
Behavioral Changes of Solitary Housed Female Pygmy Slow Lorises (Nycticebus pygmeaus) after Introduction into Group Enclosures



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Study Subject Testing Environment
2.3. Data Collection
2.4. Statistical Analyses
3. Results
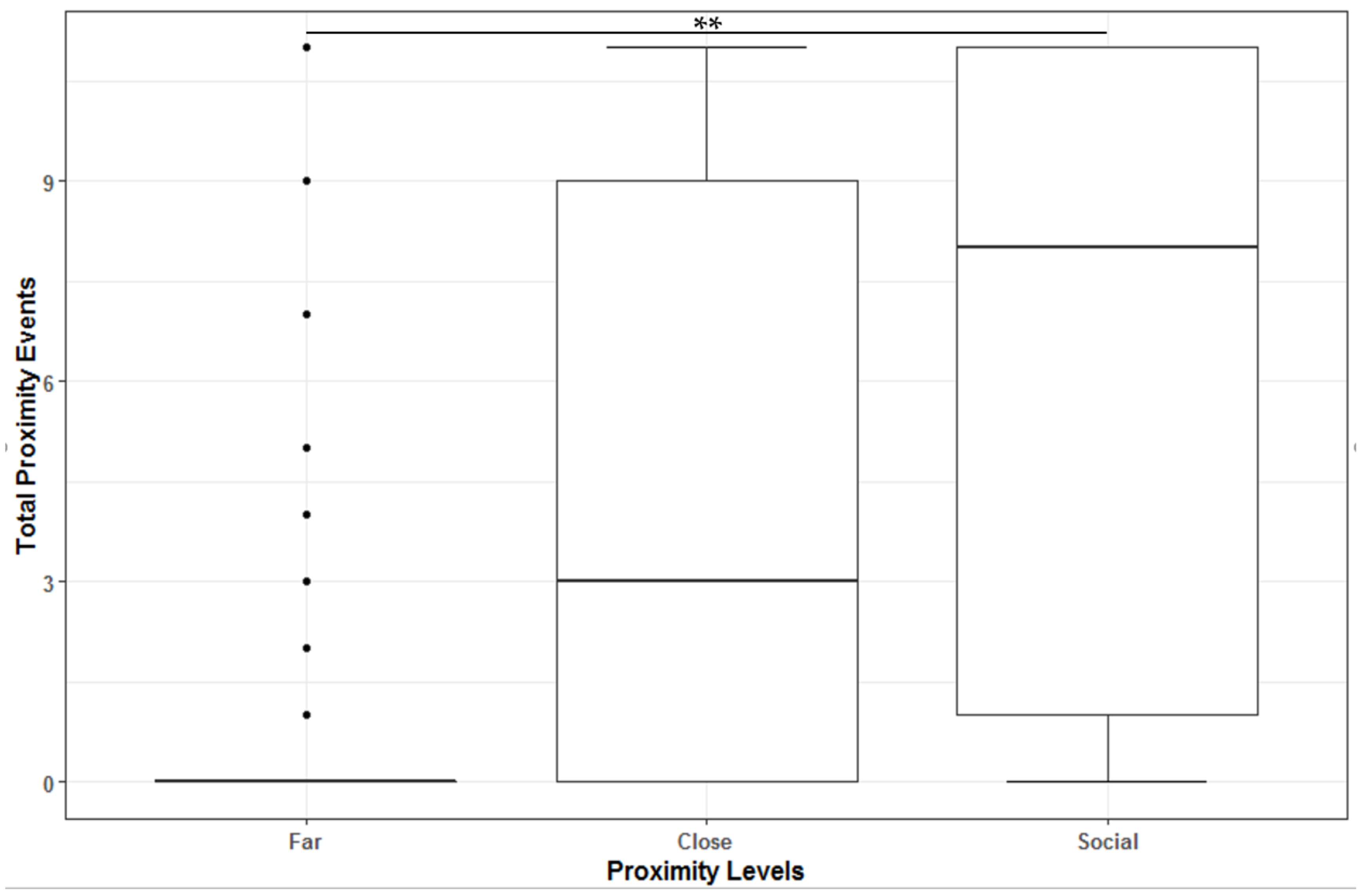
3.1. Proximity
3.2. Activity Budgets
3.3. Positional Behaviors
4. Discussion
4.1. Sociality among Female Pygmy Slow Lorises
4.2. Activity Budget
4.3. Postural and Locomotor Behaviors
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Starr, C.; Nekaris, K.; Streicher, U.; Leung, L. Field Surveys of the Vulnerable Pygmy Slow Loris Nycticebus pygmaeus using Local Knowledge in Mondulkiri Province, Cambodia. Oryx 2011, 45, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Nekaris, K.; Starr, C.R. Conservation and Ecology of the Neglected Slow Loris: Priorities and Prospects. Endanger. Species Res. 2015, 28, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Blair, M.; Nadler, T.; Ni, O.; Samun, E.; Streicher, U.; Nekaris, K. Nycticebus pygmaeus. The IUCN Red List of Threatened Species. 2020. Available online: https://dx.doi.org/10.2305/IUCN.UK.2020-2.RLTS.T14941A17971417.en (accessed on 25 August 2021).
- Musing, L.; Suzuki, K.; Nekaris, K.A.I. Crossing International Borders: The Trade of Slow Lorises (Nycticebus spp.) as Pets in Japan. Asian Primates J. 2016, 5, 12–23. [Google Scholar]
- Yamanashi, Y.; Nemoto, K.; Alejandro, J. Social Relationships Among Captive Male Pygmy Slow Lorises (Nycticebus pygmaeus): Is Forming Male Same-Sex Pairs a Feasible Management Strategy? Am. J. Primatol. 2021, 83, e23233. [Google Scholar] [CrossRef]
- Petter, J.J.; Hladik, C.M. Obeservations sur le Domaine Vital et la Densite de Population de Loris Tardigradus dans les Forets de Ceylan. Mammalia 1970, 34, 394–409. [Google Scholar] [CrossRef]
- Barrett, E. The Ecology of Some Nocturnal Arboreal Mammals in the Rain Forest of Peninsular Malaysia. Ph.D. Thesis, Department of Physical Anthropology, The University of Cambridge, Cambridge, UK, 1985. [Google Scholar]
- Bearder, S.K. Lorises, bushbabies, and tarsiers: Diverse Societies in Solitary Foragers. In Primate Societies; Smuts, B.B., Cheney, D.L., Seyfarth, R.M., Wrangham, R.W., Struhsaker, T.T., Eds.; University of Chicago Press: Chicago, IL, USA, 1987; pp. 11–24. [Google Scholar]
- Nekaris, K.A.I. Activity Budget and Positional Behavior of the Mysore Slender Loris (Loris tardigradus lydekkerianus): Implications for Slow Climbing Locomotion. Folia Primatol. 2001, 72, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Nekaris, K.A.I.; Moore, R.S.; Rode, E.J. Mad, Bad and Dangerous to Know: The Biochemistry, Ecology and Evolution of Slow Loris Venom. J. Venom Anim. Toxins Incl. Trop. Dis. 2013, 19, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bearder, S.K.; Martin, R.D. The Social Organization of a Nocturnal Prosimian Revealed by Radio-Tracking. In A Handbook on Biotelemetry and Radio-Tracking; Anamler, C.J., Macdonald, D.W., Eds.; Pergamon Press: Oxford, UK, 1980; pp. 633–648. [Google Scholar]
- Pimley, E.R.; Bearder, S.K.; Dixson, A.F. Home Range Analysis of Perodicticus potto edwardsi and Sciurocheirus cameronensis. Int. J. Primatol. 2005, 26, 191. [Google Scholar] [CrossRef]
- Radhakrishna, S. Midnight’s Children? Solitary Primates and Gregarious Chiropterans. Curr. Sci. India. 2005, 89, 1208–1213. Available online: https://www.jstor.org/stable/24110973 (accessed on 5 May 2021).
- Nekaris, K.A.I. Social Lives of Adult Mysore Slender Lorises (Loris lydekkerianus lydekkerianus). Am. J. Primatol. 2006, 68, 1171–1182. [Google Scholar] [CrossRef]
- Wiens, F. Behavior and Ecology of Wild Slow Lorises (Nycticebus coucang): Social Organization, Infant Care System, and Diet. Ph.D. Thesis, Faculty of Biology, Chemistry and Earth Sciences, University of Bayreuth, Bayreuth, Germany, 2002. Available online: https://epub.uni-bayreuth.de/1024/1/Diss.pdf (accessed on 5 May 2021).
- Goonan, P.M. Behaviour and Reproduction of the Slender Loris (Loris tardigradus) in Captivity. Folia Primatol. 1993, 60, 146–157. [Google Scholar] [CrossRef]
- Starr, C.; Nekaris, K. Ranging Patterns of the Pygmy Slow Loris (Nycticebus pygmaeus) in a Mixed Deciduous Forest in Eastern Cambodia. In Evolution, Ecology and Conservation of Lorises and Pottos; Nekaris, K., Burrows, A., Eds.; Cambridge University Press, Cambridge Studies in Biological and Evolutionary Anthropology: Cambridge, UK, 2020; pp. 228–234. [Google Scholar] [CrossRef]
- Poindexter, S.; Khoa, D.; Nekaris, K.A.I. The Ranging Patterns of Reintroduced Pygmy Slow Lorises (Nycticebus pygmaeus) in Cuc Phuong National Park, Vietnam. Viet. J. Primatol. 2017, 22, 37–49. [Google Scholar]
- Fisher, H.S.; Swaisgood, R.R.; Fitch-Snyder, H. Countermarking by Male Pygmy Lorises (Nycticebus Pygmaeus): Do Females Use Odor Cues to Select Mates with High Competitive Ability? Behav. Ecol. Sociobiol. 2003, 53, 123–130. [Google Scholar] [CrossRef]
- Fernández Lázaro, G.; Zehr, S.; Alonso García, E. Use of Primates in Research: What Do We Know About Captive Strepsirrhine Primates? J. Appl. Anim. Welf. Sci. 2017, 20, 109–122. [Google Scholar] [CrossRef]
- Agnani, P.; Kauffmann, C.; Hayes, L.D.; Schradin, C. Intra-Specific Variation in Social Organization of Strepsirrhines. Am. J. Primatol. 2018, 80, e22758. [Google Scholar] [CrossRef]
- Blanco, M.B.; Dausmann, K.H.; Faherty, S.L.; Yoder, A.D. Tropical Heterothermy is “Cool”: The Expression of Daily Torpor and Hibernation in Primates. Evol. Anthropol. 2018, 27, 147–161. [Google Scholar] [CrossRef]
- Muchlinski, M.N.; Perry, J.M. Anatomical Correlates to Nectar Feeding among the Strepsirrhines of Madagascar: Implications for Interpreting the Fossil Record. Anat. Res. Int. 2011, 2011, 378431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmen, B.; Morino, L.; Blanc, S.; Garcia, C. The Energy Allocation Trade-offs Underlying Life History Traits in Hypometabolic Strepsirrhines and other Primates. Sci. Rep. 2021, 11, 14196. [Google Scholar] [CrossRef] [PubMed]
- Price, E.E.; Stoinski, T.S. Group size: Determinants in the Wild and Implications for the Captive Housing of Wild Mammals in Zoos. Appl. Anim. Behav. Sci. 2007, 103, 255–264. [Google Scholar] [CrossRef]
- Hearn, G.W.; Berghaier, R.W.; George, D.D. Evidence for social enhancement of reproduction in two Eulemur species. Zoo Biol. 1996, 15, 1–12. [Google Scholar] [CrossRef]
- Ehrlich, A. Social and Individual Behaviors in Captive Greater Galagos. Behaviour 1977, 63, 192–214. [Google Scholar] [CrossRef]
- Bercovitch, F.B. A Sequential Analysis of Female Aggression in a Group of Lesser Galagos (Galago senegalensis). Primates 1978, 19, 505–516. [Google Scholar] [CrossRef]
- Nash, L.; Flinn, L. Group Formation in Captive Lesser Galagos (Galago senegalensis). Primates 1978, 19, 493–503. [Google Scholar] [CrossRef]
- Ehrlich, A.; Musicant, A. Social and Individual Behaviors in Captive Slow Lorises. Behaviour 1977, 60, 195–220. [Google Scholar] [CrossRef] [Green Version]
- The Duke Lemur Center, Pygmy Slow Loris (Nycticebus pygmaeus). Available online: https://lemur.duke.edu/discover/meet-the-lemurs/pygmy-slow-loris/ (accessed on 10 September 2021).
- Schulze, H.; Meier, B. Behavior of Captive Loris Tardigradus Nordicus: A Qualitative Description, Including Some Information about Morphological Bases of Behavior. In Creatures of the Dark; Alterman, L., Doyle, G.A., Izard, M.K., Eds.; Springer: Boston, MA, USA, 1995. [Google Scholar] [CrossRef]
- Putri, P.R.; Moore, R.S.; Andayani, N.; Sánchez, K.L. Effects of Environmental Enrichment on the Social Behaviour of Javan Slow Loris, Nycticebus javanicus. TAPROBANICA J. Asian Biodivers. 2015, 7, 29. [Google Scholar] [CrossRef] [Green Version]
- Fitch-Snyder, H.; Schulze, H. Management of Lorises in Captivity: A Husbandry Manual for Asian Lorisines (Nycticebus and Loris spp.); Zoological Society of San Diego: San Diego, CA, USA, 2001. [Google Scholar]
- Fuller, G.; Kuhar, C.W.; Dennis, P.M.; Lukas, K.E. A Survey of Husbandry Practices for Lorisid Primates in North American Zoos and Related Facilities. Zoo Biol. 2013, 32, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Sellers, W.A. Biomechanical Investigation into the Absence of Leaping in the Locomotor Repertoire of the Slender Loris (Loris tardigradus). Folia Primatol. 1996, 67, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, N.; Nekaris, K.A.I. Positional Behaviour and Substrate Preference of Slow Lorises, with a Case Study of Nycticebus bengalensis in Northeast India. In Evolution, Ecology and Conservation of Lorises and Pottos; Nekaris, K., Burrows, A., Eds.; Cambridge University Press, Cambridge Studies in Biological and Evolutionary Anthropology: Cambridge, UK, 2020; pp. 210–218. [Google Scholar] [CrossRef]
- Fuller, G.; Lukas, K.E.; Kuhar, C.; Dennis, P.M. A Retrospective Review of Mortality in Lorises and Pottos in North American Zoos, 1980–2010. Endanger. Species Res. 2014, 23, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Glassman, D.M.; Wells, J.P. Positional and Activity Behavior in a Captive Slow Loris: A Quantitative Assessment. Am. J. Primatol. 1984, 7, 121–132. [Google Scholar] [CrossRef]
- Jaman, M.F.; Huffman, M.A. Enclosure Environment Affects the Activity Budgets of Captive Japanese Macaques (Macaca fuscata). Am. J. Primatol. 2008, 70, 1133–1144. [Google Scholar] [CrossRef]
- Howell, C.; Cheyne, S. Complexities of Using Wild versus Captive Activity Budget Comparisons for Assessing Captive Primate Welfare. J. Appl. Anim. Welf. Sci. 2018, 22, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Cabana, F.; Plowman, A. Pygmy Slow Loris Nycticebus pygmaeus—Natural Diet Replication in Captivity. Endanger. Species Res. 2014, 23, 197–204. [Google Scholar] [CrossRef] [Green Version]
- Fuller, G. Validating Actigraphy for Circadian Monitoring of Behavior in the Pygmy Loris (Nycticebus pygmaeus) and Potto (Perodicticus potto); In the Night Shift: Lighting and Nocturnal Strepsirrhine Care in Zoos. Ph.D. Thesis, Case Reserve Western University, Cleveland, OH, USA, 2014. Available online: https://etd.ohiolink.edu/apexprod/rws_etd/send_file/send?accession=case1384463090&disposition=inline (accessed on 5 May 2021).
- Al-Razi, H.; Hasan, S.; Ahmed, T.; Muzaffar, S.B. Home Range, Activity Budgets and Habitat Use in the Bengal Slow Loris (Nycticebus bengalensis) in Bangladesh. In Evolution, Ecology and Conservation of Lorises and Pottos; Nekaris, K., Burrows, A., Eds.; Cambridge University Press, Cambridge Studies in Biological and Evolutionary Anthropology: Cambridge, UK, 2020; pp. 193–203. [Google Scholar] [CrossRef]
- Reinhardt, V.; Cowley, D.; Eisele, S.; Scheffler, J. Serum Cortisol Concentrations of Single-Housed and Isosexually Pair-Housed Adult Rhesus Macaques. J. Exp. Anim. Sci. 1991, 34, 73–76. [Google Scholar] [PubMed]
- Baker, K.C.; Bloomsmith, M.A.; Oettinger, B. Benefits of Pair Housing are Consistent Across a Diverse Population of Rhesus Macaques. Appl. Anim. Behav. Sci. 2012, 137, 148–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitch-Snyder, H. Husbandry and Reproductive Management Recommendations for Captive Lorises and Pottos (Nycticebus, Loris and Perodicticus). In Evolution, Ecology and Conservation of Lorises and Pottos; Nekaris, K., Burrows, A., Eds.; Cambridge University Press, Cambridge Studies in Biological and Evolutionary Anthropology: Cambridge, UK, 2020; pp. 263–276. [Google Scholar] [CrossRef]
- Starr, C.; Nekaris, K.A.I.; Leung, L. Hiding from the Moonlight: Luminosity and Temperature Affect Activity of Asian Nocturnal Primates in a Highly Seasonal Forest. PLoS ONE 2012, 7, e36396. [Google Scholar] [CrossRef]
- Ariana, W.; Marco, C.; Nekaris, K.A.I. Red Light for Nocturnal Observations. In Evolution, Ecology and Conservation of Lorises and Pottos; Nekaris, K., Burrows, A., Eds.; Cambridge University Press, Cambridge Studies in Biological and Evolutionary Anthropology: Cambridge, UK, 2020; pp. 492–494. [Google Scholar]
- Cabana, F.; Nekaris, K. Diets High in Fruits and Low in Gum Exudates Promote the Occurrence and Development of Dental Disease in Pygmy Slow Loris (Nycticebus pygmaeus). Zoo Biol. 2015, 34, 547–553. [Google Scholar] [CrossRef]
- Martin, P.; Bateson, P.P.G. Measuring Behaviour: An Introductory Guide; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Chinn, R.N. Communication and Social Behavior of Captive Slow Lorises (Nycticebus coucang). Master’s Thesis, University of the Pacific, Stockton, CA, USA, 1980. Available online: https://scholarlycommons.pacific.edu/uop_etds/2039 (accessed on 6 May 2021).
- Cassidy, L.C.; Hannibal, D.L.; Semple, S.; McCowan, B. Improved Behavioral Indices of Welfare in Continuous Compared to Intermittent Pair-Housing in Adult Female Rhesus Macaques (Macaca mulatta). Am. J. Primatol. 2020, 82, e23189. [Google Scholar] [CrossRef]
- Cheney, D.L. Intragroup Cohesion and Intergroup Hostility: The Relation Between Grooming Distributions and Intergroup Competition among Female Primates. Behav. Ecol. 1992, 4, 334–345. [Google Scholar] [CrossRef]
- Seyfarth, R.M. A Model of Social Grooming among Adult Female Monkeys. J. Theor. Biol. 1977, 21, 671–698. [Google Scholar] [CrossRef] [PubMed]
- Chapais, B.; Girard, M.; Primi, G. Non-kin Alliances and the Stability of Matrilineal Dominance Relations in Japanese Macaques. Anim. Behav. 1991, 41, 481–491. [Google Scholar] [CrossRef]
- Maestripieri, D.; Schino, G.; Aureli, F.; Troisi, A. A Modest Proposal—Displacement Activities as an Indicator of Emotions in Primates. Anim. Behav. 1992, 44, 967–979. [Google Scholar] [CrossRef]
- Hagey, L.R.; Fry, B.G.; Fitch-Snyder, H. Talking Defensively, a Dual Use for the Brachial Gland Exudate of Slow and Pygmy Lorises. In Primate Anti-Predator Strategies. Developments in Primatology: Progress and Prospects; Gursky, S.L., Nekaris, K.A.I., Eds.; Springer: Boston, MA, USA, 2007; pp. 253–272. [Google Scholar] [CrossRef]
- Duncan, S.D. Behavioral Variability in Captive Slow Lorises, Nycticebus coucang (Lorisidae, Primates). Master’s Thesis, University of the Pacific, Stockton, CA, USA, 1982. Available online: https://scholarlycommons.pacific.edu/uop_etds/2071 (accessed on 6 May 2021).
- Tenaza, R.; Ross, B.A.; Tanticharoenyos, P.; Berkson, G. Individual Behavior and Activity Rhythms of Captive Slow Lorises (Nycticebus coucang). Anim. Behav. 1969, 17, 664–669. [Google Scholar] [CrossRef]
- Williams, E.; Cabana, F.; Nekaris, K.A.I. Improving Diet and Activity of Insectivorous Primates in Captivity: Naturalizing the Diet of Northern Ceylon Gray Slender Loris, Loris lydekkerianus nordicus. Zoo Biol. 2015, 34, 473–482. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Behaviors | Description |
|---|---|
| Rest | The body is immobile and not engaging in any activity |
| Feed | Consuming any animal or provisioned food or mastication |
| Move | Any mobile activity in any direction |
| Self-Groom | Licking or combing with teeth its coat |
| Social-Groom | Actively licking or combing with teeth a conspecific’s coat or receiving the same |
| Postural Categories | Posture | Description |
|---|---|---|
| Locomotion Posture 1 | Climb up | climbing up using all limbs |
| Climb down | climbing down using all limbs | |
| Quadrupedal walk Cling | walking on all four limbs hanging with more than two limbs | |
| Locomotion Posture 2 | Quadrupedal stand | standing with all four legs |
| Bipedal stand Bipedal hang | standing with hind legs hanging face down with hind legs | |
| Positional Posture 3 | Sit Sleep ball | sitting or lying down curled up in a ball |
| Positional Posture 4 | Bridge | extending all limbs to grab or move between two substrates |
| Other | unclear posture or transitional position |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alejandro, J.; Yamanashi, Y.; Nemoto, K.; Bercovitch, F.B.; Huffman, M.A. Behavioral Changes of Solitary Housed Female Pygmy Slow Lorises (Nycticebus pygmeaus) after Introduction into Group Enclosures. Animals 2021, 11, 2751. https://doi.org/10.3390/ani11092751
Alejandro J, Yamanashi Y, Nemoto K, Bercovitch FB, Huffman MA. Behavioral Changes of Solitary Housed Female Pygmy Slow Lorises (Nycticebus pygmeaus) after Introduction into Group Enclosures. Animals. 2021; 11(9):2751. https://doi.org/10.3390/ani11092751
Chicago/Turabian StyleAlejandro, Josue, Yumi Yamanashi, Kei Nemoto, Fred B. Bercovitch, and Michael A. Huffman. 2021. "Behavioral Changes of Solitary Housed Female Pygmy Slow Lorises (Nycticebus pygmeaus) after Introduction into Group Enclosures" Animals 11, no. 9: 2751. https://doi.org/10.3390/ani11092751