
Effects of Bait on Male White-Tailed Deer Resource Selection

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
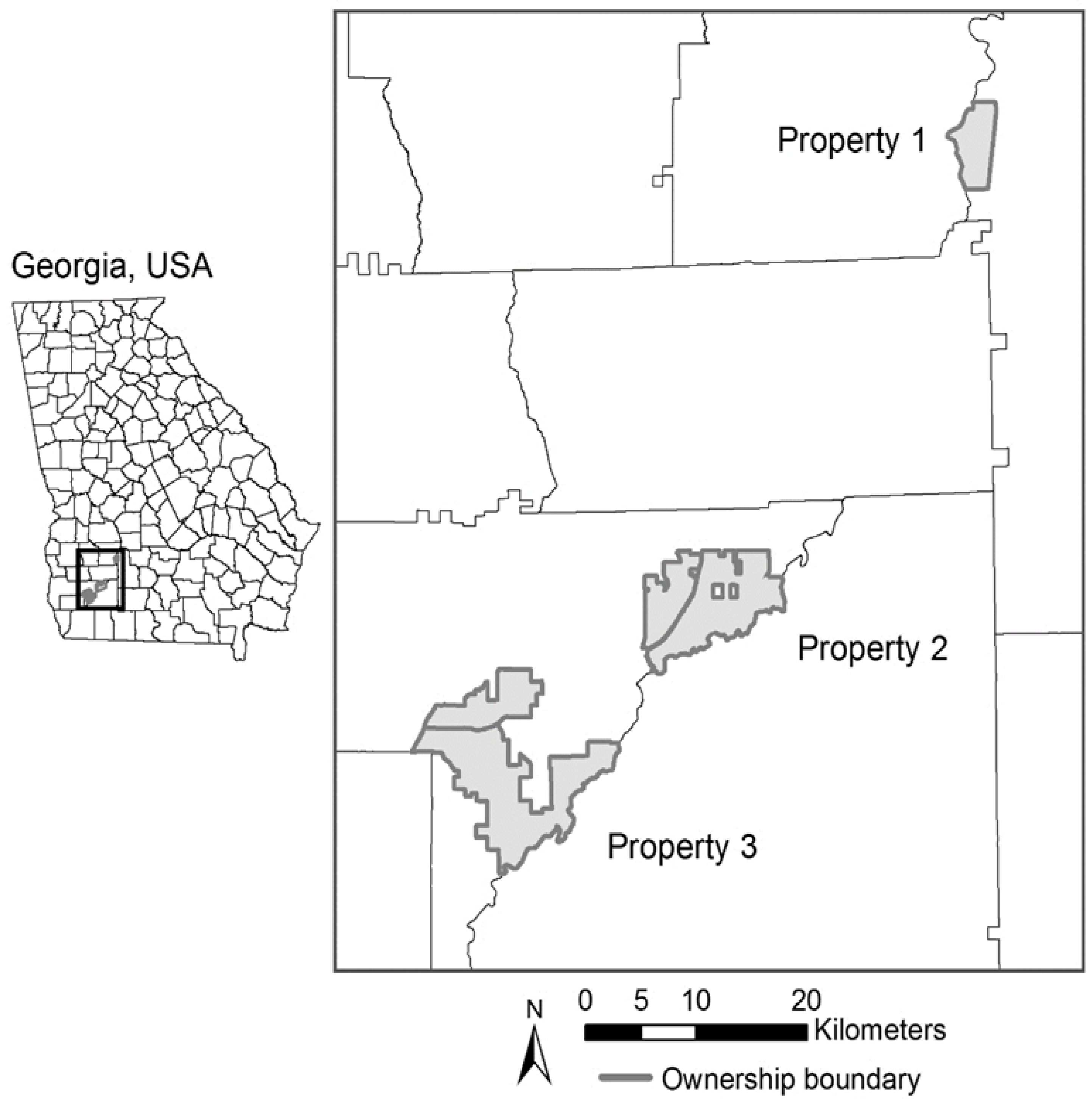
2.1. Study Area
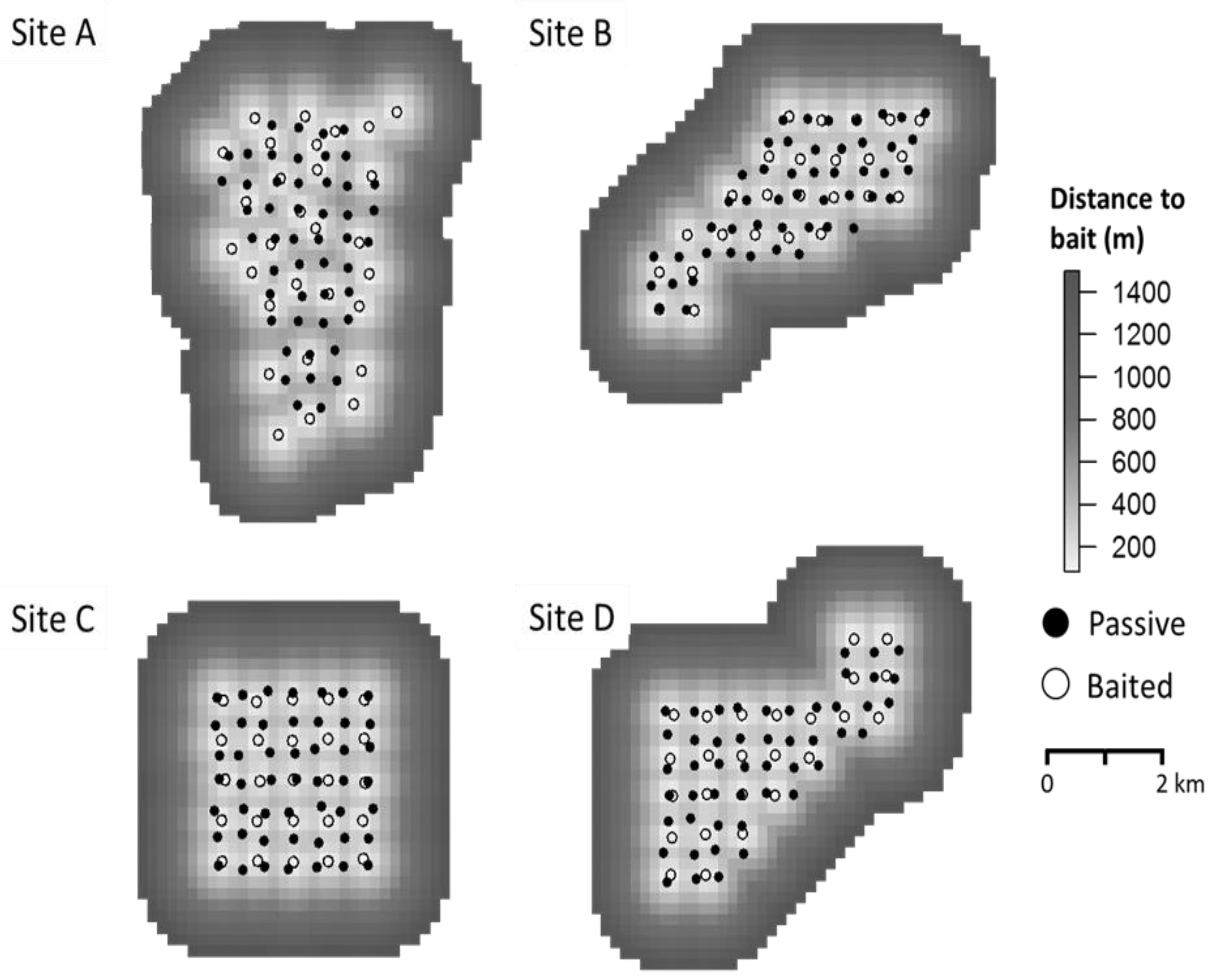
2.2. Spatial Model
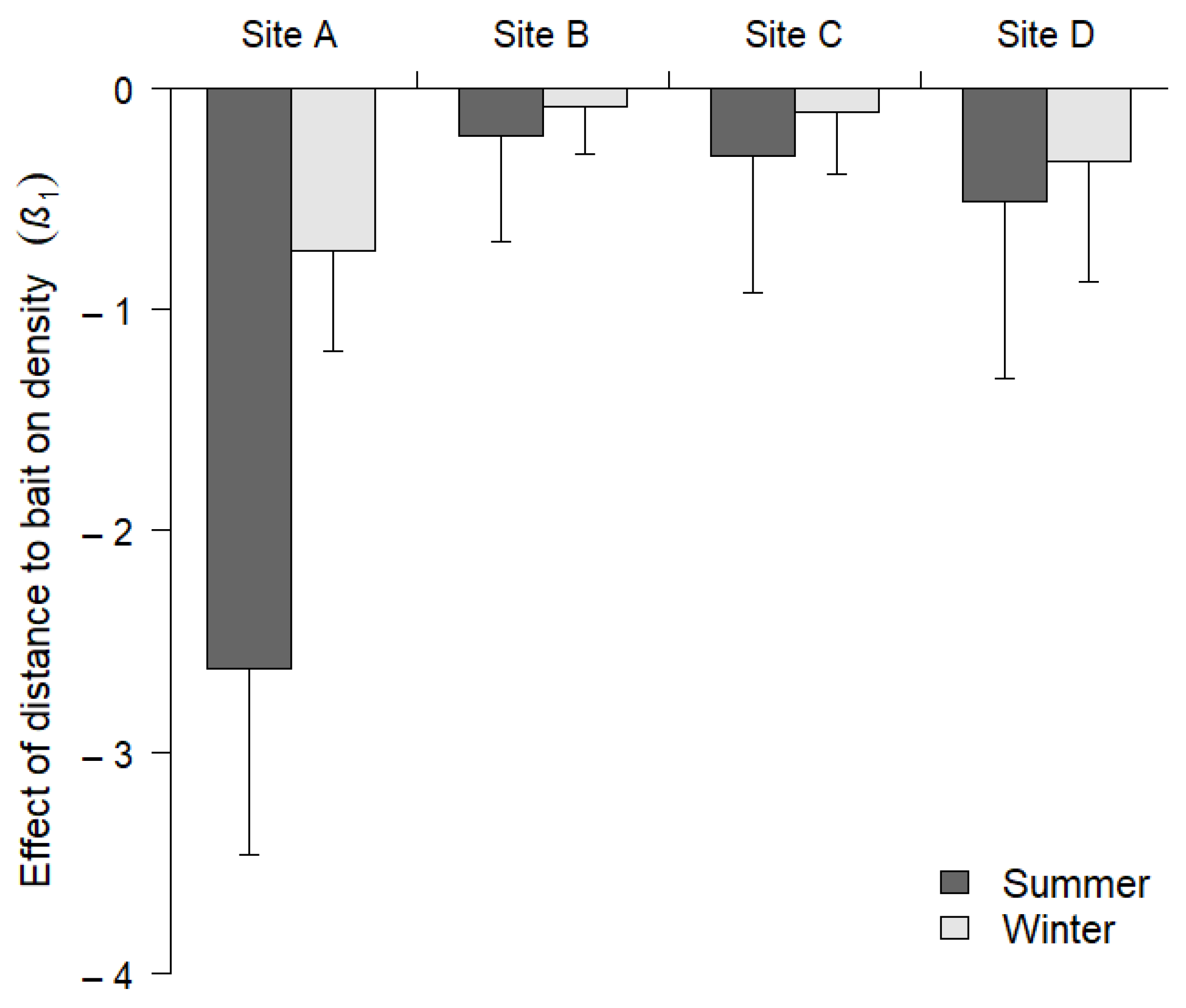
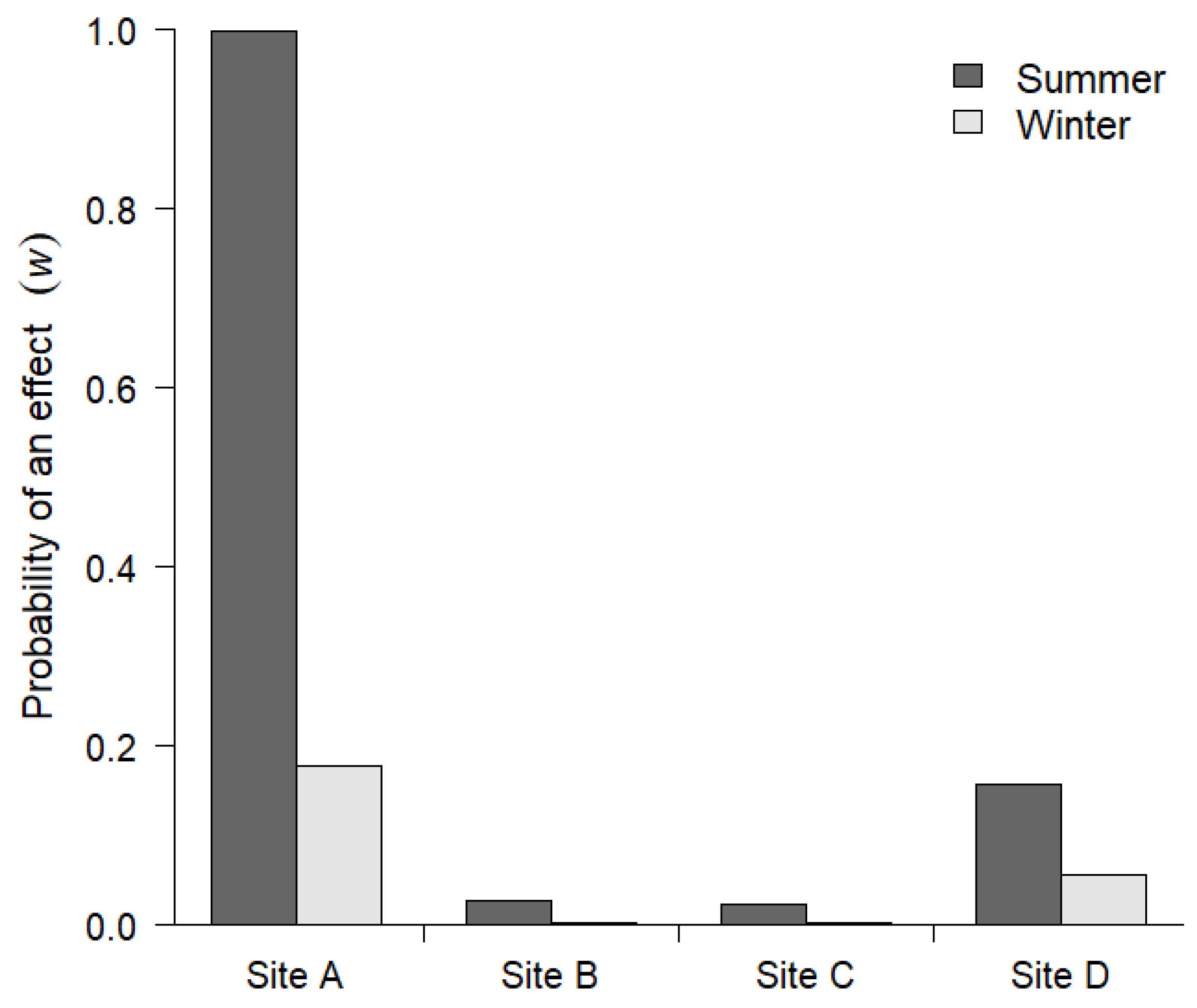
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. SCR JAGS Model
| model { |
| p0[1] ~ dbeta(1,1) |
| p0[2] ~ dbeta(1,1) |
| sigma ~ dgamma(1,1) |
| beta0 ~ dnorm(0, 0.1) |
| beta1 ~ dunif(−100,0) |
| w ~ dbern(0.5) |
| for(g in 1:nPixels) { |
| mu[g] <- exp(beta0 + beta1*bait.dist[g]*w)*pixelArea |
| pi[g] <- mu[g]/sum(mu)} |
| EN <- sum(mu) |
| psi <- EN animals-11-02334M |
| for(i in 1:M) { |
| z[i] ~ dbern(psi) |
| s[i] ~ dcat(pi[]) |
| for(j in 1:J) { |
| p[i,j] <- p0[bait[j]]*exp(-1*pow(d[s[i],j],2)/(2*sigma^2)) |
| for(k in 1:K) { |
| y[i,j,k] ~ dbern(p[i,j]*z[i]*oper[j,k])}}} |
| N <- sum(z) |
| } |
References
- Langenau, E.E., Jr.; Flegerler, E., Jr.; Hill, H. Deer Hunters’ Opinion Survey; Michigan Department of Natural Resources: Lansing, MI, USA, 1985.
- Wisconsin Bureau of Wildlife Management. Deer Baiting in Wisconsin: A Survey of Wisconsin Deer Hunters; Wisconsin Department of Natural Resources, Bureau of Wildlife Management: Madison, WI, USA, 1993.
- Frawley, B.J. 1999 Michigan Deer Hunter Survey: Deer Baiting; Wildlife Division Report No. 3315; Michigan Department of Natural Resources: Lansing, MI, USA, 2000.
- Kilpatrick, H.J.; Labonte, A.M.; Barclay, J.S. Use of bait to increase archery deer harvest in an urban-suburban landscape. J. Wildl. Manag. 2010, 74, 714–718. [Google Scholar] [CrossRef]
- Jacobson, H.A.; Kroll, J.C.; Browning, R.W.; Koerth, B.H.; Conway, M.H. Infrared-triggered cameras for censusing white-tailed deer. Wildl. Soc. Bull. 1997, 25, 547–556. [Google Scholar]
- Mills, D.; Fattebert, J.; Hunter, L.; Slotow, R. Maximising camera trap data: Using attractants to improve detection of elusive species in multi-species surveys. PLoS ONE 2019, 14, e0216447. [Google Scholar] [CrossRef]
- Powell, R.A.; Proulx, G. Trapping and marking terrestrial mammals for research: Integrating ethics, performance criteria, techniques, and common sense. ILAR J. 2003, 44, 259–276. [Google Scholar] [CrossRef] [Green Version]
- Harkins, K.M.; Keinath, D.; Ben-David, M. It’s a trap: Optimizing detection of rare small mammals. PLoS ONE 2019, 14, e0213201. [Google Scholar] [CrossRef]
- Waller, D.M.; Alverson, W.S. The white-tailed deer: A keystone herbivore. Wildl. Soc. Bull. 1997, 25, 217–226. [Google Scholar]
- Pimm, S.L.; Rosenzweig, M.L.; Mitchell, W. Competition and food selection: Field tests of a theory. Ecology 1985, 66, 798–807. [Google Scholar] [CrossRef]
- Raman, S. Ecology and management of chital and blackbuck in Guindy National Park, Madras. J. Bombay Nat. Hist. Soc. 1996, 93, 178–192. [Google Scholar]
- Eve, J.H. Management implications of disease. In Diseases and Parasites of White-Tailed Deer; Davidson, W.R., Ed.; Tall Timbers Research Station: Tallahassee, FL, USA, 1981; pp. 413–423. [Google Scholar]
- Schmitt, S.M.; Fitzgerald, S.D.; Cooley, T.M.; Bruning-Fann, C.S.; Sullivan, L.; Berry, D.; Carlson, T.; Minnus, R.B.; Payeur, J.B.; Sikarskie, J. Bovine tuberculosis in free-ranging white-tailed deer from Michigan. J. Wildl. Dis. 1997, 33, 749–758. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.D.; Cooper, S.M. The nutritional, ecological, and ethical arguments against baiting and feeding white-tailed deer. Wildl. Soc. Bull. 2006, 34, 519–524. [Google Scholar] [CrossRef]
- Sorensen, A.; van Beest, F.M.; Brook, R.K. Impacts of wildlife baiting and supplemental feeding on infectious disease transmission risk: A synthesis of knowledge. Prev. Vet. Med. 2014, 113, 356–363. [Google Scholar] [CrossRef]
- Synatzke, D.R. Effects of baiting on white-tailed deer hunting success. In Federal Aid in Wildlife Restoration Report Project W109-R-4; Texas Parks and Wildlife Department: Austin, TE, USA, 1981. [Google Scholar]
- Jacobson, H.A.; Darrow, D.A. Effects of baiting on deer movements and activity. In Proceedings of the Annual Southeast Deer Study Group Meeting, Annapolis, MD, USA, 23–26 February 1992; Volume 15, p. 23. [Google Scholar]
- Garner, M.S. Movement Patterns and Behavior at Winter Feeding and Fall Baiting Stations in a Population of White-Tailed Deer Infected with Bovine Tuberculosis in the Northeastern Lower Peninsula of Michigan. Ph.D. Thesis, Michigan State University, East Lansing, MI, USA, 2001. [Google Scholar]
- Kilpatrick, H.J.; Stober, W.A. Effects of temporary bait sites on movements of suburban white-tailed deer. Wildl. Soc. Bull. 2002, 30, 760–766. [Google Scholar]
- Palmer, M.V.; Whipple, D.L.; Waters, W.R. Experimental deer-to-deer transmission of Mycobacterium bovis. Am. J. Vet. Res. 2001, 62, 692–696. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, S.M.; Rudolph, B.A.; Winterstein, S.R.; Campa, H.; Garner, M.; Muzo, D. Eradicating bovine TB in white-tailed deer in Michigan: Identifying avenues of within herd transmission and deer migratory and movement behaviors. In Wildlife Research Summaries 2002; Wildlife Division Report No. 3368; Hill, H.R., Ed.; Michigan Department of Natural Resources: Lansing, MI, USA, 2002; pp. 27–28. [Google Scholar]
- Adams, K.; Ross, M. QDMA’s Whitetail Report; Quality Deer Management Association: Bogart, GA, USA, 2017. [Google Scholar]
- Gillin, C.M.; Mawdsley, J.R. AFWA Technical Report on Best Management Practices for Surveillance, Management and Control of Chronic Wasting Disease; Association of Fish and Wildlife Agencies: Washington, DC, USA, 2018. [Google Scholar]
- Darrow, D.A. Effects of Baiting on Deer Movement and Activity. Master’s Thesis, Mississippi State University, Starksville, MI, USA, 1993. [Google Scholar]
- Williams, S.C.; DeNicola, A.J. Spatial Movements in Response to Baiting Female White-Tailed Deer. In Proceedings of the 9th Wildlife Damage Management Conference, State College, PA, USA, 5–8 October 2000; pp. 206–224. [Google Scholar]
- Beaver, J.T. White-Tailed Deer Distributions and Movement Behavior in South-Central Texas, USA. Ph.D. Thesis, Texas A&M University, College Station, TX, USA, 2017. [Google Scholar]
- Murden, S.B.; Risenhoover, K.L. Effects of habitat enrichment on patterns of diet selection. Ecol. Appl. 1993, 3, 497–505. [Google Scholar] [CrossRef]
- Johnson, D.H. The comparison of usage and availability measurements for evaluating resource preference. Ecology 1980, 61, 65–71. [Google Scholar] [CrossRef]
- Nixon, C.M.; Hansen, L.P.; Brewer, P.A.; Chelsvig, J.E. Ecology of white-tailed deer in an intensively farmed region of Illinois. Wildl. Mono 1991, 118, 1–77. [Google Scholar]
- Borchers, D.L.; Efford, M. Spatially explicit maximum likelihood methods for capture-recapture studies. Biomet 2008, 64, 377–385. [Google Scholar] [CrossRef]
- Royle, J.A.; Chandler, R.B.; Sollmann, R.; Gardner, B. Spatial Capture-Recapture; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Royle, J.A.; Chandler, R.B.; Sun, C.C.; Fuller, A.K. Integrating resource selection information with spatial capture-recapture. Meth. Ecol. Evol. 2013, 4, 520–530. [Google Scholar] [CrossRef] [Green Version]
- Royle, J.A.; Young, K.V. A hierarchical model for spatial capture-recapture data. Ecology 2008, 89, 2281–2289. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 7 July 2020).
- Plummer, M. ‘rjags: Bayesian Graphical Models Using MCMC’. In R Package Version 4–6. Available online: https://CRAN.R-project.org/package=rjags (accessed on 3 March 2019).
- Plummer, M. JAGS: A program for analysis of Bayesian graphical models using Gibbs sampling. In Proceedings of the 3rd International Workshop on Distributed Statistical Computing, Vienna, Austria, 20–22 March 2003. [Google Scholar]
- Gelman, A.; Rubin, D.B. Inference from interactive simulation using multiple sequences. Stat. Sci. 1992, 7, 457–472. [Google Scholar] [CrossRef]
- VerCauteren, K.C.; Hyngstrom, S.E. Effects of agricultural activities and hunting on home ranges of female white-tailed deer. J. Wildl. Manag. 1998, 62, 280–285. [Google Scholar] [CrossRef]
- Walter, W.D.; VerCauteren, K.C.; Gilsdorf, J.M.; Hygnstrom, S.E. Crop, native vegetation, and bio-fuels: Response of white-tailed deer to changing management priorities. J. Wildl. Manag. 2009, 73, 339–344. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.W. Temporal and spatial dynamics of chronic wasting disease epidemics. In Chronic Wasting Disease Symposium; Kahn, R.H., Ed.; Division of Wildlife: Fort Collins, CO, USA, 2002. [Google Scholar]
- Williams, E.S.; Miller, M.W.; Kreeger, T.J.; Kahn, R.H.; Thorne, E.T. Chronic Wasting disease of deer and elk: A review with recommendations. J. Wildl. Manag. 2002, 66, 551–563. [Google Scholar] [CrossRef]
- Hawkins, R.E.; Klimstra, W.D. A preliminary study of the social organization of white-tailed deer. J. Wildl. Manag. 1970, 34, 407–419. [Google Scholar] [CrossRef]
- Marchinton, R.L.; Hirth, D.H. Behavior. In White-Tailed Deer: Ecology and Management; Halls, L.K., Ed.; Stackpole Books: Harrisburg, PA, USA, 1984; pp. 129–168. [Google Scholar]
- Brinkman, T.J.; DePerno, C.S.; Jenks, J.A.; Haroldson, B.S. Movement of female white-tailed deer: Effects of climate and intense row crop agriculture. J. Wildl. Manag. 2005, 69, 1099–1111. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site A | Site B | Site C | Site D | |
|---|---|---|---|---|
| Summer baited photos | 10,613 | 3609 | 3006 | 2188 |
| Summer passive photos | 240 | 143 | 57 | 48 |
| Summer unique male individuals | 208 | 93 | 94 | 75 |
| Winter baited photos | 11,318 | 3587 | 2222 | 2502 |
| Winter passive photos | 141 | 114 | 67 | 68 |
| Winter unique male individuals | 177 | 85 | 89 | 72 |
| Summer | Winter | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Site | Parameter | Mean | SD | 2.5% | 97.5% | Mean | SD | 2.5% | 97.5% |
| A | N | 261.5 | 8.925 | 247.0 | 282.0 | 214.0 | 5.167 | 205.0 | 225.0 |
| β0 | 3.247 | 0.140 | 2.972 | 3.521 | 2.133 | 0.134 | 1.869 | 2.392 | |
| β1 | −2.621 | 0.416 | −3.456 | −1.838 | −0.734 | 0.223 | −1.182 | −0.309 | |
| p0 [passive] | 0.028 | 0.002 | 0.023 | 0.033 | 0.012 | 0.001 | 0.010 | 0.015 | |
| p0 [baited] | 0.673 | 0.017 | 0.641 | 0.707 | 0.414 | 0.011 | 0.393 | 0.436 | |
| σ | 0.338 | 0.004 | 0.331 | 0.345 | 0.607 | 0.008 | 0.592 | 0.623 | |
| B | N | 138.6 | 8.041 | 124.0 | 155.0 | 100.3 | 1.180 | 99.0 | 103.0 |
| β0 | 1.529 | 0.125 | 1.312 | 1.805 | 1.112 | 0.099 | 0.922 | 1.315 | |
| β1 | −0.210 | 0.174 | −0.651 | −0.007 | −0.081 | 0.074 | −0.275 | −0.002 | |
| p0 [passive] | 0.028 | 0.004 | 0.022 | 0.036 | 0.011 | 0.001 | 0.009 | 0.014 | |
| p0 [baited] | 0.420 | 0.023 | 0.378 | 0.467 | 0.255 | 0.011 | 0.233 | 0.278 | |
| σ | 0.487 | 0.012 | 0.463 | 0.510 | 0.872 | 0.018 | 0.838 | 0.907 | |
| C | N | 168.5 | 14.598 | 141.0 | 198.0 | 108.2 | 4.387 | 101.0 | 118.0 |
| β0 | 1.940 | 0.124 | 1.719 | 2.207 | 1.367 | 0.108 | 1.160 | 1.589 | |
| β1 | −0.302 | 0.237 | −0.874 | −0.011 | −0.108 | 0.097 | −0.358 | −0.003 | |
| p0 [passive] | 0.020 | 0.003 | 0.014 | 0.027 | 0.014 | 0.002 | 0.010 | 0.018 | |
| p0 [baited] | 0.370 | 0.021 | 0.331 | 0.411 | 0.244 | 0.014 | 0.219 | 0.272 | |
| σ | 0.375 | 0.009 | 0.358 | 0.394 | 0.652 | 0.019 | 0.615 | 0.691 | |
| D | N | 104.6 | 8.606 | 89.0 | 123.0 | 85.5 | 2.349 | 82.0 | 91.0 |
| β0 | 1.459 | 0.183 | 1.137 | 1.848 | 1.141 | 0.174 | 0.832 | 1.511 | |
| β1 | −0.509 | 0.333 | −1.263 | −0.031 | −0.327 | 0.223 | −0.838 | −0.016 | |
| p0 [passive] | 0.022 | 0.004 | 0.015 | 0.030 | 0.013 | 0.002 | 0.009 | 0.018 | |
| p0 [baited] | 0.457 | 0.025 | 0.409 | 0.506 | 0.263 | 0.013 | 0.239 | 0.289 | |
| σ | 0.443 | 0.012 | 0.421 | 0.467 | 0.750 | 0.019 | 0.713 | 0.789 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, J.T.; Chandler, R.B.; Conner, L.M.; Cherry, M.J.; Killmaster, C.H.; Johannsen, K.L.; Miller, K.V. Effects of Bait on Male White-Tailed Deer Resource Selection. Animals 2021, 11, 2334. https://doi.org/10.3390/ani11082334
Johnson JT, Chandler RB, Conner LM, Cherry MJ, Killmaster CH, Johannsen KL, Miller KV. Effects of Bait on Male White-Tailed Deer Resource Selection. Animals. 2021; 11(8):2334. https://doi.org/10.3390/ani11082334
Chicago/Turabian StyleJohnson, James T., Richard B. Chandler, L. Mike Conner, Michael J. Cherry, Charlie H. Killmaster, Kristina L. Johannsen, and Karl V. Miller. 2021. "Effects of Bait on Male White-Tailed Deer Resource Selection" Animals 11, no. 8: 2334. https://doi.org/10.3390/ani11082334