
High Stocking Density and Food Deprivation Increase Brain Monoaminergic Activity in Gilthead Sea Bream (Sparus aurata)

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Conditions
2.2. Sampling
2.3. Analytical Methods
2.3.1. Plasma Cortisol and Catecholamine Levels
2.3.2. Analysis of Brain Monoamines
2.4. Statistics
3. Results
3.1. Plasma Cortisol, Adrenaline, and Noradrenaline Levels
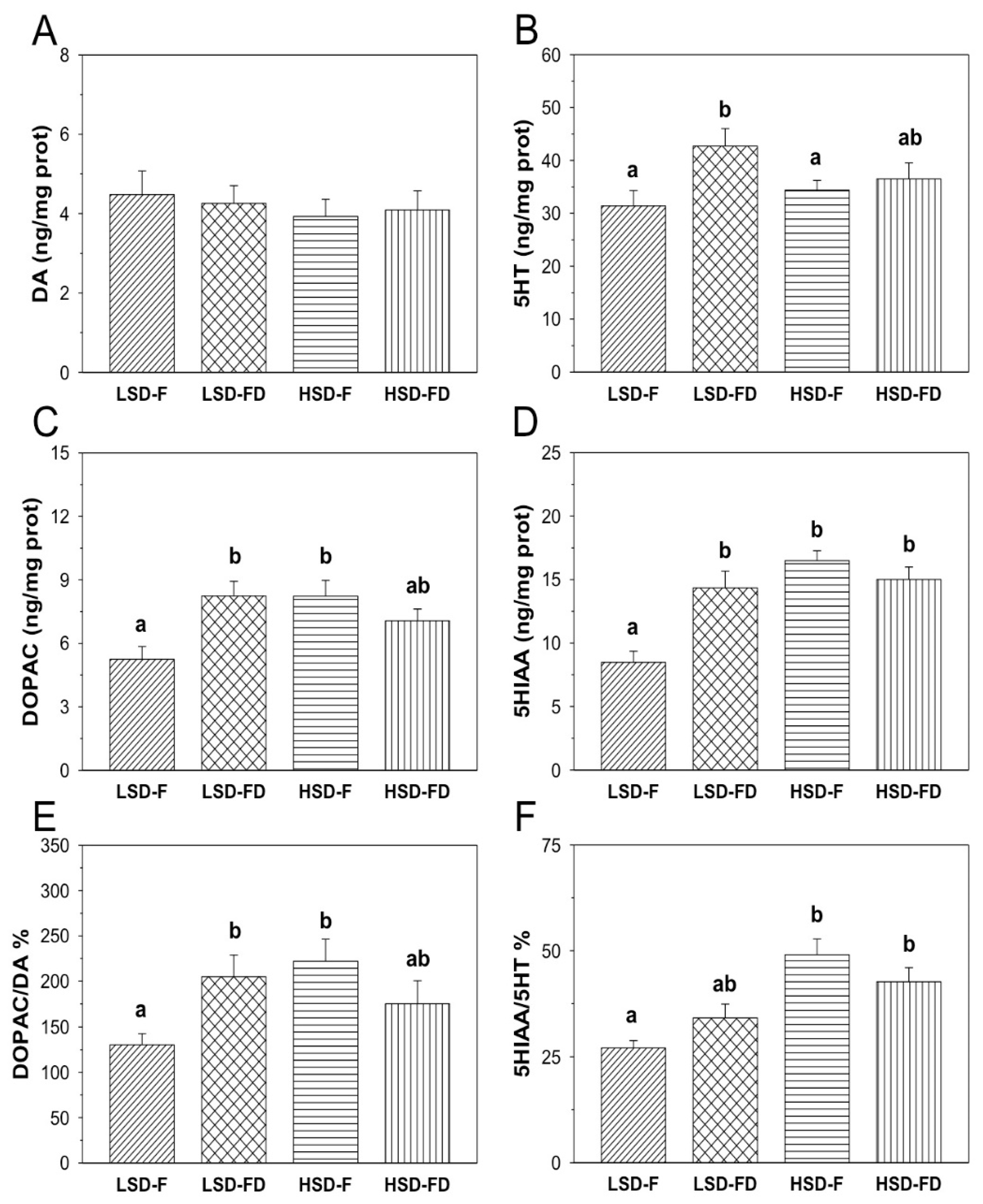
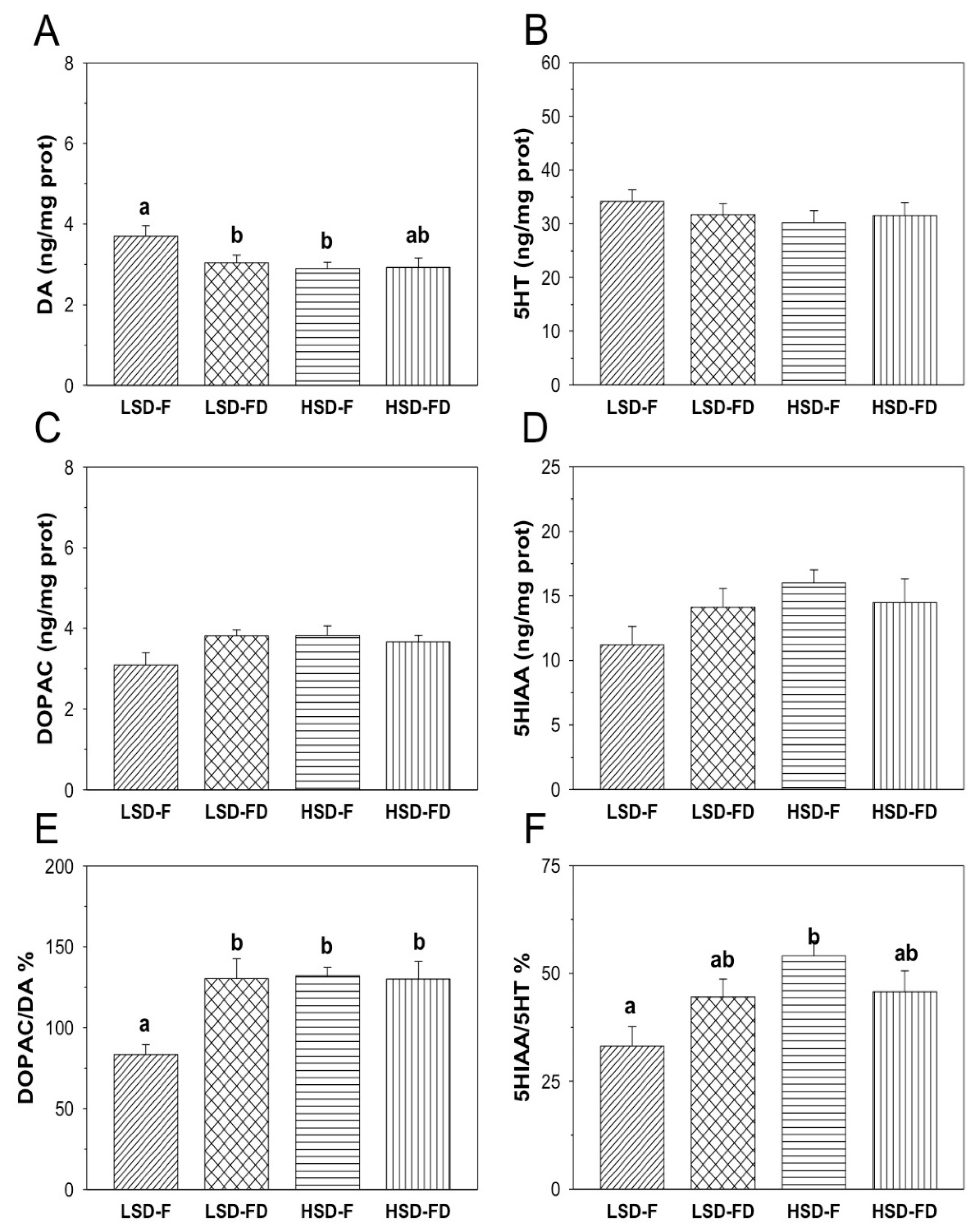
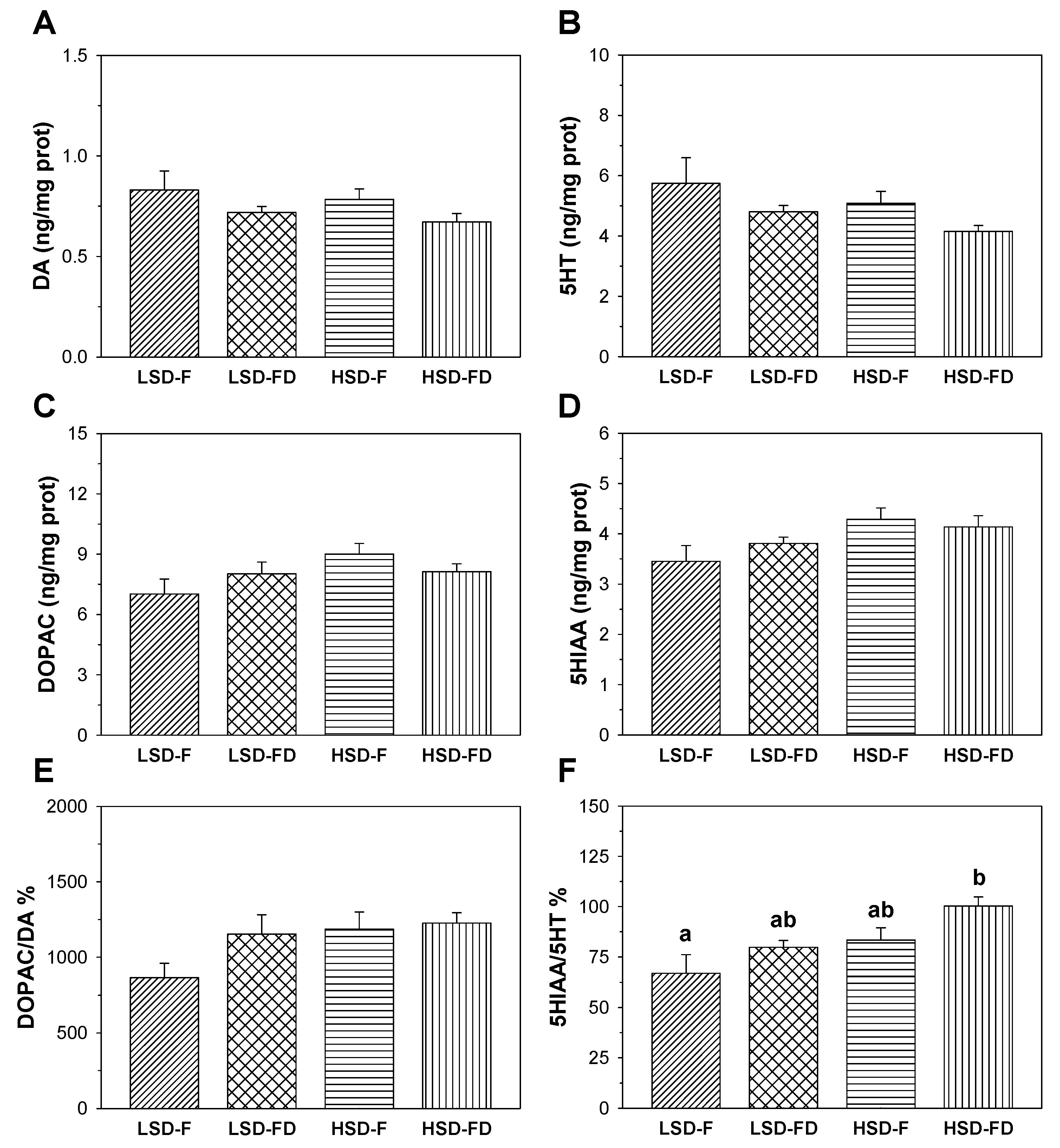
3.2. Brain Serotonergic and Dopaminergic Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Wendelaar Bonga, S.E. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef] [PubMed]
- Schreck, C.B.; Tort, L. The concept of stress in fish. In Biology of Stress in Fish; Fish Physiology Series 2016; Schreck, C.B., Tort, L., Farrell, A.T., Brauner, C.J., Eds.; Academic Press-Elsevier: Cambridge, MA, USA, 2016; Volume 35, pp. 2–34. [Google Scholar]
- Mommsen, T.P.; Vijayan, M.M.; Moon, T.W. Cortisol in teleosts: Dynamics, mechanisms of action, and metabolic regulation. Rev. Fish Biol. Fisher. 1999, 9, 211–268. [Google Scholar] [CrossRef]
- Iwama, G.K.; Afonso, L.O.B.; Vijayan, M.M. Stress in fishes. In The Physiology of Fishes; Evans, D.H., Claiborne, J.B., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 319–342. [Google Scholar]
- Ellis, T.; Yildiz, H.Y.; López-Olmeda, F.J.; Spedicato, M.T.; Tort, L.; Øverli, Ø.; Martins, C.I. Cortisol and finfish welfare. Fish Physiol. Biochem. 2012, 38, 163–188. [Google Scholar] [CrossRef] [PubMed]
- Gorissen, M.; Flik, G. The Endocrinology of the Stress Response in Fish: An Adaptation-Physiological View. In Biology of Stress in Fish; Fish Physiology Series; Schreck, C.B., Tort, L., Farrell, A.T., Brauner, C.J., Eds.; Academic Press-Elsevier: Cambridge, MA, USA, 2016; Volume 35, pp. 75–111. [Google Scholar]
- Flik, G.; Peter, H.M.; Klaren, P.H.M.; Erwin, H.; Van den Burg, J.R.; Huising, M.O. CRF and stress in fish. Gen. Comp. Endocrinol. 2006, 146, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Connell, L.A.; Fontenot, M.R.; Hofman, H.A. Characterization of the dopaminergic system in the brain of an african cichlid fish, Astatotilapia burtoni. J. Comp. Neurol. 2011, 519, 75–92. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Ruuskanen, J.O.; Wullimann, M.F.; Vernier, P. Two tyrosine hydroxylase genes in vertebrates: New dopaminergic territories revealed in the zebrafish brain. Mol. Cell. Neurosci. 2010, 43, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Moret, F.; Guilland, J.C.; Coudouel, S.; Rochette, L.; Vernier, P. Distribution of tyrosine hyfroxilase, dopamine, and serotonin in the central nervous system of amphioxus (Branchiostoma lanceolatum): Implications for the evolution of catecholamine systems in vertebrates. J. Comp. Neurol. 2004, 468, 135–150. [Google Scholar] [CrossRef]
- Lillesaar, C. The serotonergic system in fish. J. Chem. Neuroanat. 2011, 41, 294–308. [Google Scholar] [CrossRef]
- Pierre, J.; Repérant, J.; Ward, R.; Vesselkin, N.P.; Rio, J.-P.; Miceli, D.; Kratskin, I. The serotoninergic system of the brain of the lamprey, Lampetra fluviatilis: An evolutionary perspective. J. Chem. Neuroanat. 1992, 5, 195–219. [Google Scholar] [CrossRef]
- Øverli, Ø.; Winberg, S.; Pottinger, T.G. Behavioral and neuroendocrine correlates of selection for stress responsiveness in rainbow trout-a review. Integr. Comp. Biol. 2005, 45, 463–474. [Google Scholar] [CrossRef] [Green Version]
- Gesto, M.; Soengas, J.L.; Míguez, J.M. Acute and prolonged stress responses of brain monoaminergic activity and plasma cortisol levels in rainbow trout are modified by PAHs (naphthalene, β-naphthoflavone and benzo(a)pyrene) treatment. Aquat. Toxicol. 2008, 86, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Winberg, S.; Lepage, O. Elevation of brain 5-HT activity, POMC expression, and plasma cortisol in socially subordinate rainbow trout. Am. J. Physiol. Reg. Int. Comp. Physiol. 1998, 274, 645–R65. [Google Scholar] [CrossRef] [PubMed]
- Höglund, E.; Weltzien, F.A.; Schjolden, J.; Winberg, S.; Ursin, H.; Døving, K.B. Avoidance behavior and brain monoamines in fish. Brain Res. 2005, 1032, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Ruibal, C.; Soengas, J.L.; Aldegunde, M. Brain serotonin and the control of food intake in rainbow trout (Oncorhynchus mykiss): Effects of changes in plasma glucose levels. J. Comp. Physiol. A 2002, 188, 479–484. [Google Scholar]
- De Pedro, N.; Delgado, M.J.; Gancedo, B.; Alonso-Bedate, M. Changes in glucose, glycogen, thyroid activity and hypothalamic catecholamines in tench by starvation and refeeding. J. Comp. Physiol. 2003, 173, 475–481. [Google Scholar] [CrossRef] [PubMed]
- Øverli, Ø.; Harris, C.A.; Winberg, S. Short-term effects of fights for social dominance and the establishment of dominant-subordinate relationships on brain monoamines and cortisol in rainbow trout. Brain Behav. Evol. 1999, 54, 263–275. [Google Scholar] [CrossRef]
- Øverli, Ø.; Pottinger, T.G.; Carrick, T.R.; Øverli, E.; Winberg, S. Brain monoaminergic activity in rainbow trout selected for high and low stress responsiveness. Brain Behav. Evol. 2001, 57, 214–224. [Google Scholar] [CrossRef] [Green Version]
- López-Patiño, M.A.; Conde-Sieira, M.; Gesto, M.; Librán-Pérez, M.; Soengas, J.L.; Míguez, J.M. Melatonin partially minimizes the adverse stress effects in Senegalese sole (Solea senegalensis). Aquaculture 2013, 388, 165–172. [Google Scholar] [CrossRef]
- Conde-Sieira, M.; Muñoz, J.L.P.; López-Patiño, M.A.; Gesto, M.; Soengas, J.L.; Míguez, J.M. Oral administration of melatonin counteracts several of the effects of chronic stress in rainbow trout. Domest. Anim. Endocrinol. 2014, 46, 26–36. [Google Scholar] [CrossRef]
- Samaras, A.; Espírito Santo, C.; Papandroulakis, N.; Mitrizakis, N.; Pavlidis, M.; Höglund, E.; Pelgrim, T.N.M.; Zethof, J.; Spanings, F.A.T.; Vindas, M.A.; et al. Allostatic load and stress physiology in European seabass (Dicentrarchus labrax L.) and gilthead Seabream (Sparus aurata L.). Front. Endocrinol. 2018, 9, art 451. [Google Scholar] [CrossRef] [Green Version]
- Herrera, M.; Fernández-Alacid, L.; Sanahuja, I.; Ibarz, A.; Salamanca, N.; Morales, E.; Giráldez, I. Physiological and metabolic effects of a tryptophan-enriched diet to face up chronic stress in meagre (Argyrosomus regius). Aquaculture 2020, 522, 735102. [Google Scholar] [CrossRef]
- Pavlidis, M.; Mylonas, C. Sparidae: Biology and Aquaculture of Gilthead Sea Bream and Other Species; John Wiley and Sons: Southern Gate, UK, 2011. [Google Scholar]
- Mancera, J.M.; Vargas-Chacoff, L.; García-López, A.; Kleszczyńska, A.; Kalamarz, H.; Martínez-Rodríguez, G.; Kulczykowska, E. High density and food deprivation affect arginine vasotocin, isotocin and melatonin in gilthead sea bream (Sparus auratus). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2008, 149, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Skrzynska, A.K.; Martos-Sitcha, J.A.; Martínez-Rodríguez, G.; Mancera, J.M. Unraveling vasotocinergic, isotocinergic and stress pathways after food deprivation and high stocking density in the gilthead sea bream. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2018, 215, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Arends, R.J.; Mancera, J.M.; Muñoz, J.L.; Wendelaar Bonga, S.E.; Flik, G. The stress response of the gilthead sea bream (Sparus aurata L.) to air exposure and confinement. J. Endocrinol. 1999, 163, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Sangiao-Alvarellos, S.; Guzmán, J.M.; Láiz-Carrión, R.; Míguez, J.M.; Martín del Río, M.P.; Mancera, J.M.; Soengas, J.L. Interactive effects of high stocking density and food-deprivation on carbohydrate metabolism in several tissues of gilthead sea bream, Sparus aurata. J. Exp. Zool. 2005, 303, 761–775. [Google Scholar] [CrossRef]
- Martos-Sitcha, J.A.; Wunderink, Y.S.; Straatjes, J.; Skrzynska, A.K.; Mancera, J.M.; Martínez-Rodríguez, G. Different stressors induce differential responses of the CRH-stress system in the gilthead sea bream (Sparus aurata). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2014, 177, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Gesto, M.; López-Patiño, M.A.; Hernández, J.; Soengas, J.L.; Míguez, J.M. The response of brain serotonergic and dopaminergic systems to an acute stressor in rainbow trout: A time course study. J. Exp. Biol. 2013, 216, 4435–4442. [Google Scholar] [CrossRef] [Green Version]
- Skrzynska, A.K.; Maiorano, E.; Bastaroli, M.; Naderi, F.; Míguez, J.M.; Martínez-Rodríguez, G.; Mancera, J.M.; Martos-Sitcha, J.A. Impact of air exposure on vasotocinergic and isotocinergic systems in gilthead sea bream (Sparus aurata): New insights on fish stress response. Front. Physiol. 2018, 9, 96. [Google Scholar] [CrossRef] [Green Version]
- Gesto, M.; Tintos, A.; Soengas, J.L.; Míguez, J.M. Effects of acute and prolonged naphthalene exposure on brain monoaminergic neurotransmitters in rainbow trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. 2006, 144C, 173–183. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef]
- Seginer, I. Growth models of gilthead sea bream (Sparus aurata L.) for aquaculture: A review. Aquacult. Eng. 2016, 70, 15–32. [Google Scholar] [CrossRef]
- Gesto, M.; Otero-Rodiño, C.; López-Patiño, M.A.; Míguez, J.M.; Soengas, J.L.; Conde-Sieira, M. Is plasma cortisol response to stress in rainbow trout regulated by catecholamine-induced hyperglycemia? Gen. Comp. Endocrinol. 2014, 205, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Winberg, S.; Nilsson, A.; Hylland, P.; Söderstöm, V.; Nilsson, G.E. Serotonin as a regulator of hypothalamic-pituitary-interrenal activity in teleost fish. Neurosci. Lett. 1997, 230, 113–116. [Google Scholar] [CrossRef]
- Reid, S.G.; Bernier, N.J.; Perry, S.F. The adrenergic stress response in fish: Control of catecholamine storage and release. Comp. Biochem. Physiol. 1998, 120C, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Fabbri, E.; Moon, T.W. Adrenergic signaling in teleost fish liver, a challenging path. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2016, 199, 74–86. [Google Scholar] [CrossRef]
- Atwood, H.L.; Tomasso, J.R.; Ronan, P.J.; Barton, B.A.; Renner, K.J. Brain monoamine concentrations as predictors of growth inhibition in channel catfish exposed to ammonia. J. Aquat. Anim. Health 2000, 12, 69–73. [Google Scholar] [CrossRef]
- Ortega, V.A.; Renner, K.J.; Bernier, N.J. Appetite-suppressing effects of ammonia exposure in rainbow trout associated with regional and temporal activation of brain monoaminergic and CRF systems. J. Exp. Biol. 2005, 208, 1855–1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldegunde, M.; Mancebo, M. Effect of neuropeptide Y on food intake and brain biogenic amines in the rainbow trout (Oncorhynchus mykiss). Peptides 2006, 27, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Soengas, J.L.; Aldegunde, M. Brain glucose and insulin: Effects on food intake and brain biogenic amines of rainbow trout. J. Comp. Physiol. A 2004, 190, 641–649. [Google Scholar] [CrossRef]
- Lepage, O.; Tottmar, O.; Winberg, S. Elevated dietary intake of l-tryptophan counteracts the stress-induced elevation of plasma cortisol in rainbow trout (Oncorhynchus mykiss). J. Exp. Biol. 2002, 205, 3679–3687. [Google Scholar] [CrossRef]
- Höglund, E.; Balm, P.H.M.; Winberg, S. Skin darkening, a potential social signal insubordinate Arctic charr (Salvelinus alpinus): The regulatory role of brain monoaminesand pro-opiomelanocortin-derived peptides. J. Exp. Biol. 2000, 203, 1711–1721. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Parameter | Density | Fed | Food-Deprived |
|---|---|---|---|
| Cortisol | Low | 4.17 ± 0.67 | 10.06 ± 2.68 * |
| High | 27.36 ± 6.82 # | 23.87 ± 6.61 # | |
| Adrenaline | Low | 97.56 ± 10.05 | 80.85 ± 13.97 |
| High | 148.74 ± 27.53 | 108.63 ± 35.57 | |
| Noradrenaline | Low | 79.78 ± 6.72 | 49.03 ± 4.99 * |
| High | 91.88 ± 13.67 | 70.92 ± 24.62 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Patiño, M.A.; Skrzynska, A.K.; Naderi, F.; Mancera, J.M.; Míguez, J.M.; Martos-Sitcha, J.A. High Stocking Density and Food Deprivation Increase Brain Monoaminergic Activity in Gilthead Sea Bream (Sparus aurata). Animals 2021, 11, 1503. https://doi.org/10.3390/ani11061503
López-Patiño MA, Skrzynska AK, Naderi F, Mancera JM, Míguez JM, Martos-Sitcha JA. High Stocking Density and Food Deprivation Increase Brain Monoaminergic Activity in Gilthead Sea Bream (Sparus aurata). Animals. 2021; 11(6):1503. https://doi.org/10.3390/ani11061503
Chicago/Turabian StyleLópez-Patiño, Marcos Antonio, Arleta Krystyna Skrzynska, Fatemeh Naderi, Juan Miguel Mancera, Jesús Manuel Míguez, and Juan Antonio Martos-Sitcha. 2021. "High Stocking Density and Food Deprivation Increase Brain Monoaminergic Activity in Gilthead Sea Bream (Sparus aurata)" Animals 11, no. 6: 1503. https://doi.org/10.3390/ani11061503