
Retinol and α-Tocopherol Contents, Fat Color, and Lipid Oxidation as Traceability Tools of the Feeding System in Suckling Payoya Kids

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Handling and Feeding
- Mountain grazing (MG): 12 suckling kids were slaughtered at an average age of 37 days. During lactation, their mothers grazed in a dehesa system [28] for 8 h and were stabled for the remaining time. In addition, dams were supplemented with 800 g of commercial feed.
- Cultivated meadow (CM): 13 suckling kids were slaughtered at an average age of 32 days. During lactation, their mothers grazed in a cultivated pasture of oats for 8 h (the remainder of the time, they were stabled) and were supplemented with 500 g of compound feed (28.45% of the same commercial feed as the MG kid + 35% sunflower seed (Helianthus annuus) + 28.3% oat seed (Avena sativa) + 8.25% pea seed (Pisum sativum)).
- Total mixed ration (TMR): 14 suckling kids were slaughtered at an average age of 30 days. Their mothers were kept permanently indoors and fed 1.5 kg of the same commercial feed and hay ad libitum.Further details of the dam feeding systems and management were described in a previous study [29].
- Milk replacer (MR): Sixteen kids were fed a milk replacer (CORDEVIT calostrado-50, Lemasa, Spain) enhanced with 80,000 IU/kg of vitamin A and 30 IU/kg of vitamin E. The composition of the milk replacer administered to kids is shown in Table 1. They were slaughtered at 47 days old. These kids ingested approximately 1.25 L of milk per day [30], with a concentration of 200 g of power per liter, as indicated in the product instructions. Table 1 shows the composition of the milk replacer.
2.2. Slaughter, Plasma Collection, Carcass Characteristics, and Sampling
2.3. Analytical Procedures
2.3.1. Fat Color
2.3.2. Intramuscular Fat Amount, Lipid Oxidation, and Vitamin Extraction
2.3.3. Extraction and Determinations of Vitamins from Plasma
2.4. HPLC Conditions
2.5. Statistical Analysis
3. Results and Discussion
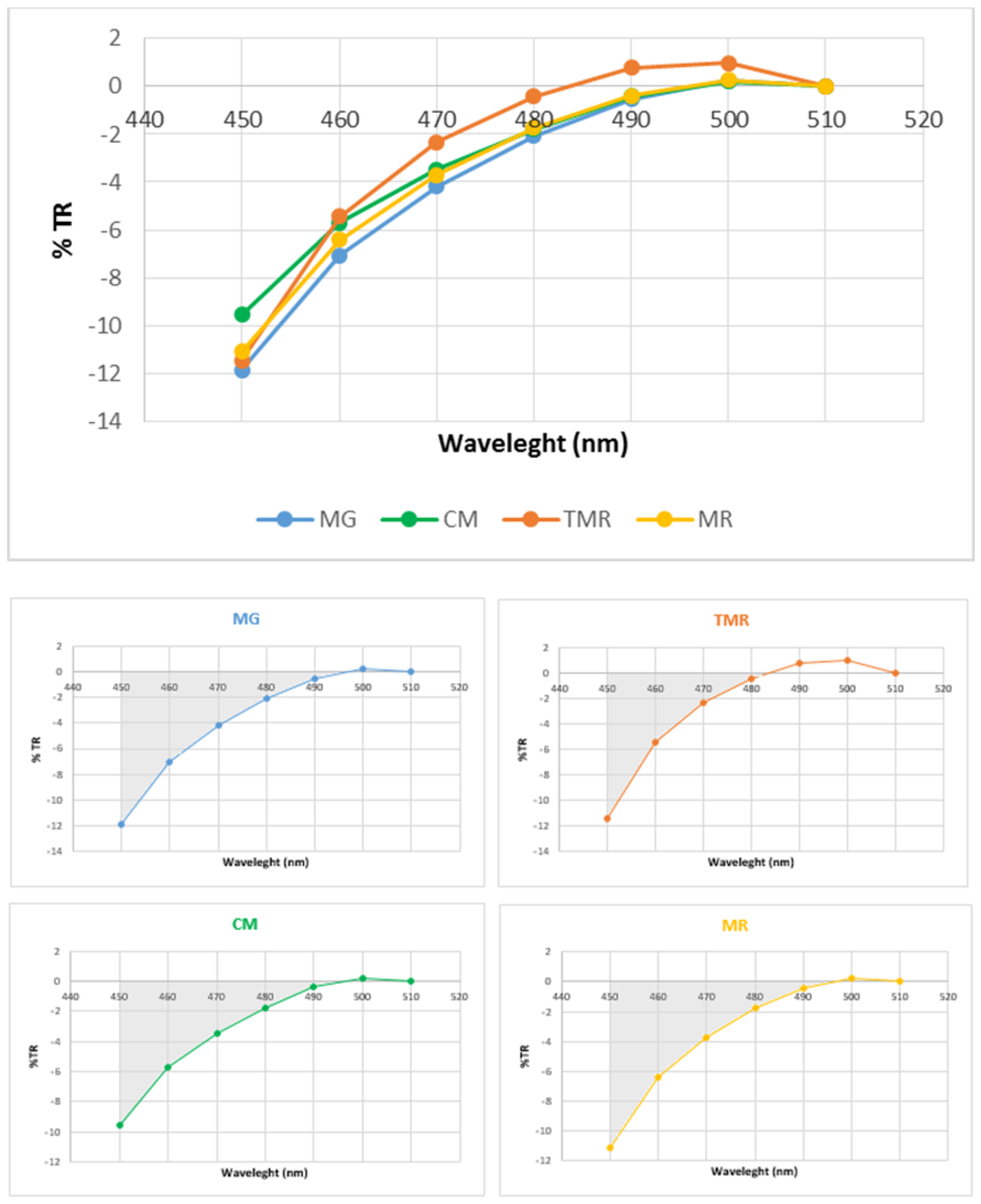
3.1. Carcass Traits and Kidney Fat Color Parameters
3.2. Intramuscular Fat Content and Lipid Oxidation
3.3. Retinol and α-Tocopherol Contents
3.3.1. Concentrations in Intermuscular Fat
3.3.2. Concentration in Plasma
3.4. Discriminant Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ripoll, G.; Alcalde, M.J.; Horcada, A.; Campo, M.M.; Sañudo, C.; Teixeira, A.; Panea, B. Effect of slaughter weight and breed on instrumental and sensory meat quality of suckling kids. Meat Sci. 2012, 92, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Castel, J.M.; Ruiz, F.A.; Mena, Y.; Sánchez-Rodríguez, M. Present situation and future perspectives for goat production systems in Spain. Small Rumin. Res. 2010, 89, 207–210. [Google Scholar] [CrossRef]
- MAPA. Anuario de Estadìstica Agraria. Ministerio de Agricultura Pesca y Alimentación. Available online: https://www.mapa.gob.es/estadistica/pags/anuario/2019/anuario/AE19.pdf (accessed on 1 February 2021).
- Mena, Y.; Ligero, M.; Ruiz, F.A.; Nahed, J.; Castel, J.M.; Acosta, J.M.; Guzmán, J.L. Organic and Conventional Dairy Goat Production Systems in Andalusian Mountainous Areas. In Changes in Sheep and Goat Farming Systems at the Beginning of the 21st Century: Research, Tools, Methods and Initiatives in Favour of a Sustainable Development; Pacheco, F., Morand-Fehr, P., Eds.; Options Méditerranéennes: Série, A. Séminaires Méditerranéens; n. 91; CIHEAM/DRAP-Norte/FAO: Zaragoza, Spain, 2009; pp. 253–256. Available online: http://hdl.handle.net/11441/43920 (accessed on 24 October 2021).
- Ripoll, G.; Alcalde, M.J.; Horcada, A.; Sañudo, C.; Panea, B. Influencia del sistema de lactancia sobre el color de la grasa de la carne y la grasa renal de cabritos de las razas Murciano-Granadina y Malagueña. XIII Jorn. Sobre Prod. Anim. (AIDA) 2009, II, 589–591. Available online: https://www.aida-itea.org/aida-itea/files/jornadas/2009/comunicaciones/2009_CdP_35.pdf (accessed on 15 August 2021).
- Bañón, S.; Vila, R.; Price, A.; Ferrandini, E.; Garrido, M.D. Effects of goat milk or milk replacer diet on meat quality and fat composition of suckling goat kids. Meat Sci. 2006, 72, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Morales-Jerrett, E.; Mancilla-Leytón, J.M.; Delgado-Pertíñez, M.; Mena, Y. The contribution of traditional meat goat farming systems to human wellbeing and its importance for the sustainability of this livestock subsector. Sustainability 2020, 12, 1181. [Google Scholar] [CrossRef] [Green Version]
- Castel, J.; Mena, Y.; Ruiz, F.A.; Camúñez-Ruiz, J.; Sánchez-Rodríguez, M. Changes occurring in dairy goat production systems in less favoured areas of Spain. Small Rumin. Res. 2011, 96, 83–92. [Google Scholar] [CrossRef]
- Bernués, A.; Ruiz, R.; Olaizola, A.; Villalba, D.; Casasús, I. Sustainability of pasture-based livestock farming systems in the European Mediterranean context: Synergies and trade-offs. Livest. Sci. 2011, 139, 44–57. [Google Scholar] [CrossRef]
- Zervas, G.; Tsiplakou, E. The effect of feeding systems on the characteristics of products from small ruminants. Small Rumin. Res. 2011, 101, 140–149. [Google Scholar] [CrossRef]
- Alcalde, M.J.; Ripoll, G.; Panea, B. Consumer attitudes towards meat consumption in Spain with special reference to quality marks and kid meat. In Consumer Attitudes to Food Quality Products; Klopčič, M., Kuipers, A., Hocquette, J.F., Eds.; EAAP—European Federation of Animal Science; Wageningen Academic Publishers: Wageningen, The Netherlands, 2013; Volume 133, pp. 97–107. [Google Scholar] [CrossRef]
- Storøy, J.; Thakur, M.; Olsen, P. The TraceFood Framework—Principles and guidelines for implementing traceability in food value chains. J. Food Eng. 2013, 115, 41–48. [Google Scholar] [CrossRef]
- van Dijka, H.; Houghton, J.; van Kleef, E.; van der Lans, I.; Rowe, G.; Frewer, L. Consumer responses to communication about food risk management. Appetite 2008, 50, 340–352. [Google Scholar] [CrossRef]
- Ballin, N.Z. Authentication of meat and meat products. Meat Sci. 2010, 86, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Tume, R.K.A. Comparison of beta-carotene-splitting activity isolated from intestinal mucosa of pasture-grazed sheep, goats and cattle. Biochem. Mol. Biol. Int. 1993, 30, 209–217. [Google Scholar]
- Prache, S.; Cornu, A.; Berdagué, J.L.; Priolo, A. Traceability of animal feeding diet in the meat and milk of small ruminants. Small Rumin. Res. 2005, 59, 157–168. [Google Scholar] [CrossRef]
- van den Berg, H.; Faulks, R.; Fernando Granado, H.; Hirschberg, J.; Olmedilla, B.; Sandmann, G.; Southon, S.; Stahl, W. The potential for the improvement of carotenoid levels in foods and the likely systemic effects. J. Sci. Food Agric. 2000, 80, 880–912. [Google Scholar] [CrossRef]
- Cozzolino, D.; De Mattos, D.; Vaz Martins, D. Visible/near infrared reflectance spectroscopy for predicting composition and tracing system of production of beef muscle. Anim. Sci. 2020, 74, 477–484. [Google Scholar] [CrossRef]
- Prache, S.; Theriez, M. Traceability of lamb production systems: Carotenoids in plasma and adipose tissue. Anim. Sci. 1999, 69, 29–36. [Google Scholar] [CrossRef]
- Alvarez, R.; Meléndez-Martinez, A.J.; Vicario, M.I.; Alcalde, M.J. Effect of pasture and concentrate diets on concentrations of carotenoids, vitamin A and vitamin E in plasma and adipose tissue of lambs. J. Food Compos. Anal. 2014, 36, 59–65. [Google Scholar] [CrossRef]
- Ripoll, G.; Joy, M.; Muñoz, F.; Alberíı, P. Meat and fat colour as a tool to trace grass-feeding systems in light lamb production. Meat Sci. 2008, 80, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Blanco, M.; Lobón, S.; Bertolín, J.R.; Joy, M. Effect of the maternal feeding on the carotenoid and tocopherol content of suckling lamb tissues. Arch. Anim. Nutr. 2019, 73, 472–484. [Google Scholar] [CrossRef] [PubMed]
- Rufino-Moya, P.J.; Joy, M.; Lobón, S.; Bertolín, J.R.; Blanco, M. Carotenoids and Liposoluble Vitamins in the Plasma and Tissues of Light Lambs Given Different Maternal. Animals 2020, 10, 1813. [Google Scholar] [CrossRef]
- European Union. Directive 2010/63/EU of the european parliament and of the council of 22 September 2010 on the proteccion of animals used for scientific purposes. Off. J. Eur. Union 2010, 276, 33–79. Available online: http://data.europa.eu/eli/dir/2010/63/oj (accessed on 14 January 2021).
- El Tiempo en Grazalema. Datos Pluviométricos Por Año Hidrológico. 2021. Available online: https://modaenlana.com/el-tiempo-en-grazalema/ (accessed on 21 June 2021).
- Delgado-Pertíñez, M.; Gutiérrez-Peña, R.; Mena, Y.; Fernández-Cabanás, V.M.; Laberye, D. Milk production, fatty acid composition and vitamin E content of Payoya goats according to grazing level in summer on Mediterranean shrublands. Small Rumin. Res. 2013, 114, 167–175. [Google Scholar] [CrossRef]
- MAPA. Catálogo Oficial de Razas Ganaderas. 2021. Available online: https://www.mapa.gob.es/es/ganaderia/temas/zootecnia/razas-ganaderas/razas/catalogo-razas/caprino/payoya/default.aspx (accessed on 14 February 2021).
- Ferrer, C.; San Miguel, A.; Olea, L. Nomenclator básico de pastos en España. Rev. Soc. Española Para El Estud. Los Pastos (SEEP) 2001, Pastos XXI, 7–44. [Google Scholar] [CrossRef]
- Roncero-Díaz, M.; Panea, B.; Argüello, A.; Alcalde, M.J. How Management System Affects the Concentration of Retinol and α-Tocopherol in Plasma and Milk of Payoya Lactating Goats: Possible Use as Traceability Biomarkers. Animals 2021, 11, 2326. [Google Scholar] [CrossRef]
- Peña, F.; Doménech, V.; Acero, R.; Perea, J.; García, A. Effect of Sex and Feed (Maternal Milk vs. Milk Substitute) on the Growth and Carcass Characteristics in Florida Goat Kids. Rev. Científica Fac. Cienc. Vet. Univ. Zulia 2009, 19, 619–629. Available online: http://www.redalyc.org/articulo.oa?id=95911621009 (accessed on 14 October 2021).
- European Union. Council Regilation (EC) No 1099/2009 of 24 September 2009 on the Protection of Animals at the Time of Killing. Off. J. Eur. Union 2009, 303, 30. Available online: http://data.europa.eu/eli/reg/2009/1099/oj (accessed on 20 January 2021).
- Commission Internationale de L’Eclairage. Colorimetry, 2nd ed.; CIE No. 15.2, 78 + vi: 64–65; Central Bureau of the CIE: Vienna, Austria, 1986. [Google Scholar] [CrossRef]
- Priolo, A.; Prache, S.; Micol, D.; Agabriel, J. Reflectance spectrum of adipose tissue to trace grass feeding in sheep: Influence of measurement site and shrinkage time after slaughter. J. Anim. Sci. 2002, 80, 886–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bligh, E.G.; Dyer, W. A rapid method of total lipid extration and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, G.; Jørgensen, S.S. A critical examination of some experimental variables in the 2-thiobarbituric acid (TBA) test for lipid oxidation in meat products. Z. Lebensm.-Unters. Forsch. 1996, 202, 205–210. [Google Scholar] [CrossRef]
- Nozière, P.; Graulet, B.; Lucas, A.; Martin, B.; Grolier, P.; Doreau, M. Carotenoids for ruminants: From forages to dairy products. Anim. Feed. Sci. Technol. 2006, 131, 418–450. [Google Scholar] [CrossRef]
- Lyan, B.; Azaïs-Braesco, V.; Cardinault, N.; Tyssandier, V.; Borel, P.; Alexandre-Gouabau, M.C.; Grolier, P. Simple method for clinical determination of 13 carotenoids in human plasma using an isocratic high-performance liquid chromatographic method. J. Chromatogr. B: Biomed. Sci. Appl. 2001, 751, 297–303. [Google Scholar] [CrossRef]
- Mouly, P.P.; Gaydou, E.M.; Corsetti, J. Determination of the geographical origin of Valencia orange juice using carotenoid liquid chromatographic profiles. J. Chromatogr. A 1999, 844, 149–159. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.J.; Britton, G.; Vicario, I.M.; Heredia, F.J. HPLC analysis of geometrical isomers of lutein epoxide isolated from dandelion (Taraxacum officinale F. Weber ex Wiggers). Phytochemistry 2006, 67, 771–777. [Google Scholar] [CrossRef]
- Schweigert, F.J. Metabolism of carotenoids in mammals. In Carotenoids, Volume 3: Biosynthesis and Metabolism, 1st ed.; Britton, G., Pfander, H., Liaaen-Jensen, L.S., Eds.; Birkhäuser: Basel, Switzerland, 1998; pp. 249–284. [Google Scholar]
- Lietz, G.; Oxley, A.; Boesch-Saadatmandi, C.; Kobayashi, D. Importance of β,β-carotene 15,15′-monooxygenase 1 (BCMO1) and β,β-carotene 9′,10′-dioxygenase 2 (BCDO2) in nutrition and health. Mol. Nutr. Food Res. 2012, 56, 241–250. [Google Scholar] [CrossRef]
- Abril, N.; Bárcena, J.A.; Fernández, E.; Galván, A.; Jorrín, J.; Peinado, J.; Toribio, F.; Túnez, I. Espectrofotometría: Espectros de absorción y cuantificación colorimétrica de biomoléculas. Espectrofometría 2000, 1–8, published online. Available online: https://www.uco.es/dptos/bioquimica-biol-mol/pdfs/08_ESPECTROFOTOMETRIA.pdf (accessed on 14 June 2021).
- Dunne, P.G.; Monahan, F.J.; O’Mara, F.P.; Moloney, A.P. Colour of bovine subcutaneous adipose tissue: A review of contributory factors, associations with carcass and meat quality and its potential utility in authentication of dietary history. Meat Sci. 2009, 81, 28–45. [Google Scholar] [CrossRef] [PubMed]
- Arnett, A.M.; Dikeman, M.E.; Spaeth, C.W.; Johnson, B.J.; Hildabrand, B. Effects of vitamin A supplementation in young lambs on performance, serum lipid, and longissimus muscle lipid composition. J. Anim. Sci. 2007, 85, 3062–3071. [Google Scholar] [CrossRef]
- Ripoll, G.; Joy, M.; Muñoz, P.; Albertí, P.; Delfa, R. Fat colour, a traceability parameter of grass feeding in lambs. Options Mediterr. Mediterr. Livest. Prod. Uncertainties Oppor. 2008, 78, 301–306. Available online: https://www.researchgate.net/publication/232724739_Fat_colour_a_traceability_parameter_of_grass_feeding_in_lambs (accessed on 1 June 2021).
- Ripoll, G.; Albertí, P.; Argüello, A.; Alcalde, M.J.; Panea, B. Efecto del sistema de lactancia sobre la calidad de la canal y de la carne de cabra de Guadarrama. In XXXIX Congreso Nacional SEOC; 2014; pp. 255–261. Available online: https://idus.us.es/bitstream/handle/11441/44060/SEOC2014.pdf?sequence=1&isAllowed=y (accessed on 1 June 2021).
- Dian, P.H.M.; Andueza, D.; Barbosa, C.M.P.; Amoureux, S.; Jestin, M.; Carvalho, P.C.F.; Prache, S. Methodological developments in the use of visible reflectance spectroscopy for discriminating pasture-fed from concentrate-fed lamb carcasses. Anim. Int. J. Anim. Biosci. 2007, 1, 1198–1208. [Google Scholar] [CrossRef] [Green Version]
- Ripoll, G.; Casasús, I.; Joy, M.; Molino, F.; Blanco, M. Fat color and reflectance spectra to evaluate the β-carotene, lutein and α-tocopherol in the plasma of bovines finished on meadows or on a dry total mixed ration. Anim. Feed. Sci. Technol. 2015, 207, 20–30. [Google Scholar] [CrossRef]
- Gentili, A.; Caretti, F.; Bellante, S.; Ventura, S.; Canepari, S.; Curini, R. Comprehensive Pro fi ling of Carotenoids and Fat-Soluble Vitamins in Milk from Different Animal Species by LC-DAD-MS/MS Hyphenation. J. Agric. Food Chem. 2013, 61, 1628–1639. [Google Scholar] [CrossRef] [PubMed]
- Vega, F.; Guzmán, J.L.; Delgado-Pertiñez, M.; Zarazaga, L.A.; Argüello, A. Fatty acid composition of muscle and internal fat depots of organic and conventional Payoya goat kids. Span. J. Agric. Res. 2013, 11, 759–769. [Google Scholar] [CrossRef] [Green Version]
- Guzmán, J.L.; Delgado-Pertíñez, M.; Beriáin, M.J.; Pino, R.; Zarazaga, L.Á.; Horcada, A. The use of concentrates rich in orange by-products in goat feed and its effects on physico-chemical, textural, fatty acids, volatile compounds and sensory characteristics of the meat of suckling kids. Animals 2020, 10, 766. [Google Scholar] [CrossRef]
- Sanz-Sampelayo, M.R.; Fernández, J.R.; Ramos, E.; Hermoso, R.; Gil, F.; Boza, J. Effect of providing a polyunsaturated fatty acid-rich protected fat to lactating goats on growth and body composition of suckling goat kids. Anim. Sci. 2006, 82, 337–344. [Google Scholar] [CrossRef]
- Fundación Española Para el Desarrollo de la Nutrición Animal. Concentrados de Proteína Vegetal. Available online: http://www.fundacionfedna.org/concentrados_proteina_vegetal (accessed on 25 April 2021).
- Beratto, E.; Rivas, R. Calidad del grano de avena: Características y parámetros de comercialización. Temuco: Bol. INIA—Inst. Investig. Agropecu. 2002, 87, 9–41. Available online: https://biblioteca.inia.cl/handle/123456789/7785 (accessed on 25 April 2021).
- Bastianelli, D.; Grosjean, F.; Peyronnet, C.; Duparque, M.; Regnier, J.M. Feeding value of pea (Pisum sativum L.) Chemical composition of different categories of pea. Anim. Sci. 1998, 67, 609–619. [Google Scholar] [CrossRef]
- Morrissey, P.A.; Sheehy, P.J.A.; Galvin, K.; Kerryh, J.P.; Buckleyh, D. Lipid Stability in Meat and Meat Products. Meat Sci. 1998, 49, S73–S86. [Google Scholar] [CrossRef]
- Joy, M.; Ripoll, G.; Molino, F.; Dervishi, E.; Álvarez-Rodriguez, J. Influence of the type of forage supplied to ewes in pre- and post-partum periods on the meat fatty acids of suckling lambs. Meat Sci. 2012, 90, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Ripoll, G.; Alcalde, M.J.; Argüello, A.; Córdoba, M.G.; Panea, B. Effect of rearing system on the straight and branched fatty acids of goat milk and meat of suckling kids. Foods 2020, 9, 471. [Google Scholar] [CrossRef] [Green Version]
- Karami, M.; Alimon, A.R.; Sazili, A.Q.; Goh, Y.M.; Ivan, M. Effects of dietary antioxidants on the quality, fatty acid profile, and lipid oxidation of longissimus muscle in Kacang goat with aging time. Meat Sci. 2011, 88, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Osorio, M.T.; Zumalacarregui, J.M.; Cabeza, E.A.; Figueira, A.; Mateo, J. Effect of rearing system on some meat quality traits and volatile compounds of suckling lamb meat. Small Rumin. Res. 2008, 78, 1–12. [Google Scholar] [CrossRef]
- Asadian, A.; Mézes, M. Effects of vitamins A and E supplementation on vitamins A and E status of blood plasma, liver and tail fat of fat-tailed sheep. Small Rumin. Res. 1996, 23, 1–6. [Google Scholar] [CrossRef]
- Canto Guerrero, L. Efecto de las Vitaminas A, D y E Sobre la Expresión del Gen de la Proteína que une al Retinol-4 (RBP-4) en Adipositos 3T3-L1. Universidad Autónoma de Queretaro. Facultad de Medicina. November 2018. Available online: http://ri-ng.uaq.mx/bitstream/123456789/1082/1/ME-0011-Lucero%20Canto%20Guerrero.pdf (accessed on 12 October 2021).
- Liu, Y.; Albrecht, E.; Dannenberger, D.; Hammon, H.M.; Kuehn, C.; Sauerwein, H.; Yang, R.; Zhao, Z.; Maak, S. Retinol binding protein 4 abundance in plasma and tissues is related to body fat deposition in cattle. Sci. Rep. 2019, 9, 8056. [Google Scholar] [CrossRef] [Green Version]
- Debier, C.; Larondelle, Y. Vitamins A and E: Metabolism, roles and transfer to offspring. Br. J. Nutr. 2005, 93, 153–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasapidou, E.; Enser, M.; Wood, J.D.; Richardson, R.I.; Wilkinson, R.G.; Sinclair, L.A. Influence of vitamin E supplementation and basal diet on the vitamin E status, performance and tissue fatty acid concentration in lambs. Animal 2009, 3, 516–526. [Google Scholar] [CrossRef] [Green Version]
- Lobo, G.P.; Amengual, J.; Palczewski, G.; Babino, D.; von Lintig, J. Mammalian Carotenoid-oxygenases: Key players for carotenoid function and homeostasis. Biochim. Biophys. Acta—Mol. Cell Biol. Lipids 2012, 1821, 78–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Analytical Components and Additives | Proportions |
|---|---|
| Crude protein | 23.50% |
| Oils and Fat | 26.00% |
| Crude Fiber | 0.10% |
| Raw ash | 7.00% |
| Calcium | 0.90% |
| Phosphorus | 0.70% |
| Sodium | 0.40% |
| Vitamin A (E-672) | 80,000 UI/Kg |
| Vitamin D3 (E-671) | 4,250 UI/Kg |
| Vitamin E (all-rac-alpha-tocopherol acetate) | 30 UI/Kg |
| Iron (E1) (Sulfate monohydrate) | 40 mg/Kg |
| Iodine (E2) (Calcium iodate anhydrous) | 0.15 mg/Kg |
| Cobalt (E3) (Basic carbonate monohydrate) | 0.20 mg/Kg |
| Copper (E4) (Sulfate Pentahydrate) | 5 mg/kg |
| Magnesium (E5) (Sulfate monohydrate) | 25 mg/Kg |
| Zinc (E6) (Oxide) | 30 mg/Kg |
| Selenium (3b8.12) (Selenomethionine) | 0.20 mg/Kg |
| Antioxidant (B.H.T.) (E-321) | 100 mg/Kg |
| Preservative (Potassium Sorbate) (E-202) | |
| Emulsifiers (Lecithins) (E-322) |
| Natural Milk | Milk Replacer | ||||
|---|---|---|---|---|---|
| MG | CM | TMR | MR | p-Value | |
| SA (days) | 36.75 c ± 0.27 | 31.92 b ± 0.69 | 30.07 a ± 0.44 | 47.44 d ± 0.27 | <0.001 |
| SLW (kg) | 8.53 ± 0.28 | 8.01 ± 0.22 | 7.77 ± 0.31 | 8.38 ± 0.25 | 0.187 |
| HCW (kg) | 4.97 b ± 0.18 | 4.30 a ± 0.15 | 5.26 b ± 0.16 | 4.95 b ± 0.16 | 0.002 |
| KFW (g) | 53.92 a ± 3.64 | 97.85 b ± 10.99 | 48.57 a ± 5.43 | 37.50 a ± 5.66 | <0.001 |
| Kidney fat color | |||||
| L* | 76.79 ± 1.11 | 73.50 ± 1.49 | 73.24 ± 1.38 | 72.77 ± 1.45 | 0.198 |
| a* | 2.83 ab ± 0.39 | 1.62 a ± 0.33 | 2.24 ab ± 0.38 | 2.96 b ± 0.31 | 0.036 |
| b* | 13.35 b ± 0.85 | 11.13 a ± 0.48 | 12.96 ab ± 0.41 | 13.03 ab ± 0.48 | 0.037 |
| C* | 13.68 b ± 0.89 | 11.30 a ± 0.50 | 13.20 ab ± 0.46 | 13.39 ab ± 0.53 | 0.035 |
| h° | 78.44 ± 1.26 | 82.03 ± 1.41 | 80.59 ± 1.34 | 77.57 ± 0.93 | 0.051 |
| SUM | 195.52 b ±18.05 | 159.06 ab ± 14.05 | 121.40 a ± 14.27 | 175.29 ab ± 12.40 | 0.007 |
| Natural Milk | Milk Replacer | ||||
|---|---|---|---|---|---|
| MG | CM | TMR | MR | p-Value | |
| % LTF | 1.13 a ± 0.13 | 4.90 b ± 0.48 | 4.81 b ± 0.53 | 1.63 a ± 0.21 | <0.001 |
| MDA | 0.089 a ± 0.011 | 0.270 b ± 0.025 | 0.252 b ± 0.039 | 0.086 a ± 0.017 | <0.001 |
| Fat Retinol | 26.81 b ± 1.30 | 16.52 a ± 1.63 | 15.99 a ± 1.31 | 22.63 b ± 144 | <0.001 |
| Plasma Retinol | 5.56 a ± 0.96 | 3.72 a ± 0.73 | 14.98 b ± 2.748 | 22.47 c ± 1.91 | <0.001 |
| Plasma Tocopherol | 11.43 c ± 2.50 | 8.85 bc ± 2.63 | 2.49 ab ± 0.61 | 0.52 a ± 0.16 | <0.001 |
| Discriminant | Predicted/Actual Membership | MG | CM | TMR | MR |
|---|---|---|---|---|---|
| (1) | MG | 66.7 | 0 | 0 | 33.3 |
| CM | 0 | 69.2 | 15.4 | 15.4 | |
| TMR | 0 | 7.1 | 64.3 | 28.6 | |
| MR | 31.25 | 0 | 0 | 68.75 | |
| (2) | MG | 83.3 | 16.7 | 0 | 0 |
| CM | 23.1 | 76.9 | 0 | 0 | |
| TMR | 0 | 28.6 | 42.9 | 28.6 | |
| MR | 0 | 6.7 | 6.7 | 86.7 | |
| (3) | MG | 100 | 0 | 0 | 0 |
| CM | 7.7 | 92.3 | 0 | 0 | |
| TMR | 0 | 0 | 92.9 | 7.1 | |
| MR | 0 | 6.7 | 0 | 93.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roncero-Díaz, M.; Panea, B.; Córdoba, M.d.G.; Argüello, A.; Alcalde, M.J. Retinol and α-Tocopherol Contents, Fat Color, and Lipid Oxidation as Traceability Tools of the Feeding System in Suckling Payoya Kids. Animals 2022, 12, 104. https://doi.org/10.3390/ani12010104
Roncero-Díaz M, Panea B, Córdoba MdG, Argüello A, Alcalde MJ. Retinol and α-Tocopherol Contents, Fat Color, and Lipid Oxidation as Traceability Tools of the Feeding System in Suckling Payoya Kids. Animals. 2022; 12(1):104. https://doi.org/10.3390/ani12010104
Chicago/Turabian StyleRoncero-Díaz, Mercedes, Begoña Panea, María de Guía Córdoba, Anastasio Argüello, and María J. Alcalde. 2022. "Retinol and α-Tocopherol Contents, Fat Color, and Lipid Oxidation as Traceability Tools of the Feeding System in Suckling Payoya Kids" Animals 12, no. 1: 104. https://doi.org/10.3390/ani12010104