
Sensor and Video: Two Complementary Approaches for Evaluation of Dairy Cow Behavior after Calving Sensor Attachment


Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Sensor Systems to Improve Calving Management
1.2. How Can Behavioral Changes in Dairy Cows Equipped with Tail-Attached Calving Sensors Be Assessed?
2. Materials and Methods
2.1. Data Collection
2.1.1. Animals and Housing
2.1.2. Sensors and Calving Management
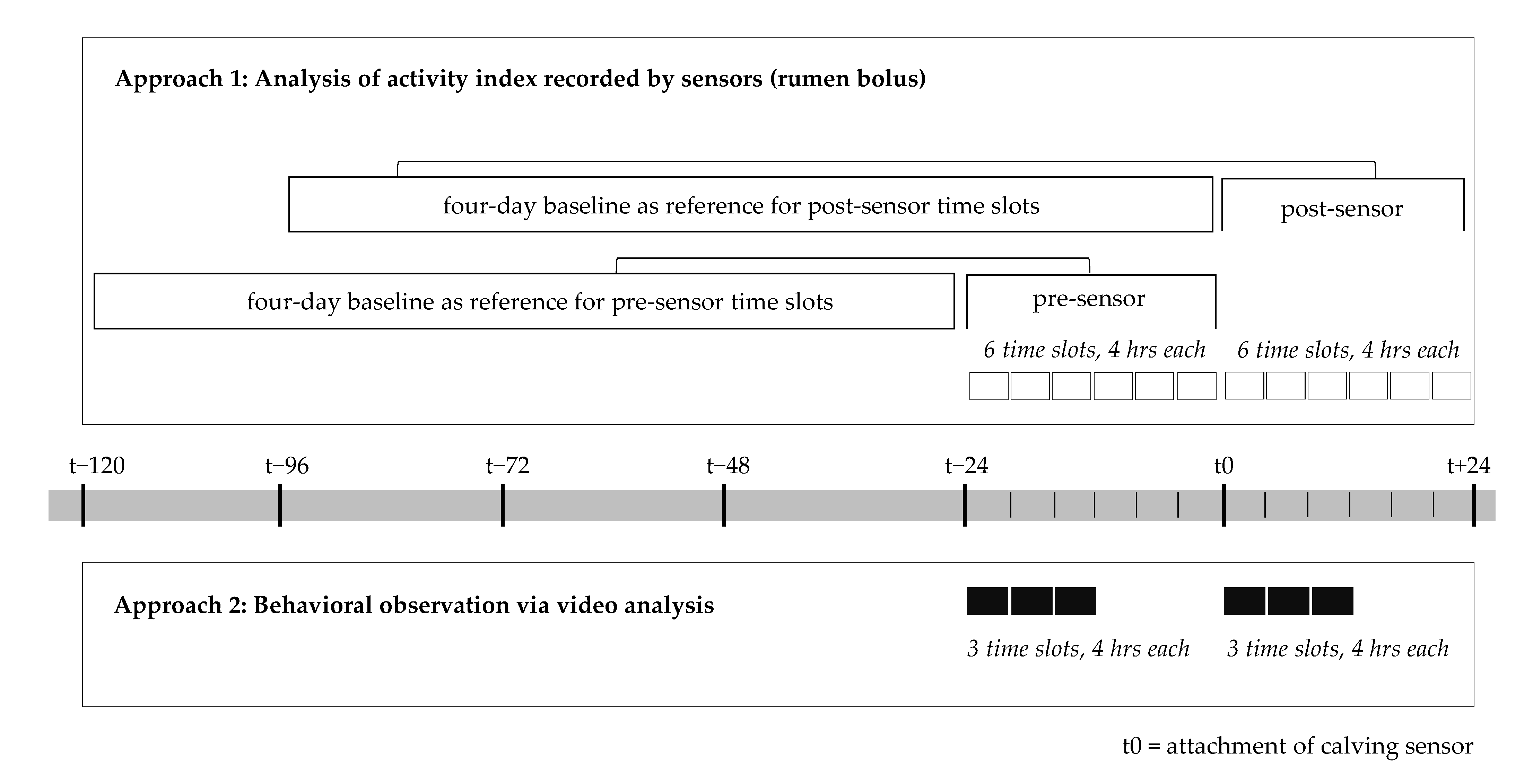
2.2. Methodological Approach 1: Analysis of the Activity Index Recorded by Sensors (Rumen Bolus)
2.3. Methodological Approach 2: Behavioral Observation via Video Analysis
3. Results and Discussion
3.1. Documentation Concerning Calving Sensor Attachment, Abnormalities after Calving Sensor Attachment, and Health
- In cow 2, conspicuous activity behavior was observed immediately after attaching the sensor (CalveSense) to the tail. She rubbed her tail heavily on the water trough for the first 15 min, although this decreased afterwards (also observed in video).
- About an hour after the calving sensor (Moocall) was attached to cow 7, the fit of the sensor on the tail had to be readjusted. The cow was fixed in the feed fence for a short time and the sensor was reattached (attachment: 08:24 a.m.; reattachment: 09:35 a.m.).
- Cow 15 showed discomfort in her activity behavior 23 h and 15 min after attachment of the calving sensor (Moocall). As pressure points and slight swelling were visible on the tail, the sensor was removed immediately (no video available).
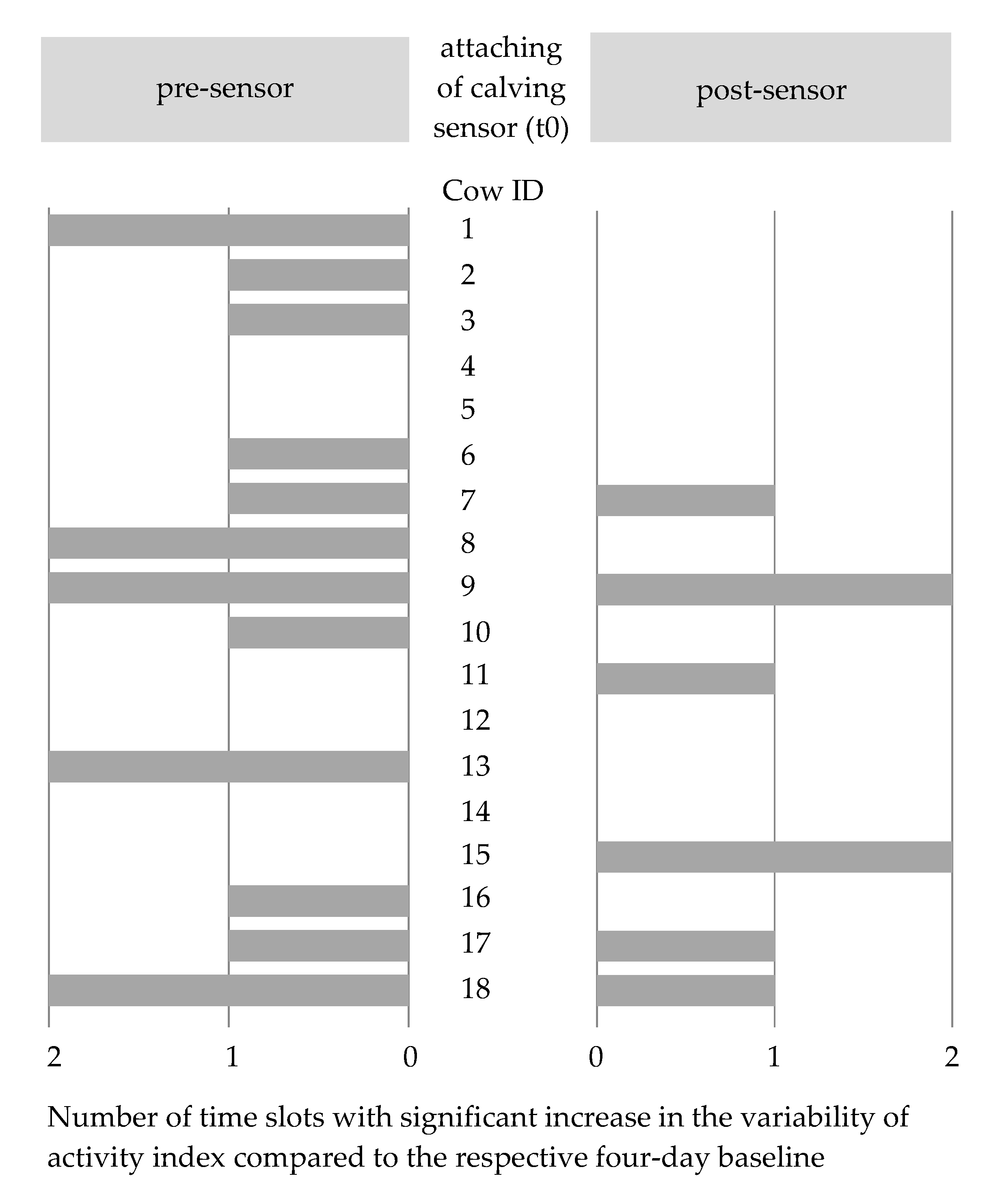
3.2. Methodological Approach 1: Changes in the Variability of the Activity Index
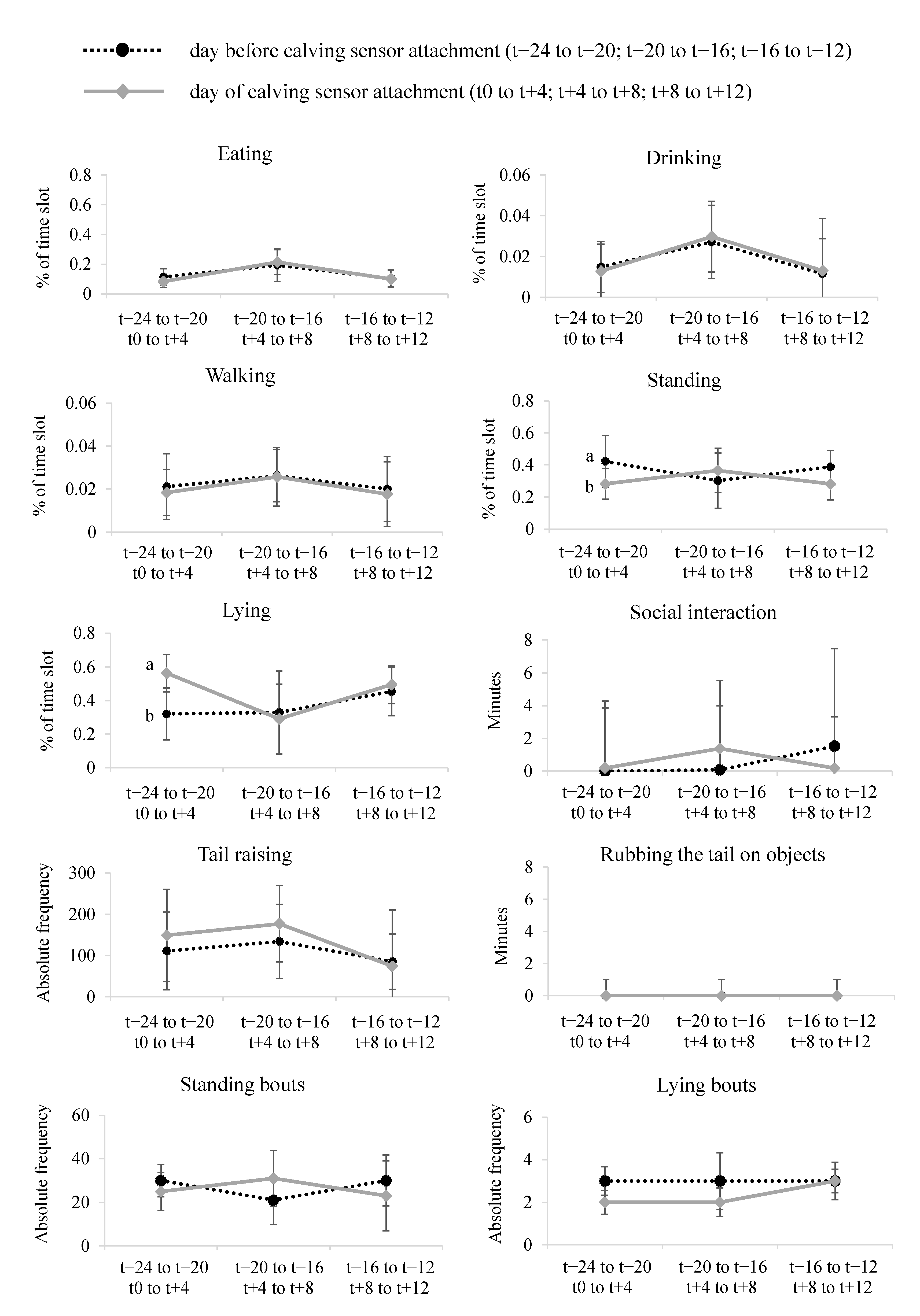
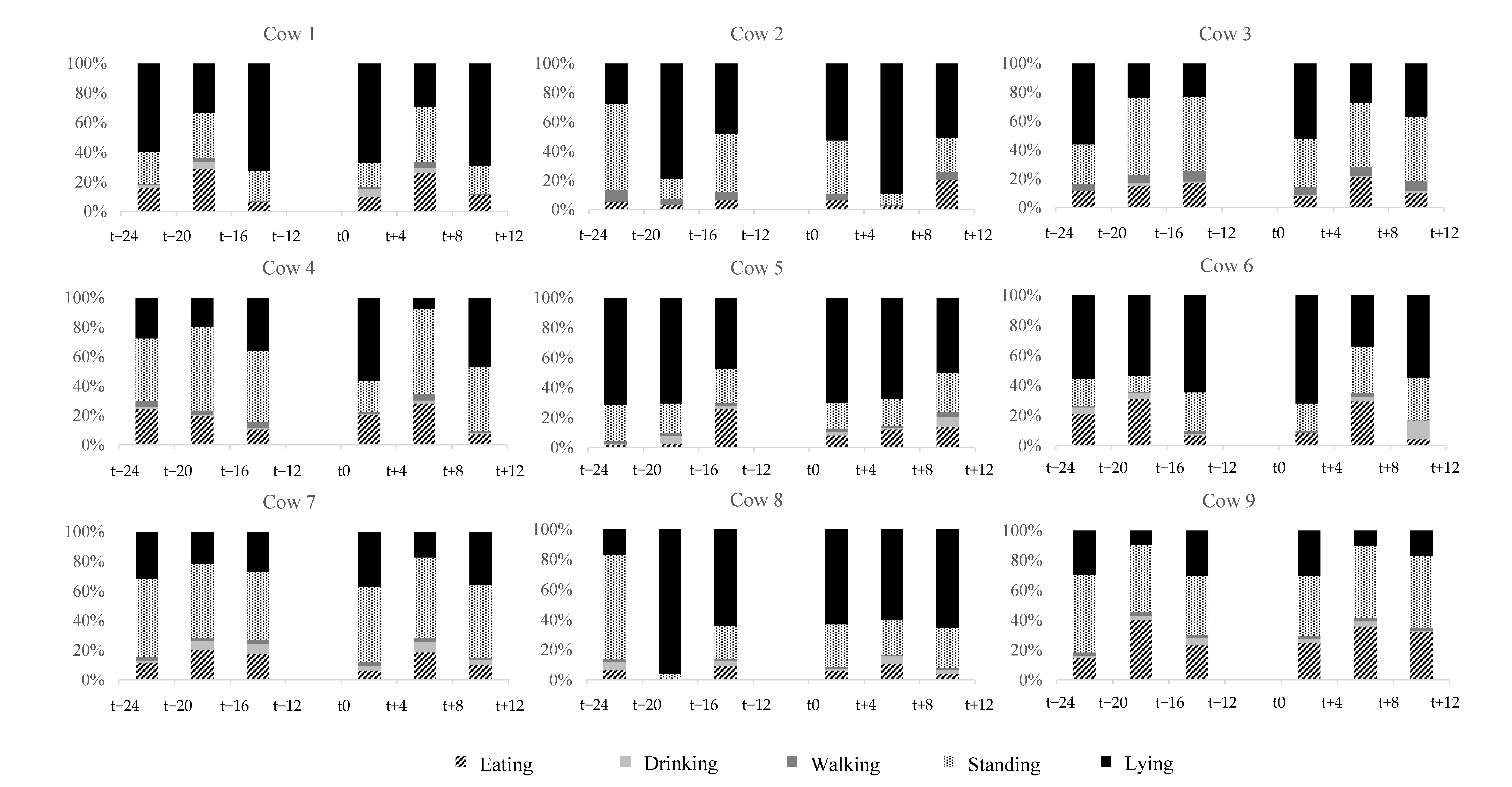
3.3. Methodological Approach 2: Behavioral Observation via Video Analysis
3.3.1. Standing, Walking, Lying, Eating, Drinking
3.3.2. Tail Raising and Rubbing the Tail on Objects
3.3.3. Social Interaction
3.4. General Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cow ID | Breed | Age [years] | Parity | Type of Sensor | Included in Methodological Approach |
|---|---|---|---|---|---|
| 1 | Simmental | 4.1 | 3 | Moocall | 1 a and 2 b |
| 2 | Simmental | 2.3 | 1 | CalveSense | 1 and 2 |
| 3 | Brown-Swiss | 2.1 | 1 | CalveSense | 1 and 2 |
| 4 | Simmental | 2.4 | 1 | CalveSense | 1 and 2 |
| 5 | Simmental | 10.4 | 8 | CalveSense | 1 and 2 |
| 6 | Holstein | 9.3 | 8 | CalveSense | 1 and 2 |
| 7 | Simmental | 8.6 | 7 | Moocall | 1 and 2 |
| 8 | Simmental | 4.3 | 3 | Moocall | 1 and 2 |
| 9 | Brown-Swiss | 5.5 | 4 | CalveSense | 1 and 2 |
| 10 | Brown-Swiss | 4.2 | 3 | CalveSense | 1 |
| 11 | Simmental | 4.3 | 3 | CalveSense | 1 |
| 12 | Simmental | 10.1 | 9 | CalveSense | 1 |
| 13 | Holstein | 6.6 | 5 | Moocall | 1 |
| 14 | Simmental | 6.8 | 5 | CalveSense | 1 |
| 15 | Simmental | 5.1 | 4 | Moocall | 1 |
| 16 | Simmental | 4.6 | 3 | CalveSense | 1 |
| 17 | Holstein | 4.2 | 3 | CalveSense | 1 |
| 18 | Brown-Swiss | 4.6 | 3 | CalveSense | 1 |
| Cow ID | Breed | Time Sensor Was Attached | Type of Sensor | Item | Baseline b (Pre-Sensor) | Baseline c (Post-Sensor) | t − 24 to t − 20 | t − 20 to t − 16 | t − 16 to t − 12 | t − 12 to t − 8 | t − 8 to t − 4 | t − 4 to t0 | t0 to t + 4 | t + 4 to t + 8 | t + 8 to t + 12 | t + 12 to t + 16 | t + 16 to t + 20 | t + 20 to t + 24 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | S | 09:15 | M | MAD | 0.66 | 0.73 | 1.10 | 0.69 | 0.43 | 0.98 | 0.62 | 0.55 | 0.44 | 0.60 | 0.53 | 0.26 | 0.76 | 0.33 |
| median | −0.01 | 0.01 | 0.59 | −0.09 | −0.37 | 0.06 | −0.05 | 0.25 | 0.12 | −0.10 | −0.05 | 0.04 | −0.07 | 0.12 | ||||
| BF a | sig.I | sig.I | sig.D | sig.D | ||||||||||||||
| 2 | S | 19:20 | C | MAD | 0.66 | 0.65 | 1.00 | 0.50 | 0.52 | 0.60 | 0.73 | 0.59 | 0.86 | 0.50 | 0.67 | 0.55 | 0.48 | 0.29 |
| median | 0.02 | 0.01 | −0.37 | −0.21 | 0.18 | 0.13 | 0.11 | −0.09 | −0.19 | 0.12 | 0.35 | 0.28 | −0.04 | −0.03 | ||||
| BF | sig.I | sig.D | ||||||||||||||||
| 3 | BS | 09:50 | C | MAD | 0.79 | 0.77 | 0.97 | 1.28 | 0.51 | 0.55 | 0.42 | 0.58 | 0.97 | 0.90 | 0.70 | 0.63 | 0.55 | 0.60 |
| median | 0.03 | −0.03 | −0.10 | 0.39 | −0.27 | −0.34 | −0.04 | 0.06 | 0.03 | 0.15 | −0.20 | 0.12 | −0.21 | 0.23 | ||||
| BF | sig.I | sig.D | ||||||||||||||||
| 4 | S | 08:50 | C | MAD | 0.62 | 0.61 | 0.68 | 0.68 | 0.47 | 0.46 | 0.81 | 0.50 | 0.56 | 0.77 | 0.70 | 0.51 | 0.58 | 0.45 |
| median | 0.03 | −0.01 | −0.10 | 0.30 | −0.24 | −0.20 | 0.30 | −0.46 | 0.17 | 0.20 | −0.38 | 0.12 | −0.22 | −0.02 | ||||
| BF | ||||||||||||||||||
| 5 | S | 08:20 | C | MAD | 0.54 | 0.54 | 0.36 | 0.52 | 0.38 | 0.33 | 0.44 | 0.55 | 0.51 | 0.49 | 0.75 | 0.32 | 0.42 | 0.38 |
| median | −0.04 | −0.02 | 0.01 | −0.13 | 0.12 | −0.04 | −0.02 | −0.01 | −0.11 | 0.06 | 0.24 | −0.05 | 0.10 | −0.05 | ||||
| BF | ||||||||||||||||||
| 6 | H | 08:50 | C | MAD | 0.36 | 0.35 | 0.28 | 0.54 | 0.27 | 0.45 | 0.12 | 0.35 | 0.21 | 0.37 | 0.29 | 0.27 | 0.34 | 0.19 |
| median | −0.03 | −0.03 | 0.24 | −0.16 | −0.06 | 0.09 | −0.07 | 0.09 | 0.09 | 0.09 | −0.20 | −0.19 | −0.03 | −0.04 | ||||
| BF | sig.I | sig.D | sig.D | |||||||||||||||
| 7 | S | 08:24 | M | MAD | 0.53 | 0.55 | 0.39 | 0.31 | 0.32 | 0.41 | 0.55 | 0.94 | 0.87 | 0.66 | 0.36 | 0.48 | 0.69 | 0.50 |
| median | 0.00 | 0.01 | 0.11 | −0.24 | 0.13 | −0.03 | −0.02 | 0.05 | −0.11 | 0.02 | 0.15 | 0.20 | −0.20 | 0.10 | ||||
| BF | sig.I | sig.I | ||||||||||||||||
| 8 | S | 17:55 | M | MAD | 0.82 | 0.82 | 1.44 | 0.65 | 0.79 | 1.03 | 0.70 | 1.28 | 0.71 | 0.90 | 0.54 | 0.82 | 0.96 | 0.77 |
| median | −0.01 | −0.01 | −0.21 | −0.02 | −0.01 | 0.02 | −0.38 | −0.33 | −0.31 | −0.25 | −0.23 | 0.09 | 0.03 | −0.19 | ||||
| BF | sig.I | sig.I | ||||||||||||||||
| 9 | BS | 08:25 | C | MAD | 0.97 | 1.06 | 0.34 | 0.50 | 1.10 | 0.66 | 2.11 | 1.92 | 0.78 | 0.27 | 0.50 | 1.70 | 0.61 | 2.05 |
| median | 0.01 | 0.03 | 0.17 | −0.145 | 0.25 | −0.09 | 0.56 | −0.87 | −0.20 | 0.04 | −0.02 | −0.29 | −0.11 | −0.69 | ||||
| BF | sig.D | sig.I | sig.I | sig.D | sig.I | sig.I | ||||||||||||
| 10 | BS | 09:00 | C | MAD | 0.84 | 0.86 | 0.67 | 0.73 | 2.05 | 0.73 | 0.93 | 0.68 | 0.97 | 0.85 | 0.46 | 0.67 | 0.52 | 0.65 |
| median | 0.01 | −0.07 | −0.08 | −0.32 | −0.46 | −0.43 | −0.30 | 0.05 | −0.06 | 0.05 | −0.24 | 0.00 | 0.45 | −0.31 | ||||
| BF | sig.I | sig.D | ||||||||||||||||
| 11 | S | 18:45 | C | MAD | 0.52 | 0.49 | 0.34 | 0.38 | 0.31 | 0.50 | 0.71 | 0.59 | 0.39 | 0.46 | 0.46 | 0.48 | 0.63 | 0.73 |
| median | −0.04 | −0.02 | −0.18 | 0.06 | 0.20 | −0.10 | −0.16 | −0.22 | −0.05 | 0.11 | −0.10 | −0.10 | 0.28 | −0.47 | ||||
| BF | sig.I | |||||||||||||||||
| 12 | S | 18:30 | C | MAD | 0.41 | 0.41 | 0.49 | 0.25 | 0.30 | 0.36 | 0.21 | 0.48 | 0.33 | 0.45 | 0.24 | 0.47 | 0.28 | 0.34 |
| median | −0.01 | 0.00 | −0.18 | 0.18 | −0.16 | 0.16 | 0.01 | −0.05 | −0.13 | −0.06 | 0.20 | −0.12 | 0.31 | −0.10 | ||||
| BF | sig.D | sig.D | ||||||||||||||||
| 13 | H | 17:45 | M | MAD | 0.35 | 0.37 | 0.59 | 0.55 | 0.29 | 0.33 | 0.31 | 0.42 | 0.41 | 0.26 | 0.50 | 0.29 | 0.33 | 0.29 |
| median | 0.02 | 0.01 | 0.11 | −0.14 | 0.04 | −0.03 | −0.09 | 0.15 | −0.08 | −0.07 | −0.11 | −0.04 | −0.13 | 0.04 | ||||
| BF | sig.I | sig.I | ||||||||||||||||
| 14 | S | 10:00 | C | MAD | 0.38 | 0.38 | 0.21 | 0.46 | 0.41 | 0.27 | 0.30 | 0.23 | 0.34 | 0.28 | 0.27 | 0.50 | 0.32 | 0.34 |
| median | 0.01 | 0.01 | 0.20 | −0.01 | 0.11 | −0.22 | 0.04 | −0.10 | 0.01 | 0.12 | −0.12 | −0.18 | 0.03 | 0.12 | ||||
| BF | ||||||||||||||||||
| 15 | S | 08:50 | M | MAD | 0.48 | 0.46 | 0.45 | 0.55 | 0.32 | 0.53 | 0.24 | 0.37 | 0.86 | 0.57 | 0.45 | 0.40 | 0.34 | 0.75 |
| median | 0.01 | 0.01 | 0.00 | −0.03 | 0.04 | 0.15 | 0.01 | 0.07 | −0.27 | 0.17 | 0.14 | −0.30 | −0.09 | 0.09 | ||||
| BF | sig.D | sig.I | sig.I | |||||||||||||||
| 16 | S | 19:20 | C | MAD | 0.56 | 0.54 | 1.09 | 0.35 | 0.25 | 0.41 | 0.53 | 0.41 | 0.37 | 0.32 | 0.41 | 0.47 | 0.39 | 0.43 |
| median | 0.00 | 0.00 | −0.32 | −0.11 | 0.08 | 0.10 | −0.02 | 0.06 | −0.03 | −0.01 | 0.06 | 0.04 | 0.07 | 0.01 | ||||
| BF | sig.I | sig.D | ||||||||||||||||
| 17 | H | 10:15 | C | MAD | 0.56 | 0.60 | 0.38 | 0.69 | 0.86 | 0.68 | 0.75 | 0.51 | 0.49 | 0.39 | 1.07 | 0.55 | 0.43 | 0.70 |
| median | 0.02 | 0.06 | 0.25 | 0.19 | 0.27 | 0.18 | −0.28 | 0.39 | 0.07 | −0.17 | 0.25 | −0.31 | 0.07 | 0.20 | ||||
| BF | sig.I | sig.I | ||||||||||||||||
| 18 | BS | 17:45 | C | MAD | 0.81 | 0.87 | 1.63 | 0.47 | 0.82 | 1.02 | 1.04 | 1.28 | 0.86 | 0.51 | 0.73 | 1.08 | 1.19 | 1.77 |
| median | −0.04 | −0.07 | −0.5 | 0.25 | 0.12 | 0.01 | −0.28 | 0.35 | −0.39 | −0.14 | 0.22 | 0.43 | −0.69 | −0.26 | ||||
| BF | sig.I | sig.I | sig.I |

References
- Maroto Molina, F.; Pérez Marín, C.C.; Molina Moreno, L.; Agüera Buendía, E.I.; Pérez Marín, D.C. Welfare Quality® for dairy cows: Towards a sensor-based assessment. J. Dairy Res. 2020, 87, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Ratnakaran, A.P.; Sejian, V.; Jose, V.S.; Vaswani, S.; Bagath, M.; Krishnan, G.; Beena, V.; Indira Devi, P.; Varma, G.; Bhatta, R. Behavioral responses to livestock adaptation to heat stress challenges. Asian J. Anim. Sci. 2017, 11, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Saint-Dizier, M.; Chastant-Maillard, S. Methods and on-farm devices to predict calving time in cattle. Vet. J. 2015, 205, 349–356. [Google Scholar] [CrossRef] [PubMed]
- Miedema, H.M.; Cockram, M.S.; Dwyer, C.M.; Macrae, A.I. Changes in the behaviour of dairy cows during the 24 h before normal calving compared with behaviour during late pregnancy. Appl. Anim. Behav. Sci. 2011, 131, 8–14. [Google Scholar] [CrossRef]
- Giaretta, E.; Marliani, G.; Postiglione, G.; Magazzù, G.; Pantò, F.; Mari, G.; Formigoni, A.; Accorsi, P.A.; Mordenti, A. Calving time identified by the automatic detection of tail movements and rumination time, and observation of cow behavioural changes. Animal 2021, 15, 100071. [Google Scholar] [CrossRef]
- Paolucci, M.; Di Giambattista, A.; Sylla, L.; Menichelli, M.; Banchio, A.; Monaci, M. Predicting time of parturition in Holstein Friesian cows by using C6 Birth Control. Reprod. Domest. Anim. 2008, 43, 57–58. [Google Scholar] [CrossRef]
- Marchesi, G.; Leonardi, S.; Tangorra, F.M.; Calcante, A.; Beretta, E.; Pofcher, E.; Lazzari, M. Evaluation of an electronic system for automatic calving detection on a dairy farm. Anim. Prod. Sci. 2013, 53, 1112–1114. [Google Scholar] [CrossRef]
- Voss, A.L.; Fischer-Tenhagen, C.; Bartel, A.; Heuwieser, W. Sensitivity and specificity of a tail-activity measuring device for calving prediction in dairy cattle. J. Dairy Sci. 2021, 104, 3353–3363. [Google Scholar] [CrossRef]
- Horváth, A.; Lénárt, L.; Csepreghy, A.; Madar, M.; Pálffy, M.; Szenci, O. A field study using different technologies to detect calving at a large-scale hungarian dairy farm. Reprod. Domest. Anim. 2021, 56, 673–679. [Google Scholar] [CrossRef]
- Lind, A.K.; Lindahl, C. Calving alert system—A helping technique or a welfare problem? In Proceedings of the 9th European Conference on Precision Livestock Farming, Cork, Ireland, 26–29 August 2019; pp. 385–388. [Google Scholar]
- Rushen, J.; Boissy, A.; Terlouw, E.M.C.; De Passillé, A.M.B. Opioid peptides and behavioral and physiological responses of dairy cows to social isolation in unfamiliar surroundings. J. Anim. Sci. 1999, 77, 2918–2924. [Google Scholar] [CrossRef] [Green Version]
- Rushen, J.; Munksgaard, L.; Marnet, P.G.; DePassillé, A.M. Human contact and the effects of acute stress on cows at milking. Appl. Anim. Behav. Sci. 2001, 73, 1–14. [Google Scholar] [CrossRef]
- Müller, R.; Schrader, L. Behavioural consistency during social separation and personality in dairy cows. Behaviour 2005, 142, 1289–1306. [Google Scholar] [CrossRef]
- Boissy, A.; Le Neindre, P. Behavioral, cardiac and cortisol responses to brief peer separation and reunion in cattle. Physiol. Behav. 1997, 61, 693–699. [Google Scholar] [CrossRef]
- Kovács, L.; Kézér, F.L.; Póti, P.; Jurkovich, V.; Szenci, O.; Nagy, K. Heart rate variability, step, and rumination behavior of dairy cows milked in a rotary milking system. J. Dairy Sci. 2019, 102, 5525–5529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenzel, C.; Schönreiter-Fischer, S.; Unshelm, J. Studies on step–kick behavior and stress of cows during milking in an automatic milking system. Livest. Prod. Sci. 2003, 83, 237–246. [Google Scholar] [CrossRef]
- Winter, A.; Hillerton, J.E. Behaviour associated with feeding and milking of early lactation cows housed in an experimental automatic milking system. Appl. Anim. Behav. Sci. 1995, 46, 1–15. [Google Scholar] [CrossRef]
- Gorewit, R.C.; Scott, N.R.; Czarniecki, C.S. Responses of dairy cows to alternating electrical current administered semirandomly in a nonavoidance environment. J. Dairy Sci. 1985, 68, 718–725. [Google Scholar] [CrossRef]
- Reinemann, D.J.; Stetson, L.E.; LeMire, S.D. Comparison of dairy cow aversion to continuous and intermittent current. Trans. ASAE 2004, 47, 1257. [Google Scholar] [CrossRef]
- Rigalma, K.; Duvaux-Ponter, C.; Barrier, A.; Charles, C.; Ponter, A.A.; Deschamps, F.; Roussel, S. Medium-term effects of repeated exposure to stray voltage on activity, stress physiology, and milk production and composition in dairy cows. J. Dairy Sci. 2010, 93, 3542–3552. [Google Scholar] [CrossRef]
- Lefcourt, A.M.; Kahl, S.; Akers, R.M. Correlation of indices of stress with intensity of electrical shock for cows. J. Dairy Sci. 1986, 69, 833–842. [Google Scholar] [CrossRef]
- Ominski, K.H.; Kennedy, A.D.; Wittenberg, K.M.; Nia, S.M. Physiological and production responses to feeding schedule in lactating dairy cows exposed to short-term, moderate heat stress. J. Dairy Sci. 2002, 85, 730–737. [Google Scholar] [CrossRef]
- Abeni, F.; Galli, A. Monitoring cow activity and rumination time for an early detection of heat stress in dairy cow. Int. J. Biometeorol. 2017, 61, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Cook, N.B.; Mentink, R.L.; Bennett, T.B.; Burgi, K. The effect of heat stress and lameness on time budgets of lactating dairy cows. J. Dairy Sci. 2007, 90, 1674–1682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilatti, J.A.; Vieira, F.M.C.; Rankrape, F.; Vismara, E.S. Diurnal behaviors and herd characteristics of dairy cows housed in a compost-bedded pack barn system under hot and humid conditions. Animal 2019, 13, 399–406. [Google Scholar] [CrossRef] [Green Version]
- Heinicke, J.; Ibscher, S.; Belik, V.; Amon, T. Cow individual activity response to the accumulation of heat load duration. J. Therm. Biol. 2019, 82, 23–32. [Google Scholar] [CrossRef]
- Wise, M.E.; Armstrong, D.V.; Huber, J.T.; Hunter, R.; Wiersma, F. Hormonal alterations in the lactating dairy cow in response to thermal stress. J. Dairy Sci. 1988, 71, 2480–2485. [Google Scholar] [CrossRef]
- Schütz, K.E.; Rogers, A.R.; Poulouin, Y.A.; Cox, N.R.; Tucker, C.B. The amount of shade influences the behavior and physiology of dairy cattle. J. Dairy Sci. 2010, 93, 125–133. [Google Scholar] [CrossRef]
- Schütz, K.E.; Cox, N.R.; Tucker, C.B. A field study of the behavioral and physiological effects of varying amounts of shade for lactating cows at pasture. J. Dairy Sci. 2014, 97, 3599–3605. [Google Scholar] [CrossRef]
- Hillman, P.E.; Lee, C.N.; Willard, S.T. Thermoregulatory responses associated with lying and standing in heat stressed dairy cows. Trans. ASAE 2005, 48, 795–801. [Google Scholar] [CrossRef]
- Kadzere, C.T.; Murphy, M.R.; Silanikove, N.; Maltz, E. Heat stress in lactating dairy cows: A review. Livest. Prod. Sci. 2002, 77, 59–91. [Google Scholar] [CrossRef]
- Ouellet, V.; Vasseur, E.; Heuwieser, W.; Burfeind, O.; Maldague, X.; Charbonneau, É. Evaluation of calving indicators measured by automated monitoring devices to predict the onset of calving in Holstein dairy cows. J. Dairy Sci. 2016, 99, 1539–1548. [Google Scholar] [CrossRef] [Green Version]
- Edwards, J.L.; Tozer, P.R. Using activity and milk yield as predictors of fresh cow disorders. J. Dairy Sci. 2004, 87, 524–531. [Google Scholar] [CrossRef]
- Thorup, V.M.; Munksgaard, L.; Robert, P.E.; Erhard, H.W.; Thomsen, P.T.; Friggens, N.C. Lameness detection via leg-mounted accelerometers on dairy cows on four commercial farms. Animal 2015, 9, 1704–1712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapinal, N.; De Passille, A.M.; Rushen, J.; Wagner, S. Automated methods for detecting lameness and measuring analgesia in dairy cattle. J. Dairy Sci. 2010, 93, 2007–2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rushen, J.; Pombourcq, E.; de Passillé, A.M. Validation of two measures of lameness in dairy cows. Appl. Anim. Behav. Sci. 2007, 106, 173–177. [Google Scholar] [CrossRef]
- Alsaaod, M.; Römer, C.; Kleinmanns, J.; Hendriksen, K.; Rose-Meierhöfer, S.; Plümer, L.; Büscher, W. Electronic detection of lameness in dairy cows through measuring pedometric activity and lying behavior. Appl. Anim. Behav. Sci. 2012, 142, 134–141. [Google Scholar] [CrossRef]
- Van Nuffel, A.; Vangeyte, J.; Mertens, K.C.; Pluym, L.; De Campeneere, S.; Saeys, W.; Opsomer, G.; Van Weyenberg, S. Exploration of measurement variation of gait variables for early lameness detection in cattle using the GAITWISE. Livest. Sci. 2013, 156, 88–95. [Google Scholar] [CrossRef]
- Smaxtec 2017. Smaxtec Basic Bolus. Available online: https://smaxtec.com/en/smaxtec-system-in-detail/#boli (accessed on 1 July 2019).
- Oberegger, A.; Smaxtec Animal Care GmbH, Graz, Austria. Personal communication, 2019.
- Ammer, S.; Lambertz, C.; Gauly, M. Comparison of different measuring methods for body temperature in lactating cows under different climatic conditions. J. Dairy Res. 2016, 83, 165–172. [Google Scholar] [CrossRef]
- Antanaitis, R.; Juozaitienė, V.; Rutkauskas, A.; Televičius, M.; Stasiulevičiūtė, I. Reticulorumen temperature and pH as indicators of the likelihood of reproductive success. J. Dairy Res. 2018, 85, 23–26. [Google Scholar] [CrossRef]
- Sharpe, K.T.; Heins, B.J.; Buchanan, E.S.; Reese, M.H. Evaluation of solar photovoltaic systems to shade cows in a pasture-based dairy herd. J. Dairy Sci. 2020, 104, 2794–2806. [Google Scholar] [CrossRef]
- Stein, S. Activity-based heat detection with the smaXtec intraruminal bolus system. In Proceedings of the Conference on Precision Dairy Farming, Lexington, KY, USA, 30 May–1 June 2017; pp. 63–66. [Google Scholar]
- Pfeiffer, J.; Gandorfer, M.; Ettema, J.F. Evaluation of activity meters for estrus detection: A stochastic bioeconomic modeling approach. J. Dairy Sci. 2020, 103, 492–506. [Google Scholar] [CrossRef]
- Fregonesi, J.A.; Leaver, J.D. Behaviour, performance and health indicators of welfare for dairy cows housed in strawyard or cubicle systems. Livest. Prod. Sci. 2001, 68, 205–216. [Google Scholar] [CrossRef]
- Walker, S.L.; Smith, R.F.; Routly, J.E.; Jones, D.N.; Morris, M.J.; Dobson, H. Lameness, activity time-budgets, and estrus expression in dairy cattle. J. Dairy Sci. 2008, 91, 4552–4559. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org (accessed on 28 April 2020).
- Trapletti, A.; Hornik, K. Tseries: Time Series Analysis and Computational Finance. R package version 0.10-48. 2020. Available online: https://CRAN.R-project.org/package=tseries (accessed on 28 October 2019).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 28 October 2019).
- Signorell, A.; Aho, L.; Alfons, A.; Anderegg, A.; Aragon, T.; Arachchige, C.; Arppe, A.; Baddeley, A.; Barton, K.; Bolker, B.; et al. DescTools: Tools for descriptive statistics. R package version 0.99.39. 2020. Available online: https://CRAN.R-project.org/package=DescTools (accessed on 28 October 2019).
- Jensen, M.B. Behaviour around the time of calving in dairy cows. Appl. Anim. Behav. Sci. 2012, 139, 195–202. [Google Scholar] [CrossRef]
- Rutten, C.J.; Kamphuis, C.; Hogeveen, H.; Huijps, K.; Nielen, M.; Steeneveld, W. Sensor data on cow activity, rumination, and ear temperature improve prediction of the start of calving in dairy cows. Comput. Electron. Agric. 2017, 132, 108–118. [Google Scholar] [CrossRef]
- Brown, M.B.; Forsythe, A.B. Robust tests for the equality of variances. J. Am. Stat. Assoc. 1974, 69, 364–367. [Google Scholar] [CrossRef]
- Olejnik, S.F.; Algina, J. Tests of variance equality when distributions differ in form and location. Educ. Psychol. Meas. 1988, 48, 317–329. [Google Scholar] [CrossRef]
- Conover, W.J.; Johnson, M.E.; Johnson, M.M. A comparative study of tests for homogeneity of variances, with applications to the outer continental shelf bidding data. Technometrics 1981, 23, 351–361. [Google Scholar] [CrossRef]
- Plourde, A.; Watkins, G.C. Crude oil prices between 1985 and 1994: How volatile in relation to other commodities? Resour. Energy Econ. 1998, 20, 245–262. [Google Scholar] [CrossRef]
- Artavia, M.; Deppermann, A.; Filler, G.; Grethe, H.; Häger, A.; Kirschke, D.; Odening, M. Ertrags-und Preisinstabilität auf Agrarmärkten in Deutschland und der EU [Yield and price instability in agricultural markets in the Germany and the EU]. Proc. Schr. Ges. Wirtsch. Soz. Landbaues eV 2011, 46, 259–274. [Google Scholar] [CrossRef]
- Gandorfer, M.; Porsch, A.; Bitsch, V. Producer price volatility in the German fruit and vegetable industry. Eur. J. Hortic. Sci. 2017, 82, 149–154. [Google Scholar] [CrossRef] [Green Version]
- Gastwirth, J.L. Screening and selection. In International Encyclopedia of Social Sciences; Smelser, N.J., Bates, P.B., Eds.; Elsevier: Oxford, UK, 2001; pp. 13755–13767. [Google Scholar]
- Pepe, M. The Statistical Evaluation of Medical Tests for Classification and Prediction; Wiley: Chichester, UK, 2003. [Google Scholar]
- Kvamme, K.L.; Stark, M.T.; Longacre, M.A. Alternative procedures for assessing standardization in ceramic assemblages. Am. Antiq. 1996, 61, 116–126. [Google Scholar] [CrossRef] [Green Version]
- Gastwirth, J.L.; Gel, Y.R.; Miao, W. The impact of Levene’s test of equality of variances on statistical theory and practice. Stat. Sci. 2009, 343–360. [Google Scholar] [CrossRef] [Green Version]
- Boudadi-Maligne, M.; Escarguel, G. A biometric re-evaluation of recent claims for Early Upper Palaeolithic wolf domestication in Eurasia. J. Archaeol. Sci. 2014, 45, 80–89. [Google Scholar] [CrossRef]
- Endres, M.I.; Barberg, A.E. Behavior of dairy cows in an alternative bedded-pack housing system. J. Dairy Sci. 2007, 90, 4192–4200. [Google Scholar] [CrossRef]
- Martiskainen, P.; Järvinen, M.; Skön, J.P.; Tiirikainen, J.; Kolehmainen, M.; Mononen, J. Cow behaviour pattern recognition using a three-dimensional accelerometer and support vector machines. Appl. Anim. Behav. Sci. 2009, 119, 32–38. [Google Scholar] [CrossRef]
- Rousing, T.; Wemelsfelder, F. Qualitative assessment of social behaviour of dairy cows housed in loose housing systems. Appl. Anim. Behav. Sci. 2006, 101, 40–53. [Google Scholar] [CrossRef]
- Bouissou, M.F.; Boissy, A.; Le Neindre, P.; Veissier, I. The social behaviour of cattle. In Social Behaviour in Farm Animals; Keeling, L.J., Gonyou, H.W., Eds.; CAB International: Wallingford, Oxon, UK, 2001; pp. 113–145. [Google Scholar]
- Nordlund, K.V.; Strassburg, P.; Bennett, T.B.; Oetzel, G.R.; Cook, N.B. Thermodynamics of standing and lying behavior in lactating dairy cows in freestall and parlor holding pens during conditions of heat stress. J. Dairy Sci. 2019, 102, 6495–6507. [Google Scholar] [CrossRef] [PubMed]
- Haley, D.B.; Rushen, J.; Passillé, A.D. Behavioural indicators of cow comfort: Activity and resting behaviour of dairy cows in two types of housing. Can. J. Anim. Sci. 2000, 80, 257–263. [Google Scholar] [CrossRef]
- Bewley, J.M.; Boyce, R.E.; Hockin, J.; Munksgaard, L.; Eicher, S.D.; Einstein, M.E.; Schutz, M.M. Influence of milk yield, stage of lactation, and body condition on dairy cattle lying behaviour measured using an automated activity monitoring sensor. J. Dairy Res. 2010, 77, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brzozowska, A.; Lukaszewicz, M.; Sender, G.; Kolasinska, D.; Oprzadek, J. Locomotor activities in dairy cows in relation to season and lactation. Appl. Anim. Behav. Sci. 2014, 156, 6–11. [Google Scholar] [CrossRef]
- Cooper, M.D.; Arney, D.R.; Phillips, C.J.C. Two-or four-hour lying deprivation on the behavior of lactating dairy cows. J. Dairy Sci. 2007, 90, 1149–1158. [Google Scholar] [CrossRef] [Green Version]
- Schütz, K.E.; Cave, V.M.; Cox, N.R.; Huddart, F.J.; Tucker, C.B. Effects of 3 surface types on dairy cattle behavior, preference, and hygiene. J. Dairy Sci. 2019, 102, 1530–1541. [Google Scholar] [CrossRef] [Green Version]
- Drenkard, D.H.; Gorewit, R.C.; Scott, N.R.; Sagi, R. Milk production, health, behavior, and endocrine responses of cows exposed to electrical current during milking. J. Dairy Sci. 1985, 68, 2694–2702. [Google Scholar] [CrossRef]
- Noble, W.S. How does multiple testing correction work? Nat. Biotechnol. 2009, 27, 1135–1137. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, R.A. When to use the Bonferroni correction. Ophthalmic Physiol. Opt. 2014, 34, 502–508. [Google Scholar] [CrossRef]
- Winckler, C.; Buehnemann, A.; Seidel, K. Social behaviour of commercial dairy herds as a parameter for on-farm welfare assessment. In Proceedings of the 36th International Congress of the ISAE, Wageningen, The Netherlands, 6–10 August 2002; p. 86. [Google Scholar]
- Galindo, F.; Broom, D.M. The effects of lameness on social and individual behavior of dairy cows. J. Appl. Anim. Welf. Sci. 2002, 5, 193–201. [Google Scholar] [CrossRef]
- Weary, D.M.; Niel, L.; Flower, F.C.; Fraser, D. Identifying and preventing pain in animals. Appl. Anim. Behav. Sci. 2006, 100, 64–76. [Google Scholar] [CrossRef] [Green Version]
- O’Callaghan, K.A.O.; Cripps, P.J.; Downham, D.Y.; Murray, R.D. Subjective and objective assessment of pain and discomfort due to lameness in dairy cattle. Anim. Welf. 2003, 12, 605–610. [Google Scholar]
- Gleerup, K.B.; Andersen, P.H.; Munksgaard, L.; Forkman, B. Pain evaluation in dairy cattle. Appl. Anim. Behav. Sci. 2015, 171, 25–32. [Google Scholar] [CrossRef] [Green Version]



| Unit | Behavior | Description |
|---|---|---|
| Duration of | Walking | The cow is moving all four legs (walking or running) [13] |
| Standing | The cow is standing without moving [13,25] | |
| Lying | The cow is lying in different natural lying positions [65] | |
| Eating | The cow places its head above the feeding table and searches, masticates or sorts the feed (silage) [66] | |
| Drinking | The cow places its head over the water trough [24] | |
| Frequency of | Tail raising 1 | Lateral > 90° |
| Duration of | Rubbing the tail on objects 1 | Rubbing the tail on objects (other cow in maternity pen, penning, or water trough) |
| Social interaction 1 | social licking: licking another cow’s head, neck, and/or shoulder areas sniffing head: head or muzzle stretched towards/maybe touching another cow’s head sniffing body: head or muzzle stretched towards/maybe touching another cow’s body gentle pushing: hard push of body against body head butting: blow with the forehead directed at another cow fighting: head-to-head pushing, sometimes followed by head to neck pushing and manoeuvring for position ([67], based on work from [68]) |
| Baseline a (Pre-Sensor) | Baseline b (Post-Sensor) | t − 24 to t − 20 | t − 20 to t − 16 | t − 16 to t − 12 | t − 12 to t − 8 | t − 8 to t − 4 | t − 4 to t0 | t0 to t + 4 | t + 4 to t + 8 | t + 8 to t + 12 | t + 12 to t + 16 | t + 16 to t + 20 | t + 20 to t + 24 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MAD | 0.16 | 0.17 | 0.23 | 0.25 | 0.19 | 0.16 | 0.09 | 0.30 | 0.24 | 0.15 | 0.12 | 0.11 | 0.13 | 0.23 |
| median | −0.02 | −0.02 | −0.05 | −0.06 | −0.06 | 0.09 | 0.01 | 0.01 | 0.01 | −0.05 | 0.01 | 0.01 | −0.04 | 0.05 |
| Brown–Forsythe test | sig.I | sig.I | sig.D | sig.I |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pfeiffer, J.; Spykman, O.; Gandorfer, M. Sensor and Video: Two Complementary Approaches for Evaluation of Dairy Cow Behavior after Calving Sensor Attachment. Animals 2021, 11, 1917. https://doi.org/10.3390/ani11071917
Pfeiffer J, Spykman O, Gandorfer M. Sensor and Video: Two Complementary Approaches for Evaluation of Dairy Cow Behavior after Calving Sensor Attachment. Animals. 2021; 11(7):1917. https://doi.org/10.3390/ani11071917
Chicago/Turabian StylePfeiffer, Johanna, Olivia Spykman, and Markus Gandorfer. 2021. "Sensor and Video: Two Complementary Approaches for Evaluation of Dairy Cow Behavior after Calving Sensor Attachment" Animals 11, no. 7: 1917. https://doi.org/10.3390/ani11071917