
Sexual Dimorphism in Crowned Lemur Scent-Marking


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and Housing
2.2. Behavioural Data Collection and Analysis
2.3. Odour Sampling and Analysis
3. Results
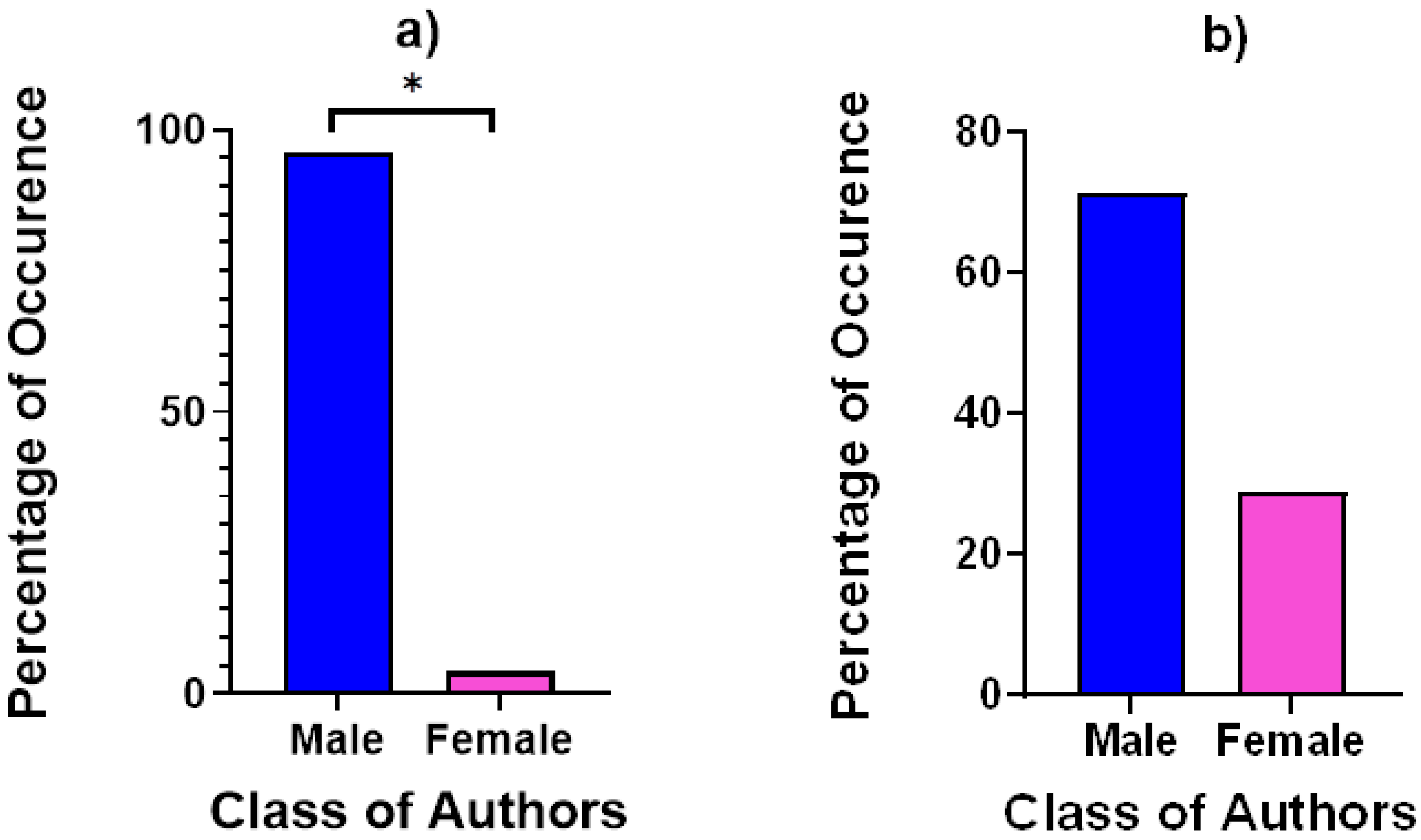
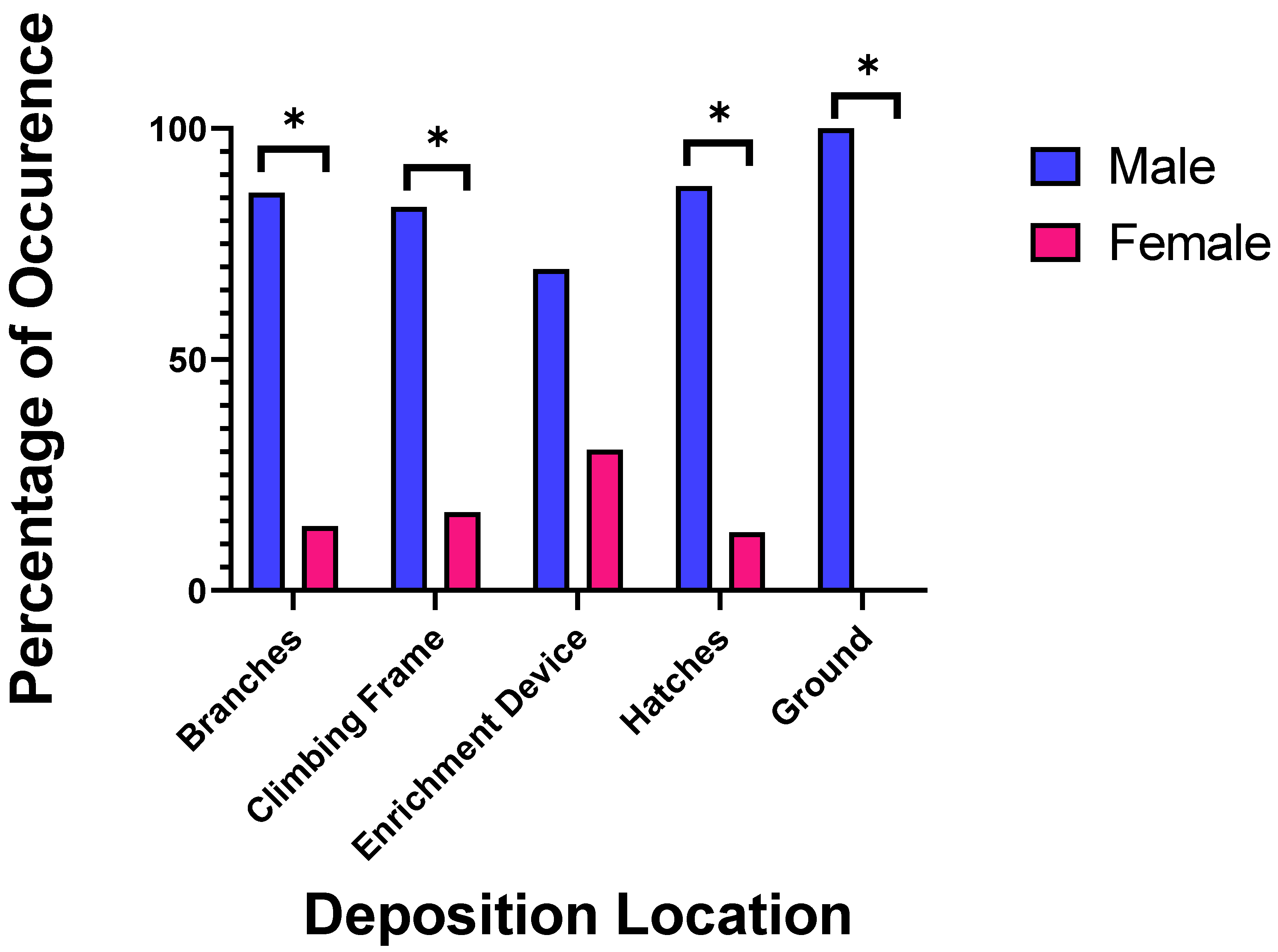
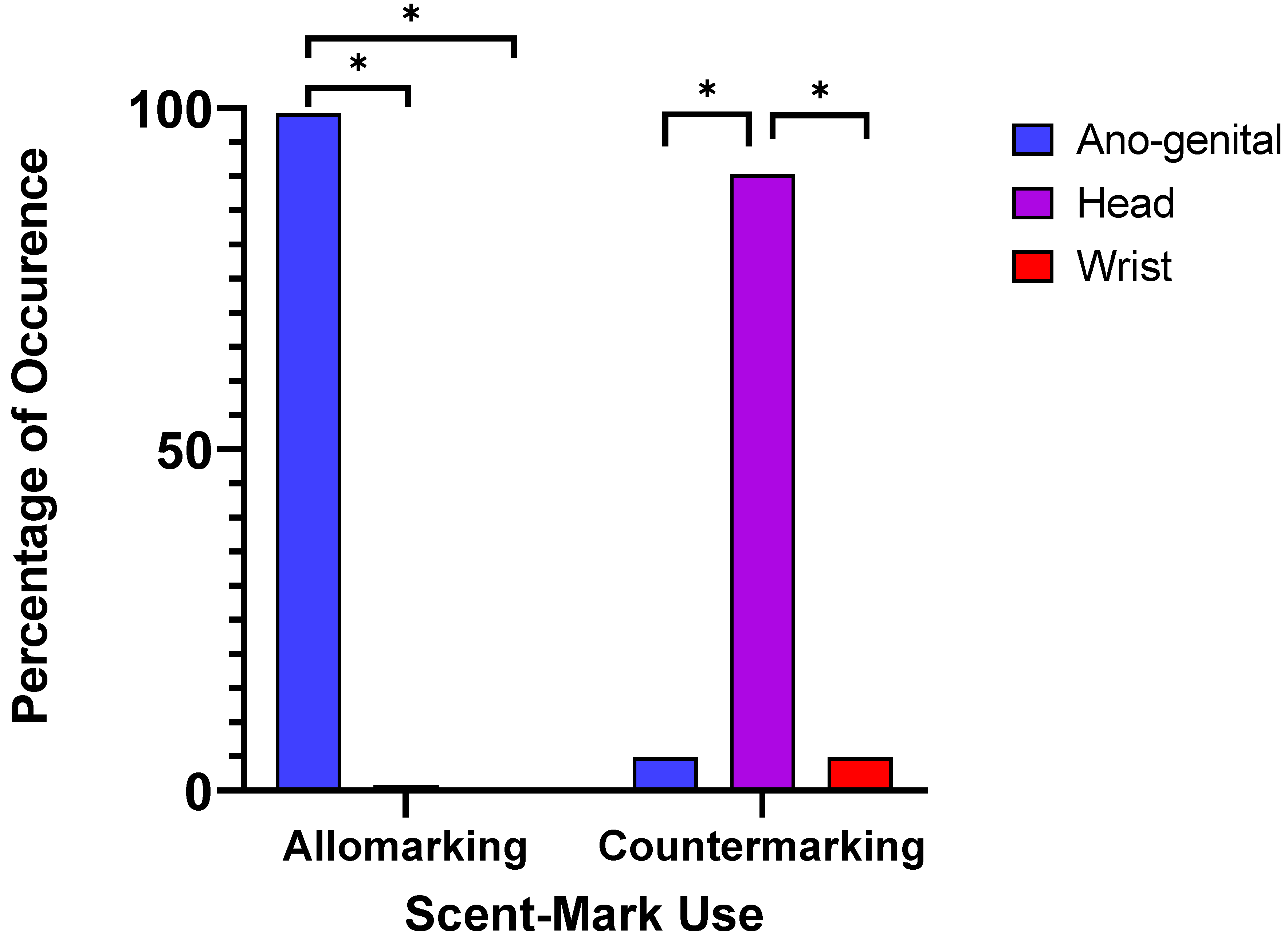
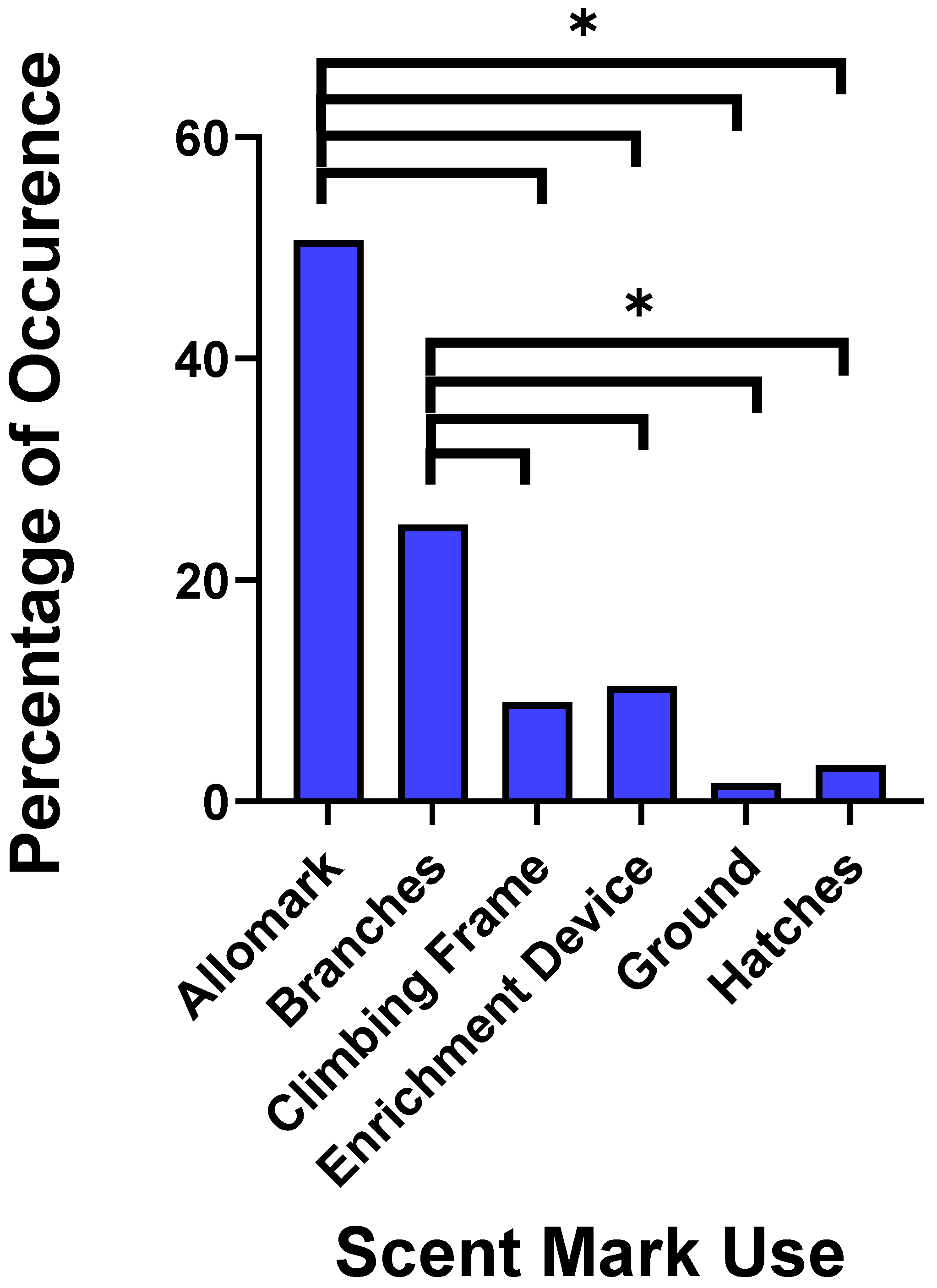
3.1. Behavioural Observations
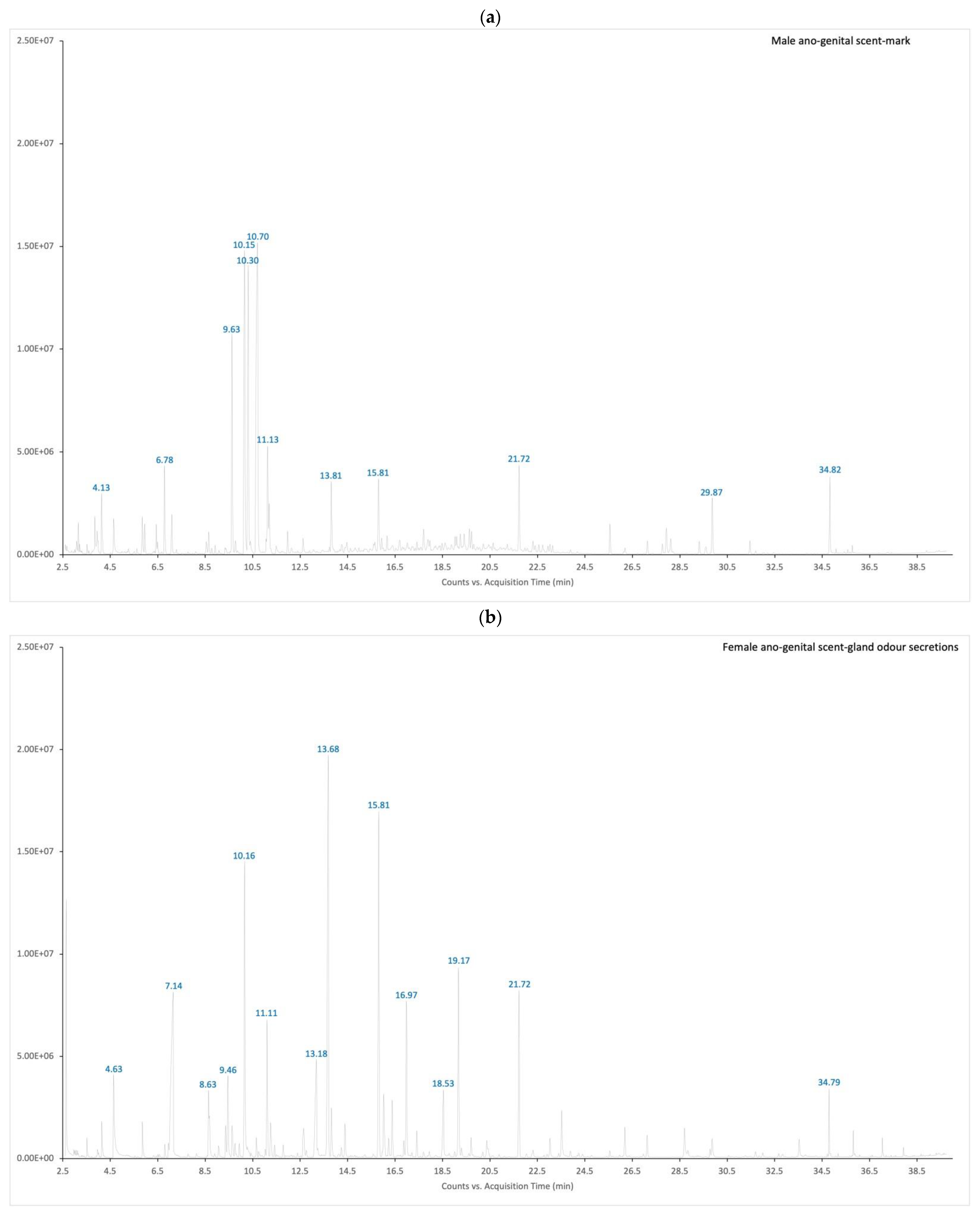
3.2. Odour Secretions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seyfarth, R.M.; Cheney, D.L. Signalers and receivers in animal communication. Annu. Rev. Psychol. 2003, 54, 145–173. [Google Scholar] [CrossRef] [Green Version]
- Osorio, D.; Vorobyev, M. A review of the evolution of animal colour vision and visual communication signals. Vision Res. 2008, 48, 2042–2051. [Google Scholar] [CrossRef] [Green Version]
- Fitch, W.T.; Hauser, M.D. Unpacking “honesty”: Vertebrate vocal production and the evolution of acoustic signals. In Acoustic Communication; Simmons, A.M., Fay, R.R., Popper, A.N., Eds.; Springer: New York, NY, USA, 2003; Volume 16. [Google Scholar]
- Campbell-Palmer, R.; Rosell, F. The importance of chemical communication studies to mammalian conservation biology: A review. Biol. Conserv. 2011, 144, 1919–1930. [Google Scholar] [CrossRef] [Green Version]
- Wackermannová, M.; Pinc, L.; Jebavý, L. Olfactory sensitivity in mammalian species. Physiol. Res. 2016, 65, 369–390. [Google Scholar] [CrossRef] [PubMed]
- Scordato, E.S.; Drea, C.M. Scents and sensibility: Information content of olfactory signals in the ring-tailed lemur, Lemur catta. Anim. Behav. 2007, 73, 301–314. [Google Scholar] [CrossRef]
- Hayes, R.A.; Morelli, T.L.; Wright, P.C. Volatile components of lemur scent secretions vary throughout the year. Am. J. Primatol. 2006, 68, 1202–1207. [Google Scholar] [CrossRef] [PubMed]
- Nevo, O.; Heymann, E.W. Led by the nose: Olfaction in primate feeding ecology. Evol. Anthropol. 2015, 24, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Hurst, J.L.; Beynon, R.J. Scent wars: The chemobiology of competitive signaling in mice. BioEssays 2004, 26, 1288–1298. [Google Scholar] [CrossRef]
- Kücklich, M.; Weiß, B.M.; Birkemeyer, C.; Einspanier, A.; Widdig, A. Chemical cues of female fertility states in a non-human primate. Sci. Rep. 2019, 9, 13716. [Google Scholar] [CrossRef] [Green Version]
- Vaglio, S.; Minicozzi, P.; Romoli, R.; Boscaro, F.; Pieraccini, G.; Moneti, G.; Moggi-Cecchi, J. Sternal gland scent-marking signals sex, age, rank and group identity in captive mandrills. Chem. Senses 2016, 41, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Janda, E.D.; Perry, K.L.; Hankinson, E.; Walker, D.; Vaglio, S. Sex differences in scent-marking in captive red-ruffed lemurs. Am. J. Primatol. 2019, 81, e22951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DelBarco-Trillo, J.; Sacha, C.R.; Dubay, G.R.; Drea, C.M. Eulemur, me lemur: The evolution of scent-signal complexity in a primate clade. Philos. Trans. R. Soc. B 2012, 367, 1909–1922. [Google Scholar] [CrossRef] [Green Version]
- Greene, L.K.; Grogan, K.E.; Smyth, K.N.; Adams, C.A.; Klager, S.A.; Drea, C.M. Mix it and fix it: Functions of composite olfactory signals in ring-tailed lemurs. R. Soc. Open Sci. 2016, 3, 160076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kücklich, M.; Möller, M.; Marcillo, A.; Einspanier, A.; Weiß, B.M.; Birkemeyer, C.; Widdig, A. Different methods for volatile sampling in mammals. PLoS ONE 2017, 12, e0183440. [Google Scholar]
- Roberts, S.C. Scent-marking. In Rodent Societies: An ecological and Evolutionary Perspective; Wolff, J.O., Sherman, P.W., Eds.; Chicago University Press: Chicago, IL, USA, 2007. [Google Scholar]
- Gosling, L.M.; Roberts, S.C. Scent-marking by male mammals: Cheat-proof signals to competitors and mates. Adv. Study Behav. 2001, 30, 169–217. [Google Scholar]
- Colquhoun, I.C. A review and interspecific comparison of nocturnal and cathemeral strepsirhine primate olfactory behavioural ecology. Int. J. Zool. 2011, 362976. [Google Scholar] [CrossRef] [Green Version]
- Drea, C.M. D’scent of man: A comparative survey of primate chemosignaling in relation to sex. Horm. Behav. 2015, 68, 117–133. [Google Scholar] [CrossRef] [PubMed]
- Gildersleeve, K.A.; Haselton, M.G.; Larson, C.M.; Pillsworth, E.G. Body odor attractiveness as a cue of impending ovulation in women: Evidence from a study using hormone-confirmed ovulation. Horm. Behav. 2012, 61, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Henkel, S.; Setchell, J.M. Group and kin recognition via olfactory cues in chimpanzees (Pan troglodytes). Proc. R. Soc. Lond. Biol. 2018, 285, 20181527. [Google Scholar] [CrossRef] [Green Version]
- Jänig, S.; Weiß, B.M.; Birkemeyer, C.; Widdig, A. Comparative chemical analysis of body odor in great apes. Am. J. Primatol. 2019, 81, e22976. [Google Scholar] [CrossRef]
- Singletary, B.; Tecot, S. Signaling across the senses: A captive case study in pair-bonded red-bellied lemurs (Eulemur rubriventer) at the Duke Lemur Center, NC, USA. Primates 2019, 60, 499–505. [Google Scholar] [CrossRef]
- Kappeler, P.M. A preliminary study of olfactory behavior of captive Lemur coronatus during the breeding season. Int. J. Primatol. 1988, 9, 135–146. [Google Scholar] [CrossRef]
- Gould, L.; Overdoff, D.J. Adult male scent-marking in Lemur catta and Eulemur fulvus rufus. Int. J. Primatol. 2002, 23, 575–586. [Google Scholar] [CrossRef]
- Scordato, E.; Dubay, G.; Drea, C. Chemical composition of scent marks in the ring-tailed lemur (Lemur catta): Glandular differences, seasonal variation, and individual signatures. Chem. Senses 2007, 32, 493–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuter, K.E.; Eppley, T.M.; Hending, D.; Pacifici, M.; Semel, B.; Zaonarivelo, J. Eulemur coronatus. The IUCN Red List of Threatened Species: E.T8199A115561046. 2020. Available online: https://www.iucnredlist.org/fr/species/8199/182239524 (accessed on 21 May 2021).
- Kappeler, P.M. Agonistic and grooming behavior of captive crowned lemurs (Lemur coronatus) during the breeding season. J. Hum. Evol. 1989, 4, 207–215. [Google Scholar] [CrossRef]
- Marolf, B.; McElligott, A.G.; Müller, A. Female social dominance in two Eulemur species with different social organizations. Zoo Biol. 2007, 26, 201–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamba, M.; Giacoma, C. Quantitative acoustic analysis of the vocal repertoire of the crowned lemur. Ethol. Ecol. Evol. 2007, 19, 323–343. [Google Scholar] [CrossRef]
- Kappeler, P.M. Reproduction in the crowned lemur (Lemur coronatus) in captivity. Am. J. Primato. 1987, 12, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Altmann, J. Observational study of behaviour: Sampling methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef] [Green Version]
- George, D.; Mallery, P. IBM SPSS Statistics 26. A Simple Guide and Reference; Routledge: New York, NY, USA; London, UK, 2020. [Google Scholar]
- Walker, D.; Vaglio, S. Sampling and analysis of animal scent signals. J. Vis. Exp. 2021, 168, e60902. [Google Scholar]
- Dulac, C.; Torello, A.T. Molecular detection of pheromone signals in mammals: From genes to behaviour. Nat. Rev. Neurosci. 2003, 4, 551–562. [Google Scholar] [CrossRef] [PubMed]
- Gilad, Y.; Wiebe, V.; Prezeworski, M.; Lancet, D.; Pääbo, S. Loss of olfactory receptor genes coincides with the acquisition of full trichromatic vision in primates. PLoS Biol. 2004, 2, 120–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negus, V. The Comparative Anatomy and Physiology of the Nose and Paranasal Sinuses; Livingston: Edinburgh, UK, 1958. [Google Scholar]
- Dominy, N.J.; Lucas, P.W. Ecological importance of trichromatic vision to primates. Nature 2001, 410, 363–366. [Google Scholar] [CrossRef]
- Fornalé, F.; Vaglio, S.; Spiezio, C.; Prato Previde, E. Red-green colour vision in three catarrhine primates. Commun. Integr. Biol. 2012, 5, 583–589. [Google Scholar] [CrossRef] [Green Version]
- Gerald, M.S. How color may guide the primate world: Possible relationships between sexual selection and sexual dichromatism. In Sexual Selection and Reproductive Competition in Primates: New Perspectives and Directions; Jones, C.B., Ed.; American Society of Primatologists Special Topics in Primatology: Oklahoma City, OK, USA, 2003. [Google Scholar]
- Porter, R.H.; Moore, J.D. Human kin recognition by olfactory cues. Physiol. Behav. 1981, 27, 493–495. [Google Scholar] [CrossRef]
- Geissman, T.; Hulftegger, A.M. Olfactory communication in gibbons? In Current Primatology: Social Development, Learning and Behaviour; Roeder, J.J., Thierry, B., Anderson, J.R., Herrenschmidt, N., Eds.; Université Louis Pasteur Press: Paris, France, 1994; pp. 199–206. [Google Scholar]
- Wedekind, C.; Seebeck, T.; Bettens, F.; Paepke, A.J. MHC-dependent mate preferences in humans. Proc. Biol. Sci. B 1995, 260, 245–249. [Google Scholar]
- Wedekind, C.; Füri, S. Body odour preferences in men and women: Do they aim for specific MHC combinations or simply heterozygosity? Proc. Biol. Sci. B 1997, 264, 1471–1479. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.; Siegel, M.I.; Bonar, C.J.; Bhatnagar, K.P.; Mooney, M.P.; Burrows, A.M.; Smith, M.A.; Maico, L.M. The existence of the vomeronasal organ in postnatal chimpanzees and evidence for its homology to that of humans. J. Anat. 2001, 198, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Jacob, S.; McClintock, M.K.; Zelano, B.; Ober, C. Paternally inherited HLA alleles are associated with women’s choice of male odor. Nat. Genet. 2002, 30, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Klailova, M.; Lee, P.C. Wild western lowland gorillas signal selectively using odor. PLoS ONE 2014, 9, e99554. [Google Scholar] [CrossRef]
- Masi, S.; Bouret, S. Odor signals in wild western lowland gorillas: An involuntary and extra-group communication hypothesis. Physiol. Behav. 2015, 145, 123–126. [Google Scholar] [CrossRef]
- Weiß, B.M.; Kücklich, M.; Thomsen, R.; Henkel, S.; Jänig, S.; Kulik, L.; Birkemeyer, C.; Widdig, A. Chemical composition of axillary odorants reflects social and individual attributes in rhesus macaques. Behav. Ecol. Sociobiol. 2018, 72, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunn, C.L. The evolution of exaggerated sexual swellings in primates and the graded signal hypothesis. Anim. Behav. 1999, 58, 229–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagel, M. The evolution of conspicuous oestrous advertisement in Old World monkeys. Anim. Behav. 1994, 47, 1333–1341. [Google Scholar] [CrossRef] [Green Version]
- Semple, S.; McComb, K. Perception of female reproductive state from vocal cues in a mammal species. Proc. R Soc. Lond. B 2000, 267, 707–712. [Google Scholar] [CrossRef] [Green Version]
- Zeller, A.C. Communication by sight and smell. In Primates Societies; Smuts, B.B., Cheney, D.L., Seyfarth, R.M., Wrangham, R.W., Struhsaker, T.T., Eds.; Chicago University Press: Chicago, IL, USA, 1987. [Google Scholar]
- Crewe, R.M.; Burger, B.V.; Le Roux, M.; Katsir, Z. Chemical constituents of the chest gland secretion of the thick-tailed galago (Galago crassicaudatus). J. Chem. Ecol. 1979, 5, 861–868. [Google Scholar] [CrossRef]
- DelBarco-Trillo, J.; Burkert, B.A.; Goodwin, T.E.; Drea, C.M. Night and day: The comparative study of strepsirrhine primates reveals socioecological and phylogenetic patterns in olfactory signals. J. Evol. Biol. 2011, 24, 82–98. [Google Scholar] [CrossRef]
- Hayes, R.; Morelli, T.; Wright, P. Anogenital gland secretions of Lemur catta and Propithecus verreauxi coquereli: A preliminary chemical examination. Am. J. Primatol. 2004, 63, 49–62. [Google Scholar] [CrossRef]
- Palagi, E.; Dapporto, L. Beyond odor discrimination: Demonstrating individual recognition by scent in Lemur catta. Chem. Senses 2006, 31, 437–443. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, E.A.; Fernandez-Duque, E.; Evans, S.; Hagey, L.R. Sex, age, and family differences in the chemical composition of owl monkey (Aotus nancymaae) subcaudal scent secretions. Am. J. Primatol. 2008, 70, 12–18. [Google Scholar] [CrossRef]
- Epple, G.; Belcher, A.M.; Kuderling, I.; Zeller, U.; Scolnick, L.; Greenfield, K.L.; Smith, A.B.I. Making sense out of scents: Species differences in scent glands, scent-marking behaviour, and scent-mark composition in the Callitrichidae. In Marmosets and Tamarins: Systematics, Behaviour, and Ecology; Rylands, A.B., Ed.; Oxford University Press: Oxford, UK, 1993. [Google Scholar]
- Poirier, A.C.; Waterhouse, S.J.; Dunn, J.C.; Smith, A.C. Scent-marks signal species, sex and reproductive status in tamarins (Saguinus spp., Neotropical Primates). Chem. Senses 2021, bjab008. [Google Scholar] [CrossRef]
- Smith, T.; Tomlinson, A.; Mlotkiewicz, J.; Abbott, D. Female marmoset monkeys (Callithrix jacchus) can be identified from the chemical composition of their scent marks. Chem. Senses 2001, 26, 449–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curtis, R.F.; Ballantine, J.A.; Keverne, E.B.; Bonsall, R.W.; Michael, R.P. Identification of primate sexual pheromones and properties of synthetic attractants. Nature 1971, 232, 396–398. [Google Scholar] [CrossRef]
- Setchell, J.; Vaglio, S.; Moggi-Cecchi, J.; Boscaro, F.; Calamai, L.; Knapp, L. Chemical composition of scent-gland secretions in an Old World monkey (Mandrillus sphinx): Influence of sex, male status, and individual identity. Chem. Senses 2010, 35, 205–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setchell, J.; Vaglio, S.; Abbot, K.; Moggi-Cecchi, J.; Boscaro, F.; Pieraccini, G.; Knapp, L. Odour signals major histocompatibility complex genotype in an Old World monkey. Proc. Biol. Sci. B 2011, 278, 274–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaglio, S.; Minicozzi, P.; Kessler, S.E.; Walker, D.; Setchell, J.M. Olfactory signals and fertility in olive baboons. Sci. Rep. 2021, 11, 8506. [Google Scholar] [CrossRef] [PubMed]
- Pochron, S.; Morelli, T.; Scirbona, J.; Wright, P. Sex differences in scent-marking in Propethicus edwardsi of ranomafana national park, Madagascar. Am. J. Primatol. 2005, 66, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Curtis, D.J.; Zaramody, A. Social structure and seasonal variation in the behaviour of Eulemur mongoz. Folia Primatol. 1999, 70, 79–96. [Google Scholar] [CrossRef] [PubMed]
- Heymann, E.W. Interspecific cariation of scent-marking behaviour in wild tamarins, Saguinus mystax and Saguinus fuscicollis. Folia Primatol. 2001, 72, 253–267. [Google Scholar] [CrossRef]
- Lewis, R.J. Sex differences in scent-marking in sifakas: Mating conflict or male services? Am. J. Phys. Anthropol. 2005, 128, 389–398. [Google Scholar] [CrossRef]
- Mertl-Millhollen, A.S. Scent marking as resource defense by female Lemur catta. Am. J. Primatol. 2006, 68, 605–621. [Google Scholar] [CrossRef] [PubMed]
- Lledo-Ferrer, Y.; Peláez, F.; Heymann, E.W. Can overmarking be considered as a means of chemical mate guarding in a wild callitrichid? Folia Primatol. 2010, 81, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Spence-Aizenberg, A.; Kimball, B.A.; Williams, L.E.; Fernandez-Duque, E. Chemical composition of glandular secretions from a pair-living monogamous primate: Sex, age, and gland differences in captive and wild owl monkeys (Aotus) spp. Am. J. Primatol. 2018, 80, e22730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fornasieri, I.; Roeder, J. Marking behaviour in two Lemur species (L. fulvus and L. macaco): Relation to social status, reproduction, aggression and environmental change. Folia Primatol. 1992, 59, 137–148. [Google Scholar] [CrossRef]
- Crawford, J.C.; Boulet, M.; Drea, C. Smelling wrong: Hormonal contraception in lemurs alters critical female odour cues. Proc. Biol. Sci. B 2011, 278, 122–130. [Google Scholar] [CrossRef] [Green Version]
- Drea, C.M.; Boulet, M.; DelBarco-Trillo, J.; Greene, L.K.; Sacha, C.R.; Goodwin, T.E.; Dubay, G.R. The “secret” in secretions: Methodological considerations in deciphering primate olfactory communication. Am. J. Primatol. 2013, 75, 621–642. [Google Scholar] [CrossRef] [PubMed]
- Belcher, A.; Epple, G.; Greenfield, K.L.; Richards, L.E.; Kuderling, I.; Smith, A.B. Proteins-biologically relevant components of the scent marks of a primate (Saguinus fuscicollis). Chem. Senses 1990, 15, 431–446. [Google Scholar] [CrossRef]
- DelBarco-Trillo, J.; Harelimana, I.H.; Goodwin, T.E.; Drea, C.M. Chemical differences between voided and bladder urine in the aye-aye (Daubentonia madagascariensis): Implications for olfactory communication studies. Am. J. Primatol. 2013, 75, 695–702. [Google Scholar] [CrossRef]
- Doty, R.L. Olfactory communication in humans. Chem. Senses 1981, 6, 351–376. [Google Scholar] [CrossRef]
- Rivera, A.J.; Stumpf, R.M.; Wilson, B.; Leigh, S.; Salyers, A.A. Baboon vaginal microbiota: An overlooked aspect of primate physiology. Am. J. Primatol. 2010, 72, 467–474. [Google Scholar] [CrossRef] [Green Version]
- Pener, M.P.; Simpson, S.J. Locust phase polyphenism: An update. Adv. Insect Physiol. 2009, 36, 1–272. [Google Scholar]
- Apps, P.J.; Weldon, P.J.; Kramer, M. Chemical signals in terrestrial vertebrates: Search for design features. Nat. Prod. Rep. 2015, 32, 1131–1153. [Google Scholar] [CrossRef] [PubMed]
- Drea, C.M. Design, delivery and perception of condition dependent chemical signals in strepsirrhine primates: Implications for human olfactory communication. Phil. Trans. R. Soc. B 2020, 375, 20190264. [Google Scholar] [CrossRef]
- Smith, T. Individual olfactory signatures in common marmosets (Callithrix jacchus). Am. J. Primatol. 2006, 68, 585–604. [Google Scholar] [CrossRef] [PubMed]
- Baker, M. Fur rubbing: Use of medicinal plants by capuchin monkeys (Cebus capucinus). Am. J. Primatol. 1996, 38, 263–270. [Google Scholar] [CrossRef]
- Poirier, A.C.; Waterhouse, J.S.; Watsa, M.; Erkenswick, G.A.; Moreira, L.A.A.; Tang, J.; Dunn, J.C.; Melin, A.D.; Smith, A.C. On the trail of primate scent signals: A field analysis of callitrichid scent-gland secretions by portable gas chromatography-mass spectrometry. Am. J. Primatol. 2021, e23236. [Google Scholar]
- Wyatt, T. Pheromones and Animal Behaviour. Chemical Signals and Signatures; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Ramirez, S.R.; Eltz, T.; Fujiwara, M.K.; Gerlach, G.; Goldman-Huertas, B.; Tsutsui, N.D.; Pierce, N.E. Asynchronous diversification in a specialized plant-pollinator mutualism. Science 2011, 333, 1742–1746. [Google Scholar] [CrossRef] [PubMed]
- Alborne, E.S. Mammalian Semiochemistry; John Wiley & Sons: New York, NY, USA, 1984. [Google Scholar]
- Hurst, J.L.; Robertson, D.; Tolladay, U.; Beynon, J. Proteins in urine scent marks of male house mice extend the longevity of olfactory signals. Anim. Behav. 1998, 55, 1289–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyatt, T. Proteins and peptides as pheromone signals and chemical signatures. Anim. Behav. 2014, 97, 273–280. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behaviour | Description |
|---|---|
| Scent-Marking: Ano-genital | Individual rubs its genital region against a conspecific, the substrate or object in the enclosure. |
| Scent-Marking: Head | Individual rubs its head against a conspecific, the substrate or object in the enclosure. |
| Scent-Marking: Wrist | Individual rubs its wrists or hands against a conspecific, the substrate or object in the enclosure. |
| Scent-Marking: Allomark | A scent-mark placed directly on the body of another individual using either ano-genital, head or wrist glands. |
| Scent-Marking: Countermark | A scent-mark placed onto the location another individual scent-marked within 30 s using either ano-genital, head or wrist glands. |
| Sniffing/Licking: Environment | Individual deliberately places its nostrils/tongue within 3 cm from the substrate or object and sniffs/licks |
| Sniffing/Licking: Conspecific | Individual deliberately places its nostrils/tongue within 3 cm from a conspecific and sniffs/licks. |
| Self-Licking | Individual uses its tongue to lick an area close to a scent-gland on its own body. |
| Deposition Location | Description |
|---|---|
| Branches | Natural structures in the enclosure consisting of logs, trees and various plant species. |
| Hatches | Small entrances that can be open or closed, linking the different enclosures together. |
| Climbing Frames | Man-made wooden structures in enclosure consisting of walkways, ropes and platforms. |
| Enrichment Devices | Removable objects that were placed daily to promote good welfare. |
| Ground | The artificial floor of the indoor enclosures and the grassy area of the outdoor enclosure. |
| Molecular Weight (Da) | Compound | Sex |
|---|---|---|
| 94.042 | Phenol | Female |
| 100.089 | Hexanal | Both |
| 106.042 | Benzaldehyde | Both |
| 106.078 | Benzene, 1,3-dimethyl- | Male |
| 107.063 | 4-Cyanocyclohexene | Female |
| 114.104 | Heptanal | Male |
| 118.099 | Ethanol, 2-butoxy- | Female |
| 108.058 | Benzyl alcohol | Female |
| 122.037 | Phenol, 4-ethyl- | Female |
| 126.104 | 5-Hepten-2-one, 6-methyl- | Male |
| 126.104 | 2-Octenal, (E)- | Male |
| 128.120 | 1-Octen-3-ol | Both |
| 128.120 | 3-Octanone | Female |
| 130.000 | 2-Ethyl- 1-Hexanol | Both |
| 136.125 | .beta.-Myrcene | Female |
| 136.125 | D-Limonene | Both |
| 136.125 | .beta.-Ocimene | Female |
| 136.158 | .alpha.-Pinene | Female |
| 138.104 | Furan, 2-pentyl- | Male |
| 142.136 | Nonanal | Both |
| 152.047 | Methyl salicylate | Female |
| 154.136 | Linalool | Female |
| 154.136 | Terpinen-4-ol | Female |
| 154.136 | α-Terpineol | Female |
| 156.151 | 7-Octen-2-ol, 2,6-dimethyl- | Female |
| 156.151 | Decanal | Both |
| 158.167 | 1-Decanol | Female |
| 182.203 | Cyclohexane, 2-butyl-1,1,3-trimethyl- | Female |
| 184.119 | Undecane, 2,6-dimethyl- | Male |
| 184.219 | Dodecane, 4-methyl- | Male |
| 196.143 | Linalyl acetate | Male |
| 196.146 | 4-Hexen-1-ol, 5-methyl-2-(1-methylethenyl)-, acetate | Female |
| 198.235 | Tridecane, 6-methyl- | Female |
| 214.099 | Benzene, 1,1′-[1,2-ethanediylbis(oxy)]bis- | Male |
| 248.114 | 2-Bromo dodecane | Female |
| 286.214 | 2,2,4-Trimethyl-1,3-pentanediol diisobutyrate | Male |
| 356.329 | Carbonic acid, decyl undecyl ester | Both |
| 426.407 | Carbonic acid, decyl hexadecyl ester | Male |
| - | Unknown hydrocarbon 01 | Male |
| - | Unknown hydrocarbon 02 | Male |
| - | Unknown hydrocarbon 03 | Male |
| - | Unknown hydrocarbon 04 | Male |
| - | Unknown hydrocarbon 05 | Male |
| - | Unknown hydrocarbon 06 | Male |
| - | Unknown hydrocarbon 07 | Male |
| - | Unknown hydrocarbon 08 | Male |
| - | Unknown hydrocarbon 09 | Male |
| - | Unknown hydrocarbon 10 | Male |
| - | Unknown hydrocarbon 11 | Male |
| - | Unknown hydrocarbon 12 | Male |
| - | Unknown hydrocarbon 13 | Male |
| - | Unknown hydrocarbon 14 | Male |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elwell, E.J.; Walker, D.; Vaglio, S. Sexual Dimorphism in Crowned Lemur Scent-Marking. Animals 2021, 11, 2091. https://doi.org/10.3390/ani11072091
Elwell EJ, Walker D, Vaglio S. Sexual Dimorphism in Crowned Lemur Scent-Marking. Animals. 2021; 11(7):2091. https://doi.org/10.3390/ani11072091
Chicago/Turabian StyleElwell, Emily J., David Walker, and Stefano Vaglio. 2021. "Sexual Dimorphism in Crowned Lemur Scent-Marking" Animals 11, no. 7: 2091. https://doi.org/10.3390/ani11072091