
Efflux Might Participate in Decreased Susceptibility to Oxytetracycline in Contagious Agalactia-Causative Mycoplasma spp.

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mycoplasma Isolates
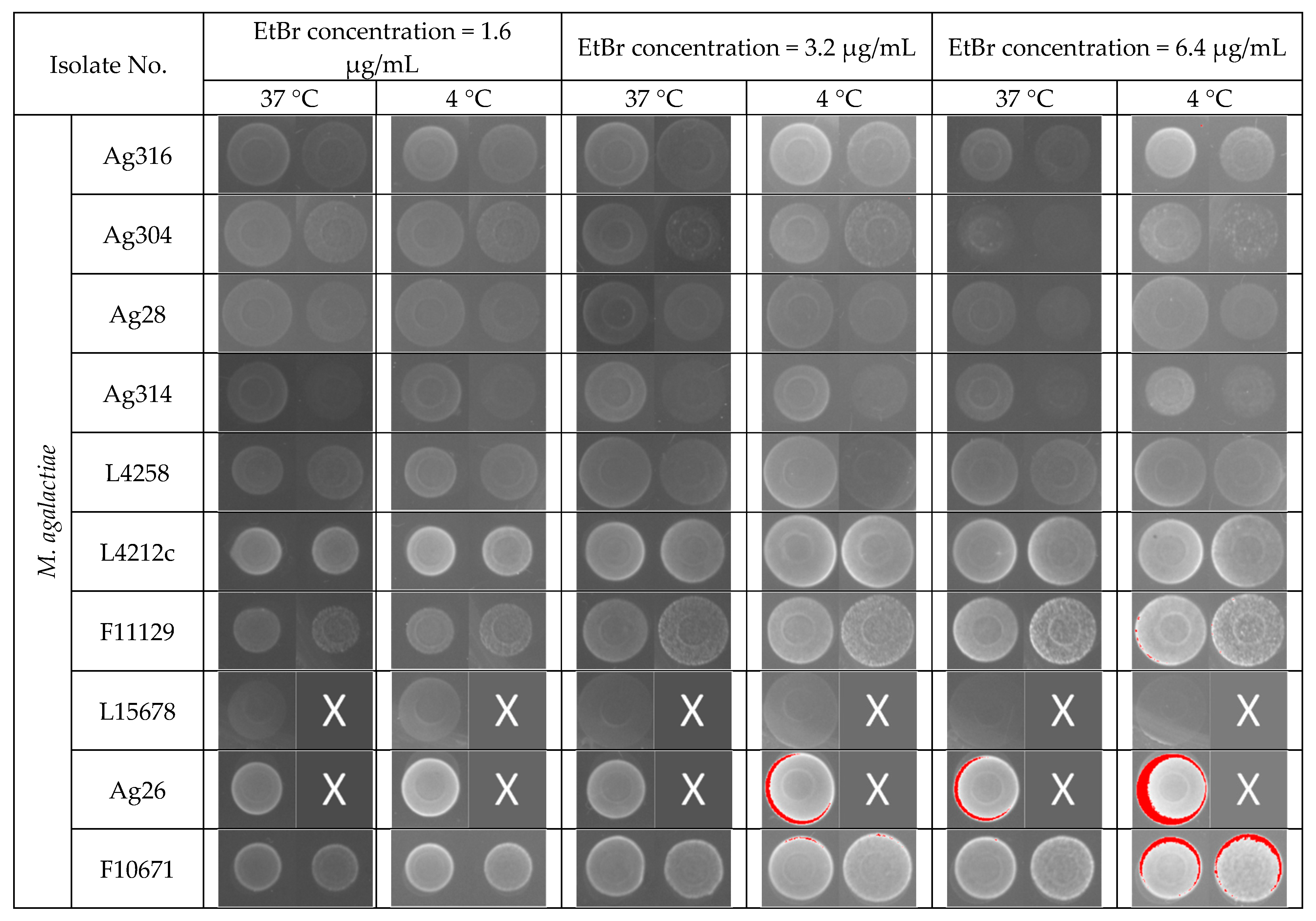
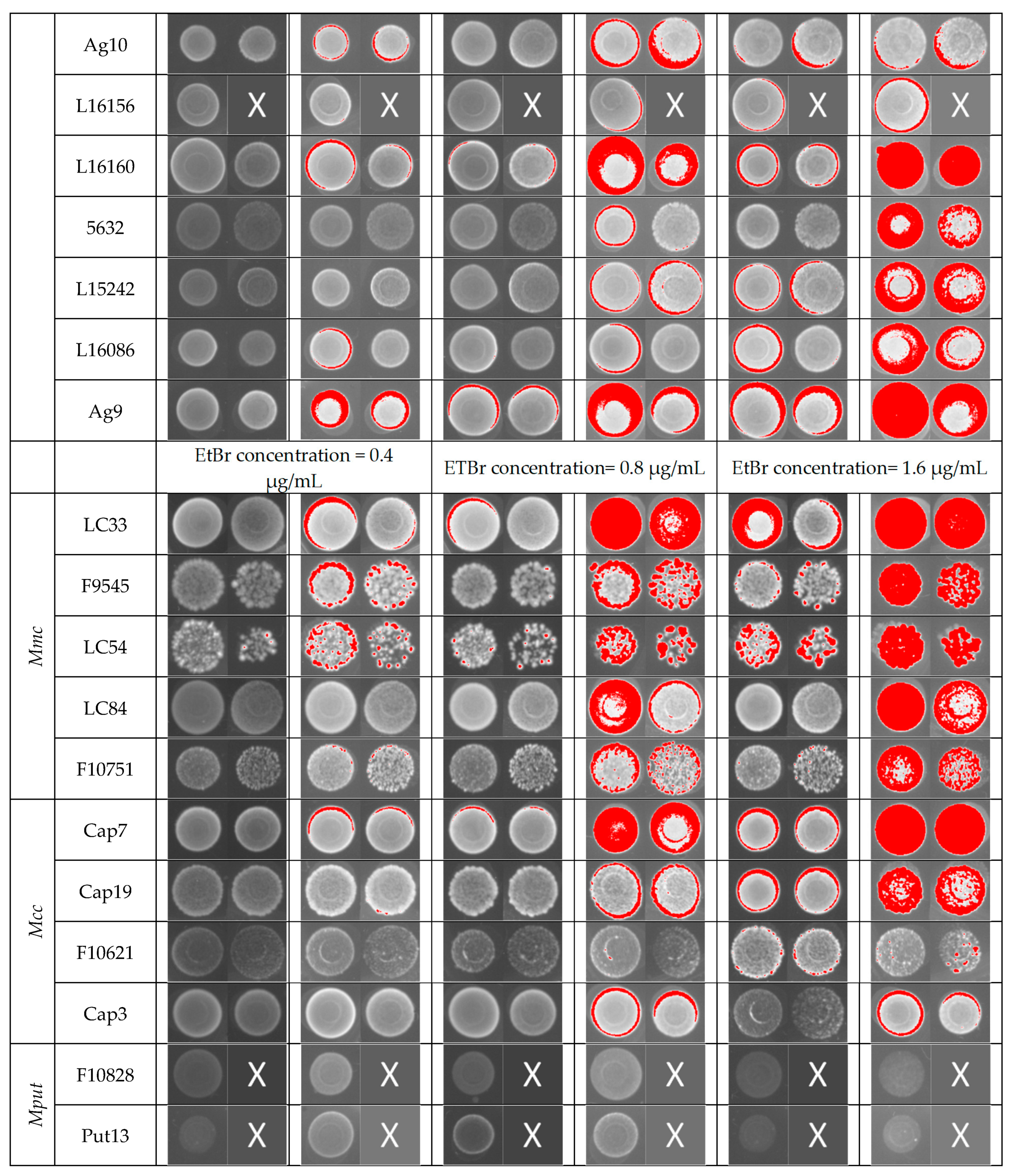
2.2. Efflux Assay by EtBr (EtBr)–Agar Method
2.3. Monitoring of EtBr Efflux by Fluorometry
2.4. Growth in the Presence of Subinhibitory Concentrations of Oxytetracycline
3. Results
3.1. Efflux Assay by EtBr (EtBr)–Agar Method
3.2. Monitoring of EtBr Efflux by Fluorometry
3.3. Growth in the Presence of Subinhibitory Concentrations of Oxytetracycline
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jaÿ, M.; Tardy, F. Contagious Agalactia In Sheep and Goats: Current Perspectives. Vet. Med. Res. Rep. 2019, 10, 229–247. [Google Scholar] [CrossRef] [PubMed]
- Corrales, J.; Esnal, A.; De la Fe, C.; Sánchez, A.; Assunçao, P.; Poveda, J.; Contreras, A. Contagious agalactia in small ruminants. Small Rumin. Res. 2007, 68, 154–166. [Google Scholar] [CrossRef]
- Ariza-Miguel, J.; Rodríguez-Lázaro, D.; Hernández, M. A survey of Mycoplasma agalactiae in dairy sheep farms in Spain. BMC Vet. Res. 2012, 8, 171. [Google Scholar] [CrossRef] [Green Version]
- Manso-Silván, L.; Perrier, X.; Thiaucourt, F. Phylogeny of the Mycoplasma mycoides cluster based on analysis of five conserved protein-coding sequences and possible implications for the taxonomy of the group. Int. J. Syst. Evol. Microbiol. 2007, 57, 2247–2258. [Google Scholar] [CrossRef] [Green Version]
- Poumarat, F.; Jarrige, N.; Tardy, F. Purpose and overview of results of the Vigimyc network for the epidemiological surveillance of mycoplasmoses in ruminants in France. EuroRéférence 2014, 12, 24–28. [Google Scholar]
- Jaÿ, M.; Poumarat, F.; Colin, A.; Tricot, A.; Tardy, F. Addressing the antimicrobial resistance of ruminant mycoplasmas using a clinical surveillance network. Front. Vet. Sci. 2021, 8, 667175. [Google Scholar] [CrossRef]
- Agnone, A.; La Manna, M.; Sireci, G.; Puleio, R.; Usticano, A.; Ozdemir, U.; Nicholas, R.A.; Chiaracane, V.; Dieli, F.; Di Marco, V.; et al. A comparison of the efficacy of commercial and experimental vaccines for contagious agalactia in sheep. Small Rumin. Res. 2013, 112, 230–234. [Google Scholar] [CrossRef]
- Gómez-Martín, Á.; Amores, J.; Paterna, A.; de la Fe, C. Contagious agalactia due to Mycoplasma spp. in small dairy ruminants: Epidemiology and prospects for diagnosis and control. Vet. J. 2013, 198, 48–56. [Google Scholar] [CrossRef] [PubMed]
- De Briyne, N.; Atkinson, J.; Borriello, S.P.; Pokludová, L. Antibiotics used most commonly to treat animals in Europe. Vet. Rec. 2014, 175, 325. [Google Scholar] [CrossRef] [Green Version]
- Prats-van der Ham, M.; Tatay-Dualde, J.; Ambroset, C.; De la Fe, C.; Tardy, F. The moderate drift towards less tetracycline-susceptible isolates of contagious agalactia causative agents might result from different molecular mechanisms. Vet. Microbiol. 2018, 220, 39–46. [Google Scholar] [CrossRef]
- Antunes, N.; Tavío, M.; Assunção, P.; Rosales, R.; Aquili, V.; de la Fe, C.; Poveda, J. In vitro susceptibilities of field isolates of Mycoplasma mycoides subsp. mycoides large colony type to 15 antimicrobials. Vet. Microbiol. 2007, 119, 72–75. [Google Scholar] [CrossRef]
- Paterna, A.; Tatay-Dualde, J.; Amores, J.; der Ham, M.P.-V.; Sánchez, A.; de la Fe, C.; Contreras, A.; Corrales, J.; Gómez-Martín, Á. In vitro assessment of the antimicrobial susceptibility of caprine isolates of Mycoplasma mycoides subsp. capri. Vet. J. 2016, 214, 96–101. [Google Scholar] [CrossRef]
- Poumarat, F.; Gautier-Bouchardon, A.; Bergonier, D.; Gay, E.; Tardy, F. Diversity and variation in antimicrobial susceptibility patterns over time in Mycoplasma agalactiae isolates collected from sheep and goats in France. J. Appl. Microbiol. 2016, 120, 1208–1218. [Google Scholar] [CrossRef] [Green Version]
- Tatay-Dualde, J.; Prats-van der Ham, M.; de La Fe, C.; Paterna, A.; Sánchez, A.; Corrales, J.C.; Contreras, A.; Tola, S.; Gómez-Martin, Á. Antimicrobial susceptibility and multilocus sequence typing of Mycoplasma capricolum subsp. capricolum. PLoS ONE 2017, 12, e0174700. [Google Scholar] [CrossRef] [Green Version]
- Grossman, T.H. Tetracycline Antibiotics and Resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025387. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, F.; Starosta, A.; Arenz, S.; Sohmen, D.; Dönhöfer, A.; Wilson, D. Tetracycline antibiotics and resistance mechanisms. Biol. Chem. 2014, 395, 559–575. [Google Scholar] [CrossRef] [PubMed]
- Meygret, A.; Le Roy, C.; Renaudin, H.; Bébéar, C.; Pereyre, S. Tetracycline and fluoroquinolone resistance in clinical Ureaplasma spp. and Mycoplasma hominis isolates in France between 2010 and 2015. J. Antimicrob. Chemother. 2018, 73, 2696–2703. [Google Scholar] [CrossRef]
- Dégrange, S.; Renaudin, H.; Charron, A.; Pereyre, S.; Bebear, C. Reduced susceptibility to tetracyclines is associated in vitro with the presence of 16S rRNA mutations in Mycoplasma hominis and Mycoplasma pneumoniae. J. Antimicrob. Chemother. 2008, 61, 1390–1392. [Google Scholar] [CrossRef] [PubMed]
- Waites, K.; Lysnyanski, I.; Bébéar, C. Emerging antimicrobial resistance in mycoplasmas of humans and animals. In Mollicutes: Molecular Biology and Pathogenesis; Browning, G.F., Citti, C., Eds.; Caister Academic Press: Norfolk, UK, 2014; pp. 289–322. [Google Scholar]
- Gautier-Bouchardon, A.V. Antimicrobial Resistance in Mycoplasma spp. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Amram, E.; Mikula, I.; Schnee, C.; Ayling, R.D.; Nicholas, R.A.J.; Rosales, R.S.; Harrus, S.; Lysnyansky, I. 16S rRNA Gene Mutations Associated with Decreased Susceptibility to Tetracycline in Mycoplasma bovis. Antimicrob. Agents Chemother. 2014, 59, 796–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalil, D.; Becker, C.A.; Tardy, F. Monitoring the Decrease in Susceptibility to Ribosomal RNAs Targeting Antimicrobials and Its Molecular Basis in Clinical Mycoplasma bovis Isolates over Time. Microb. Drug Resist. 2017, 23, 799–811. [Google Scholar] [CrossRef]
- McMurry, L.; Petrucci, J.R.E.; Levy, S.B. Active efflux of tetracycline encoded by four genetically different tetracycline resistance determinants in Escherichia coli. Proc. Nat. Acad. Sci. USA 1980, 77, 3974–3977. [Google Scholar] [CrossRef] [Green Version]
- Antunes, N.T.; Assunção, P.; Poveda, J.B.; Tavío, M.M. Mechanisms involved in quinolone resistance in Mycoplasma mycoides subsp. capri. Vet. J. 2015, 204, 327–332. [Google Scholar] [CrossRef]
- Raherison, S.; Gonzalez, P.; Renaudin, H.; Charron, A.; Bébéar, C. Increased Expression of Two Multidrug Transporter-Like Genes Is Associated with Ethidium Bromide and Ciprofloxacin Resistance in Mycoplasma hominis. Antimicrob. Agents Chemother. 2005, 49, 421–424. [Google Scholar] [CrossRef] [Green Version]
- Raherison, S.; Gonzalez, P.; Renaudin, H.; Charron, A.; Bébéar, C. Evidence of Active Efflux in Resistance to Ciprofloxacin and to Ethidium Bromide by Mycoplasma hominis. Antimicrob. Agents Chemother. 2002, 46, 672–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.L.; Sun, H.M.; Zhu, B.L.; Fei, L.I.U.; Zhao, H.Q. Whole Genome Analysis Reveals New Insights into Macrolide Resistance in Mycoplasma pneumoniae. Biomed. Environ. Sci. 2017, 30, 343–350. [Google Scholar]
- Bolhuis, H.; Molenaar, D.; Poelarends, G.; van Veen, H.W.; Poolman, B.; Driessen, A.J.; Konings, W.N. Proton motive force-driven and ATP-dependent drug extrusion systems in multidrug-resistant Lactococcus lactis. J. Bacteriol. 1994, 176, 6957–6964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.-Z.; Nikaido, H. Efflux-Mediated Drug Resistance in Bacteria. Drugs 2009, 69, 1555–1623. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.; McCusker, M.P.; Viveiros, M.; Couto, I.; Fanning, S.; Pagès, J.-M.; Amaral, L. A Simple method for assessment of MDR bacteria for over-expressed efflux pumps. Open Microbiol. J. 2013, 7, 72–82. [Google Scholar] [CrossRef]
- Martins, M.; Viveiros, M.; Couto, I.; Costa, S.S.; Pacheco, T.; Fanning, S.; Pagès, J.-M.; Amaral, L. Identification of efflux pump-mediated multidrug-resistant bacteria by the ethidium bromide-agar cartwheel method. Vivo 2011, 25, 171–178. [Google Scholar]
- Amaral, L.; Cerca, P.; Spengler, G.; Machado, L.; Martins, A.; Couto, I.; Viveiros, M.; Fanning, S.; Pages, J.-M. Ethidium bromide efflux by Salmonella: Modulation by metabolic energy, pH, ions and phenothiazines. Int. J. Antimicrob. Agents 2011, 38, 140–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anes, J.; Hurley, D.; Martins, M.; Fanning, S. Exploring the Genome and Phenotype of Multi-Drug Resistant Klebsiella pneumoniae of Clinical Origin. Front. Microbiol. 2017, 8, 1913. [Google Scholar] [CrossRef] [PubMed]
- Kovacevic, J.; Ziegler, J.; Wałecka-Zacharska, E.; Reimer, A.; Kitts, D.D.; Gilmour, M. Tolerance of Listeria monocytogenes to Quaternary Ammonium Sanitizers Is Mediated by a Novel Efflux Pump Encoded by emrE. Appl. Environ. Microbiol. 2016, 82, 939–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, P.; Hernando-Amado, S.; Reales-Calderon, J.A.; Corona, F.; Lira, F.; Alcalde-Rico, M.; Bernardini, A.; Sanchez, M.B.; Martinez, J.L. Bacterial Multidrug Efflux Pumps: Much More Than Antibiotic Resistance Determinants. Microorganisms 2016, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Khalil, D.; Becker, C.A.; Tardy, F. Alterations in the quinolone resistance-determining regions and fluoroquinolone resistance in clinical isolates and laboratory-derived mutants of Mycoplasma bovis: Not all genotypes may be equal. Appl. Environ. Microbiol. 2016, 82, 1060–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piddock, L.J.V. Clinically relevant chromosomally encoded multidrug resistance efflux pumps in bacteria. Clin. Microbiol. Rev. 2006, 19, 382–402. [Google Scholar] [CrossRef] [Green Version]
- Frempong-Manso, E.; Raygada, J.L.; Demarco, C.E.; Seo, S.M.; Kaatz, G.W. Inability of a reserpine-based screen to identify strains overexpressing efflux pump genes in clinical isolates of Staphylococcus aureus. Int. J. Antimicrob. Agents 2009, 33, 360–363. [Google Scholar] [CrossRef]
- Shirvan, M.H.; Schuldiner, S.; Rottem, S. Role of Na+ cycle in cell volume regulation of Mycoplasma gallisepticum. J. Bacteriol. 1989, 171, 4410–4416. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Isolation Context | MIC (µg/mL) | Mutations 2 | EtBr Efflux | Oxytetracycline Efflux 5 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| No. | Origin 1 | Sample | Year | Host | rrs | rpsJ | Agar 3 | Kinetics 4 | ||
| M. agalactiae | ||||||||||
| L4258 | FR-79 | Mastitic milk | <1989 | Goat | 0.25 | No | nd | n.i. | 44 ± 6 | nd |
| Ag314 | ES-ZA | Bulk tank milk | 2015 | Sheep | 0.5 | No | nd | n.i. | 44 ± 9 | nd |
| F11129 | FR-26 | Bulk tank milk | 2016 | Goat | 0.5 | nd | nd | n.i. | 47 ± 2 | nd |
| L15678 | FR-64 | Bulk tank milk | 2009 | Sheep | 0.5 | No | nd | n.i. | 39 ± 3 | nd |
| L4212c | FR-84 | Mastitic milk | 1986 | Sheep | 0.5 | nd | nd | n.i. | 14 ± 2 | nd |
| 5632 | ES-nk | Joint | <1991 | Goat | 2 | nd | nd | +++ | 37 ± 4 | + (0.8) |
| Ag304 | ES-MA | Mastitic milk | 2015 | Goat | 2 | No | nd | n.i. | 45 ± 7 | ++ (3.5) |
| Ag9 | ES-MU | Mastitic milk | 2009 | Goat | 2 | No | nd | +++ | 37 ± 1 | nd |
| Ag26 | ES-MU | Bulk tank milk | 2000 | Goat | 2 | No | nd | + | 18 ± 3 | nd |
| Ag10 | ES-MU | Mastitic milk | 2008 | Goat | 2 | No | nd | + | 27 ± 11 | nd |
| F10671 | FR-16 | Mastitic milk | 2011 | Goat | 2 | No | nd | + | 39 ± 4 | nd |
| L16086 | FR-84 | Mastitic milk | 2013 | Goat | 2 | No | nd | +++ | 38 ± 4 | nd |
| L15242 | FR-86 | Mastitic milk | 2009 | Goat | 2 | No | nd | +++ | 40 ± 9 | nd |
| Ag28 | ES-MU | Bulk tank milk | 2000 | Goat | 4 | Yes | nd | n.i. | 47 ± 10 | ++ (1.9) |
| L16160 | FR-49 | Bulk tank milk | 2010 | Goat | 4 | No | No | +++ | 38 ± 4 | ++ (1.8) |
| L16156 | FR-79 | Bulk tank milk | 2010 | Goat | 4 | No | No | + | 40 ± 3 | nd |
| Ag316 | ES-AL | Mastitic milk | 2015 | Goat | 8 | Yes | No | n.i. | 43 ± 7 | ++ (1.7) |
| M. capricolum subsp. capricolum | ||||||||||
| Cap3 | ES-AN | Mastitic milk | 2011 | Goat | 0.25 | No | nd | + | 39 ± 1 | nd |
| Cap7 | ES-MA | Auricular swab | 2009 | Goat | 0.25 | nd | No | +++ | 34 ± 9 | nd |
| Cap19 | ES-GC | Mastitic milk | 2015 | Goat | 0.25 | nd | nd | +++ | 52 ± 2 | − (0.0) |
| F10621 | FR-79 | Mastitic milk | 2015 | Goat | 2 | No | Yes | +/− | 46 ± 2 | − (0.1) |
| M. mycoides subsp. capri | ||||||||||
| LC84 | ES-GC | Mastitic milk | 2014 | Goat | 0.5 | No | nd | +++ | 41 ± 8 | nd |
| LC33 | ES-MU | Bulk tank milk | 2009 | Goat | 0.5 | No | nd | ++ | 39 ± 4 | nd |
| LC54 | ES-GC | Bulk tank milk | 2004 | Goat | 2 | No | Yes | +++ | 35 ± 10 | ++ (1.5) |
| F9545 | FR-81 | Lung | 2012 | Goat | 4 | No | Yes | +++ | 40 ± 5 | + (0.9) |
| F10751 | FR-79 | Mastitic milk | 2014 | Goat | 4 | No | No | +++ | 36 ± 1 | ++ (2.2) |
| M. putrefaciens | ||||||||||
| F10828 | FR-74 | Mastitic milk | 2016 | Goat | 0.5 | nd | nd | n.i. | 34 ± 2 | nd |
| Put13 | ES-GC | Mastitic milk | 2015 | Goat | 0.5 | No | nd | n.i. | 34 ± 2 | nd |
| F5435 | FR-36 | Mastitic milk | 2008 | Goat | 0.5 | No | nd | Inh. | 32 ± 3 | nd |
| F8131 | FR-23 | Mastitic milk | 2013 | Goat | 0.5 | No | nd | Inh. | 37 ± 5 | nd |
| F1174 | FR-79 | Bulk tank milk | 2004 | Goat | 0.5 | nd | nd | Inh. | 37 ± 3 | nd |
| Put9 | ES-GC | Mastitic milk | 2014 | Goat | 0.5 | nd | nd | Inh. | 33 ± 4 | nd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tatay-Dualde, J.; Prats-van der Ham, M.; Gaurivaud, P.; de la Fe, C.; Tardy, F. Efflux Might Participate in Decreased Susceptibility to Oxytetracycline in Contagious Agalactia-Causative Mycoplasma spp. Animals 2021, 11, 2449. https://doi.org/10.3390/ani11082449
Tatay-Dualde J, Prats-van der Ham M, Gaurivaud P, de la Fe C, Tardy F. Efflux Might Participate in Decreased Susceptibility to Oxytetracycline in Contagious Agalactia-Causative Mycoplasma spp. Animals. 2021; 11(8):2449. https://doi.org/10.3390/ani11082449
Chicago/Turabian StyleTatay-Dualde, Juan, Miranda Prats-van der Ham, Patrice Gaurivaud, Christian de la Fe, and Florence Tardy. 2021. "Efflux Might Participate in Decreased Susceptibility to Oxytetracycline in Contagious Agalactia-Causative Mycoplasma spp." Animals 11, no. 8: 2449. https://doi.org/10.3390/ani11082449