestinal viscosity, microbiota composition, and bone mineralization in broiler chickens. Front. Genet. 2014, 5, 339. [Google Scholar] [CrossRef] [PubMed]
- Baxter, M.F.A.; Latorre, J.D.; Dridi, S.; Merino-Guzman, R.; Hernandez-Velasco, X.; Hargis, B.M.; Tellez-Isaias, G. Identification of serum biomarkers for intestinal integrity in a broiler chicken malabsorption model. Front. Vet. Sci. 2019, 6, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naghizadeh, M.; Dalgaard, T.S.; Klaver, L.; Engberg, R.M. Effects of encapsulated butyrate and salinomycin on gut leakage and inflammation in broilers. Br. Poult. Sci. 2021. accepted. [Google Scholar]
- Engberg, R.M.; Grevsen, K.; Ivarsen, E.; Fretté, X.; Christensen, L.P.; Højberg, O.; Jensen, B.B.; Canibe, N. The effect of Artemisia annua on broiler performance, on intestinal microbiota and on the course of a Clostridium perfringens infection applying a necrotic enteritis disease model. Avian Pathol. 2012, 41, 369–376. [Google Scholar] [CrossRef] [Green Version]
- Engberg, R.M.; Hedemann, M.S.; Leser, T.D.; Jensen, B.B. Effect of zinc bacitracin and salinomycin on intestinal microflora and performance of broilers. Poult. Sci. 2000, 79, 1311–1319. [Google Scholar] [CrossRef] [PubMed]
- Canibe, N.; Højberg, O.; Badsberg, J.H.; Jensen, B.B. Effect of feeding fermented liquid feed and fermented grain on gastrointestinal ecology and growth performance in piglets. J. Anim. Sci. 2007, 85, 2959–2971. [Google Scholar] [CrossRef] [Green Version]
- Illumina. 16S Metagenomic Sequencing Library Preparation; Illumina: Foster City, CA, USA, 2013; pp. 1–28. [Google Scholar]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Project: Vienna, Austria, 2020. [Google Scholar]
- Douglas, A.; Maechler, M. Package ‘lme4’, 2014, 2015. Available online: https://cran.r-project.org/web/packages/lme4/lme4.pdf (accessed on 22 June 2021).
- Bretz, F.; Westfall, P.; Heiberger, R.M.; Schuetzenmeister, A.; Scheibe, S. Package ‘Multcomp’. 2021. Available online: https://cran.r-project.org/web/packages/multcomp/multcomp.pdf (accessed on 29 April 2021).
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Cisternas, P. DADA2: High resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 4–5. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, J.G. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, B.; Simpson, G.; Solymos, P.; Stevens, H.; Wagner, H. Vegan: Community Ecology Package. 2020. Available online: https://cran.r-project.org/package=vegan (accessed on 28 November 2020).
- Lahti, L.; Shetty, S. Tools for Microbiome Analysis in R. Version 2017. Available online: http://microbiome.github.com/microbiome (accessed on 28 November 2020).
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, K.S.; Kirkegaard, R.H.; Karst, S.M.; Albertsen, M. ampvis2: An R package to analyse and visualise 16S rRNA amplicon data. bioRxiv 2018. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Wu, W.; Xiao, Z.; An, W.; Dong, Y.; Zhang, B. Dietary sodium butyrate improves intestinal development and function by modulating the microbial community in broilers. PLoS ONE 2018, 13, e0197762. [Google Scholar] [CrossRef] [PubMed]
- Czerwiński, J.; Højberg, O.; Smulikowska, S.; Engberg, R.M.; Mieczkowska, A. Effects of sodium butyrate and salinomycin upon intestinal microbiota, mucosal morphology and performance of broiler chickens. Arch. Anim. Nutr. 2012, 66, 102–116. [Google Scholar] [CrossRef] [PubMed]
- Onrust, L.; Baeyen, S.; Haesebrouck, F.; Ducatelle, R.; van Immerseel, F. Effect of in feed administration of different butyrate formulations on Salmonella Enteritidis colonization and cecal microbiota in broilers. Vet. Res. 2020, 51, 56. [Google Scholar] [CrossRef] [Green Version]
- Antoszczak, M.; Steverding, D.; Huczyński, A. Anti-parasitic activity of polyether ionophores. Eur. J. Med. Chem. 2019, 166, 32–47. [Google Scholar] [CrossRef] [Green Version]
- Robinson, K.; Becker, S.; Xiao, Y.; Lyu, W.; Yang, Q.; Zhu, H.; Yang, H.; Zhao, J.; Zhang, G. Differential Impact of Subtherapeutic Antibiotics and Ionophores on Intestinal Microbiota of Broilers. Microorganisms 2019, 7, 282. [Google Scholar] [CrossRef] [Green Version]
- Rychlik, I. Composition and function of chicken gut microbiota. Animals 2020, 10, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjitkar, S.; Lawley, B.; Tannock, G.; Engberg, R.M. Bacterial Succession in the Broiler Gastrointestinal Tract. Appl. Environ. Microbiol. 2016, 82, 2399–2410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.M.; Seddiek, S.A.; Khater, H.F. Effect of butyrate, clopidol and their combination on the performance of broilers infected with Eimeria maxima. Br. Poult. Sci. 2014, 55, 474–482. [Google Scholar] [CrossRef]
- Van Immerseel, F.; Fievez, V.; De Buck, J.; Pasmans, F.; Martel, A.; Haesebrouck, F.; Ducatelle, R. Microencapsulated short-chain fatty acids in feed modify colonization and invasion early after infection with Salmonella enteritidis in young chickens. Poult. Sci. 2004, 83, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Van Immerseel, F.; Boyen, F.; Gantois, I.; Timbermont, L.; Bohez, L.; Pasmans, F.; Haesebrouck, F.; Ducatelle, R. Supplementation of coated butyric acid in the feed reduces colonization and shedding of Salmonella in poultry. Poult. Sci. 2005, 84, 1851–1856. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.Y.; Inohara, N.; Nuñez, G. Mechanisms of inflammation-driven bacterial dysbiosis in the gut. Mucosal Immunol. 2017, 10, 18–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, D.; Schwab, C.; Milinovich, G.; Reichert, J.; Ben Mahfoudh, K.; Decker, T.; Engel, M.; Hai, B.; Hainzl, E.; Heider, S.; et al. Phylotype-level 16S rRNA analysis reveals new bacterial indicators of health state in acute murine colitis. ISME J. 2012, 6, 2091–2106. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.C.-H. Microbiota dysbiosis and barrier dysfunction in inflammatory bowel disease and colorectal cancers: Exploring a common ground hypothesis. J. Biomed. Sci. 2018, 25, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lodemann, U.; Amasheh, S.; Radloff, J.; Kern, M.; Bethe, A.; Wieler, L.H.; Pieper, R.; Zentek, J.; Aschenbach, J.R. Effects of Ex Vivo Infection with ETEC on Jejunal Barrier Properties and Cytokine Expression in Probiotic-Supplemented Pigs. Dig. Dis. Sci. 2017, 62, 922–933. [Google Scholar] [CrossRef]
- Kreuzer, S.; Machnowska, P.; Aßmus, J.; Sieber, M.; Pieper, R.; Schmidt, M.F.; Brockmann, G.A.; Scharek-Tedin, L.; Johne, R. Feeding of the probiotic bacterium Enterococcus faecium NCIMB 10415 differentially affects shedding of enteric viruses in pigs. Vet. Res. 2012, 43, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Zheng, A.; Meng, K.; Chang, W.; Bai, Y.; Li, K.; Cai, H.; Liu, G.; Yao, B. Proteome changes in the intestinal mucosa of broiler (Gallus gallus) activated by probiotic Enterococcus faecium. J. Proteom. 2013, 91, 226–241. [Google Scholar] [CrossRef]
- Nisbet, D.J.; Callaway, T.R.; Edrington, T.S.; Anderson, R.C.; Poole, T.L. Effects of ionophores on Enterococcus faecalis and E. faecium growth in pure and mixed ruminal culture. Foodborne Pathog. Dis. 2008, 5, 193–198. [Google Scholar] [CrossRef]
- Danzeisen, J.L.; Kim, H.B.; Isaacson, R.E.; Tu, Z.J.; Johnson, T.J. Modulations of the chicken cecal microbiome and metagenome in response to anticoccidial and growth promoter treatment. PLoS ONE 2011, 6, e27949. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yin, F.; Yang, Y.; Lepp, D.; Yu, H.; Ruan, Z.; Yang, C.; Yin, Y.; Hou, Y.; Leeson, S.; et al. Dietary butyrate glycerides modulate intestinal microbiota composition and serum metabolites in broilers. Sci. Rep. 2018, 8, 4940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; de Angelis, M. The controversial role of human gut lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef] [PubMed]
- Hang, I.; Rinttila, T.; Zentek, J.; Kettunen, A.; Alaja, S.; Apajalahti, J.; Harmoinen, J.; de Vos, W.M.; Spillmann, T. Effect of high contents of dietary animal-derived protein or carbohydrates on canine faecal microbiota. BMC Vet. Res. 2012, 8, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torok, V.A.; Allison, G.E.; Percy, N.J.; Ophel-Keller, K.; Hughes, R.J. Influence of antimicrobial feed additives on broiler commensal posthatch gut microbiota development and performance. Appl. Environ. Microbiol. 2011, 77, 3380–3390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trela, J.; Kierończyk, B.; Hautekiet, V.; Józefiak, D. Combination of Bacillus licheniformis and Salinomycin: Effect on the Growth Performance and GIT Microbial Populations of Broiler Chickens. Animals 2020, 10, 889. [Google Scholar] [CrossRef] [PubMed]
- Lacey, J.A.; Stanley, D.; Keyburn, A.L.; Ford, M.; Chen, H.; Johanesen, P.; Lyras, D.; Moore, R.J. Clostridium perfringens-mediated necrotic enteritis is not influenced by th67e pre-existing microbiota but is promoted by large changes in the post-challenge microbiota. Vet. Microbiol. 2018, 227, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Timbermont, L.; Haesebrouck, F.; Ducatelle, R.; van Immerseel, F. Necrotic enteritis in broilers: An updated review on the pathogenesis. Avian Pathol. 2011, 40, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Knarreborg, A.; Engberg, R.M.; Jensen, S.K.; Jensen, B.B. Quantitative determination of bile salt hydrolase activity in bacteria isolated from the small intestine of chickens. Appl. Environ. Microbiol. 2002, 68, 6425–6428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qaisrani, S.N.; van Krimpen, M.M.; Verstegen, M.W.A.; Hendriks, W.H.; Kwakkel, R.P. Effects of three major protein sources on performance, gut morphology and fermentation characteristics in broilers. Br. Poult. Sci. 2020, 61, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Guban, J.; Korver, D.R.; Allison, G.E.; Tannock, G.W. Relationship of dietary antimicrobial drug administration with broiler performance, decreased population levels of Lactobacillus salivarius, and reduced bile salt deconjugation in the ileum of broiler chickens. Poult. Sci. 2006, 85, 2186–2194. [Google Scholar] [CrossRef] [PubMed]
- Knarreborg, A.; Lauridsen, C.; Engberg, R.M.; Jensen, S.K. Dietary antibiotic growth promoters enhance the bioavailability of α-tocopheryl acetate in broilers by altering lipid absorption. J. Nutr. 2004, 134, 1487–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dietary Treatment | ||||||
|---|---|---|---|---|---|---|
| Item | Age (day) | Control | Butyrate | Salinomycin | SEM | p Value |
| Body weight (g/day/bird) | 10 | 249 | 253 | 260 | 3.1 | >0.05 |
| 23 | 653 b | 673 b | 770 a | 16.6 | <0.001 | |
| 34 | 787 b | 822 b | 1104 a | 25.0 | <0.001 | |
| Body weight gain (g/day) | 1–10 | 24.9 | 25.3 | 26 | 0.31 | 0.063 |
| 11–23 | 50.2 b | 51.8 b | 59.2 a | 1.27 | <0.001 | |
| 24–34 | 71.6 b | 74.7 b | 100.4 a | 2.27 | <0.001 | |
| 1–34 | 59.7 b | 61.9 b | 72.8 a | 1.66 | <0.001 | |
| Feed intake (g/day/bird) | 1–10 | 29.7 | 28.9 | 29.1 | 0.88 | >0.05 |
| 11–23 | 73.6 b | 78.5 b | 83.8 a | 1.63 | <0.001 | |
| 24–34 | 119 b | 123 b | 148 a | 3.77 | <0.001 | |
| 1–34 | 78.7 b | 81.8 b | 92.2 a | 1.78 | <0.001 | |
| Water intake (mL/day/bird) | 1–10 | 61.8 | 62.2 | 62.5 | 1.18 | >0.05 |
| 11–23 | 140 b | 146 ab | 154 a | 3.26 | 0.024 | |
| 24–34 | 198 b | 207 b | 238 a | 6.57 | <0.001 | |
| 1–34 | 134 b | 139 b | 152 a | 3.12 | 0.002 | |
| FCR (kg diet/kg weight gain) | 1–10 | 1.19 | 1.14 | 1.10 | 0.026 | 0.076 |
| 11–23 | 1.47 ab | 1.52 a | 1.42 b | 0.021 | 0.013 | |
| 24–34 | 1.66 a | 1.64 a | 1.47 b | 0.044 | 0.006 | |
| 1–34 | 1.32 a | 1.32 a | 1.27 b | 0.016 | 0.046 | |
| Water/feed intake ratio | 1–10 | 2.11 | 2.15 | 2.15 | 0.065 | >0.05 |
| 11–23 | 1.91 ab | 1.87 b | 2.01 a | 0.035 | 0.022 | |
| 24–34 | 1.67 b | 1.70 b | 1.93 a | 0.055 | 0.005 | |
| 1–34 | 1.71 a | 1.71 a | 1.65 b | 0.016 | 0.025 | |
| Dietary Treatment | Age (day) | p Value | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Item | Control | Butyrate | Salinomycin | SEM | 21 | 24 | 29 | SEM | Diet | Age | Diet × Age |
| Lactic acid bacteria | |||||||||||
| Ileum | 8.47 | 8.23 | 8.36 | 0.09 | 8.62 a | 8.25 bc | 8.05 c | 0.09 | 0.181 | <0.001 | 0.004 |
| Ceca | 9.11 a | 9.14 a | 8.93 b | 0.05 | 9.30 a | 9.02 b | 8.89 b | 0.06 | 0.006 | <0.001 | 0.002 |
| Coliform bacteria | |||||||||||
| Ileum | 6.37 | 6.48 | 6.63 | 0.14 | 6.19 b | 6.54 ab | 6.48 ab | 0.14 | 0.362 | 0.020 | 0.102 |
| Ceca | 8.61 a | 8.69 a | 8.44 b | 0.07 | 8.07 b | 8.84 a | 8.76 a | 0.08 | 0.032 | <0.001 | 0.342 |
| Lactose-negative enterobacteria | |||||||||||
| Ileum | 4.66 b | 4.46 b | 5.03 a | 0.14 | 4.27 b | 4.29 b | 4.72 b | 0.14 | 0.011 | <0.001 | 0.014 |
| Ceca | 6.45 | 6.46 | 6.44 | 0.14 | 6.00 b | 6.72 a | 6.54 a | 0.11 | 1.000 | <0.001 | 0.006 |
| Enterococci | |||||||||||
| Ileum | 6.75 | 6.77 | 6.97 | 0.10 | 6.70 b | 6.79 ab | 6.72 b | 0.10 | 0.222 | 0.009 | 0.332 |
| Ceca | 7.46 | 7.45 | 7.30 | 0.08 | 7.31 b | 7.68 a | 7.34 ab | 0.09 | 0.289 | 0.010 | 0.012 |
| Clostridium perfringens | |||||||||||
| Ileum | 3.20 a | 3.39 a | 2.05 b | 0.28 | 2.79 | 3.13 | 2.78 | 0.25 | 0.003 | 0.621 | 0.351 |
| Ceca | 4.11 a | 4.41 a | 2.34 b | 0.35 | 4.31 a | 3.87 a | 3.49 ab | 0.28 | <0.001 | <0.001 | 0.056 |
| Lactobacillus salivarius | |||||||||||
| Ileum | 7.40 | 7.27 | 6.95 | 0.14 | 7.33 a | 7.49 a | 6.58 b | 0.13 | 0.056 | <0.001 | 0.851 |
| Ceca | 8.39 a | 8.45 a | 8.01 b | 0.07 | 8.58 a | 8.48 a | 7.62 b | 0.08 | <0.001 | <0.001 | 0.047 |
| Dietary Treatment | Age (day) | p-Value | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Item | Control | Butyrate | Salinomycin | SEM | 21 | 24 | 29 | SEM | Diet | Age | Diet × Age |
| Ileum | |||||||||||
| Lactic acid | 40.9 a | 28.9 b | 31.6 ab | 5.24 | 26.9 bc | 37.4 b | 18.0 c | 5.61 | 0.036 | <0.001 | 0.038 |
| Acetic acid | 4.8 | 4.4 | 4.78 | 0.29 | 3.30 c | 4.47 b | 4.70 b | 0.33 | 0.13 | <0.001 | 0.12 |
| Formic acid | 0.58 | 0.58 | 0.57 | 0.08 | 0.18 c | 0.50 bc | 0.54 b | 0.09 | 0.99 | <0.001 | 0.025 |
| Succinic acid | 0.31 a | 0.22 ab | 0.15 b | 0.06 | 0.28 a | 0.31 a | 0.00 b | 0.06 | 0.043 | <0.001 | 0.025 |
| Ceca | |||||||||||
| Acetic acid | 75.3 | 77.1 | 81.9 | 2.22 | 77.2 b | 71.9 b | 74.1 b | 2.56 | 0.082 | <0.001 | 0.19 |
| Succinic acid | 5.29 b | 5.59 ab | 8.57 a | 0.87 | 10.3 a | 5.98 b | 5.76 b | 0.97 | <0.001 | <0.001 | 0.025 |
| Propionic acid | 4.03 | 4.59 | 4.34 | 0.17 | 4.21 bc | 4.28 b | 3.51 c | 0.2 | 0.06 | <0.001 | 0.17 |
| Isobutyric acid | 0.54 b | 0.69 a | 0.44 c | 0.03 | 0.45 b | 0.66 a | 0.48 b | 0.03 | <0.001 | <0.001 | 0.089 |
| Butyric acid | 18.5 | 18.6 | 21.1 | 0.9 | 19.5 b | 17.6 b | 16.7 b | 1.03 | 0.054 | <0.001 | 0.009 |
| Isovaleric acid | 0.26 b | 0.35 a | 0.20 b | 0.02 | 0.23 b | 0.34 a | 0.23 b | 0.02 | <0.001 | <0.001 | 0.45 |
| Valeric acid | 1.02 ab | 1.16 a | 0.96 b | 0.04 | 1.02 ab | 1.08 a | 0.90 b | 0.04 | 0.003 | <0.001 | 0.07 |
| Lactic acid | 0.66 | 0.28 | 0.46 | 0.29 | 0.17 | 1.02 | 0.37 | 0.33 | 0.61 | 0.25 | 0.43 |
| Item | Dietary Treatment | SEM | Age (day) | SEM | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | Butyrate | Salinomycin | 21 | 24 | 29 | Diet | Days | Diet x Days | |||
| Ileum | 6.30 | 6.45 | 6.54 | 0.10 | 6.18 | 6.34 | 6.69 | 0.09 | 0.20 | <0.001 | 0.02 |
| Ceca | 6.43 a | 6.38 a | 6.09 b | 0.06 | 6.10 | 6.42 | 6.34 | 0.07 | <0.001 | 0.780 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naghizadeh, M.; Klaver, L.; Schönherz, A.A.; Rani, S.; Dalgaard, T.S.; Engberg, R.M. Impact of Dietary Sodium Butyrate and Salinomycin on Performance and Intestinal Microbiota in a Broiler Gut Leakage Model. Animals 2022, 12, 111. https://doi.org/10.3390/ani12010111
Naghizadeh M, Klaver L, Schönherz AA, Rani S, Dalgaard TS, Engberg RM. Impact of Dietary Sodium Butyrate and Salinomycin on Performance and Intestinal Microbiota in a Broiler Gut Leakage Model. Animals. 2022; 12(1):111. https://doi.org/10.3390/ani12010111
Chicago/Turabian StyleNaghizadeh, Mohammad, Laura Klaver, Anna A. Schönherz, Sundas Rani, Tina Sørensen Dalgaard, and Ricarda Margarete Engberg. 2022. "Impact of Dietary Sodium Butyrate and Salinomycin on Performance and Intestinal Microbiota in a Broiler Gut Leakage Model" Animals 12, no. 1: 111. https://doi.org/10.3390/ani12010111