1. Introduction
Fipronil (FPN) is an N-phenyl-pyrazole insecticide with a wide range and can be used in insect control in various cereal crops in agriculture, veterinary activities, and public health management [
1]. FPN is an insect neurotoxin agent, and the critical mechanism is active when the GABA-regulated chloride channels are blocked, causing depression and death in the central nervous system [
2]. FPN insecticides are the only organic toxicants to be applied to the ecosystem in a targeted manner to improve food safety by battling pests and regulating disease vectors [
3]. Insecticide toxicity is well known to have multiple consequences; creating oxidative injury is a high concept due to reactive oxygen species [
4]. Extensive use of these insecticides in agriculture and residential settings causes chronic neurological syndromes, teratogenicity, male reproductive failure, fetal growth retardation, embryo fetotoxicity, and genotoxicity [
5]. Beyond liver toxicity, FPN also had reproductive effects, as studies found that the application of FPN influences fertility [
6]. The excess output of reactive oxygen species leads to oxidative stress that decreases sperm fertility [
7]. The level of serum hepatic enzymes and renal function biomarkers (creatinine and urea), cholesterol, and lactate dehydrogenase increased markedly in FPN intoxication; however, overall serum protein, albumin, and triglycerides decreased significantly, as well as a major increase in malondialdehyde and nitric oxide levels with a significant reduction in glutathione (GSH), glutathione peroxidase (GPx), superoxide dismutase (SOD), and catalase (CAT) [
8]. FPN induced deterioration within the seminiferous tubules and apoptosis in the epididymides. Upregulated
interleukin-1β,
nitric oxide synthase 2,
caspase-3 (
Casp3) as well as downregulated
Burkitt-cell lymphomas, inhibin
B proteins, and
androgen receptor mRNA expressions
Casp3,
nitric oxide synthase,
ionized calcium-binding adapter molecule 1, and
IL-1β immunoreactions were increased. There was also a reduction of
proliferating cell nuclear antigen (
PCNA),
mouse vasa homolog (
MVH), and
SOX9 protein reactions [
9].
Many studies report the effect of fipronil on the reproductive system on different animals in which Ohi et al. [
10] reported that when fipronil was topically utilized to rats (single dose) at different concentrations (70, 140, and 280 mg/kg), it altered the cyclicity of female rats and had harmful reproductive effects in female rats. Mazzo et al. [
6] reported that male rats that received fipronil 5 mg/kg for 14 days had decreased sperm production, reduced epidydimal sperm count, a reduction in GSH, and an increase in the concentration of malondialdehyde. In addition, Eisa et al. [
5] reported that rats treated with different doses of fipronil 1/10 LD50 (2.1 mg/kg bwt) and 1/30 LD50 (0.7 mg/kg bwt) at the 6th to 15th days of pregnancy lead to teratogenic and embryotoxic effects. De Barros et al. [
11] reported that pregnant rats exposed (via gavage) to fipronil (0.03, 0.3, or 3 mg/kg) from gestational day 15 until postnatal day 7 had infertility. In addition, Kitulagodage et al. [
12] breeding female zebra finches orally dosed with single sublethal levels of fipronil (1, 5, and 10 mg/kg body weight) had a decrease in hatchability percentage.
Medicinal herbs are used extensively because of their antimicrobial, antioxidant, and less toxic effects than chemical substances. Boswellic acid (BA) is a pentacyclic triterpene, which is a compound isolated from Boswellia serrata gum resin. It is potent against several inflammatory diseases, including cancer, arthritis, ulcerative colitis, respiratory inflammatory disorders, brain tumor, fertility, and memory [
13]. This gum-resin is applied in conventional Chinese medicine to remedy many aspects of well-being [
14]. Many in vitro trials indicated that BA hinders the synthesis of the pro-inflammatory enzyme such as
5-lipoxygenase (
5-LO),
cyclooxygenase-1 (
COX-1),
human elastase of the leucocytes (
HLE),
cathepsin G (
cat G), and
microsomal prostaglandin E (
mPGES-1), together with
nuclear factor kappa B (
NF-μB) inhibition and various cytokines such as
TNFα,
IL-1β, and
IL-6, respectively [
15]. Many clinical reports have also illustrated BA’s possible value as an anti-inflammatory agent [
16,
17,
18] and antioxidant activity [
19]. Therefore, this study was designed to determine BA’s potential protective impact against oxidative and testicular damage caused by FPN insecticide poisoning on the male rat model.
4. Discussion
Chemical insecticides are widely utilized worldwide in the agriculture sector and for other purposes [
38]. Food residues, contaminated tap water, occupational exposure, repellence, household use, and application against fleas and ticks are various sources for endangering insecticides for animals and people [
39,
40]. FPN is an insecticide with phenylpyrazole in chemical form. It is a common insecticide used both in agriculture and in domestic pest management [
25]. However, few studies are evaluating its consequence on the fertility of males and reproductive efficiency. Therefore, this experiment was conducted to determine the effects of FPN on fertility test, the weight of the male reproductive organ, seminal study, serum testosterone level, oxidative status, cytokines level, some gene expression, and histopathology and to evaluate the ameliorative effect of BA. In our study, FPN has a distinct adverse effect on fertility tests by decreasing pregnant females, pregnancy index, and many litters.
Moreover, it gave rise to a noteworthy decrease in the reproductive organs’ weights due to the decline in serum testosterone concentration, sperm quantity, sperm progressive motility, and live sperm cell percentage. It considerably boosted sperm cell abnormalities percentage in the FPN-administered group, which many attribute to its hazard impact. Consequently, the findings obtained indicate that FPN decreases sperm cell quality, leading to male rats’ infertility. The pathway of insecticide toxicity on the testicular tissue can be correlated with the activation of oxidative injuries. These results agree with Mazzo, Balieira, Bizerra, and Mingatto [
6], who reported that FPN-induced harmful impacts on sperm quality. Sperm motility is impaired by FPN’s long-term exposure [
11]. In addition, the overproduction of reactive oxygen species (ROS) exceeds the cellular capability, leads to oxidative damage, and reduces sperm viability and fertility [
1]. The sperm contains a large proportion of polyunsaturated fatty acids and is highly susceptible to harm caused by excessive oxidative damage and peroxidation to its plasma membrane, leading to a loss of motility and decreased number [
41,
42]. FPN-administered rats showed a remarkable increase in MDA, which considers the consequence of lipid peroxidation and lipid degradation triggering radicals and reduction of GSH levels, which reverberate the degree of oxidative harm. These results are following the work of Mossa et al. [
43] that demonstrated decreased concentrations of GSH in the kidneys and liver of rats handled with FPN (10 mg/kg bwt) [
44]. The reduction in the content of GSH in the kidney and brain of FPN-treated mice (5 and 10 mg/kg bwt) in the same FPN-treated line (5 mg/kg bwt) resulted in a decrease in the concentration of GSH in the testis [
6]. These results are probably attributed to Fipronil’s oxidative anxiety, including reduced GSH levels, and antioxidant activity and consequent lipoperoxidation [
45]. FPN-administered rats showed an overexpression of TNF-α and IL-6 pro-inflammatory cytokines. TNF-α is a major inflammatory and immune response cytokine [
46], and IL-6 is a cytokine pleiotropic rendered by macrophages of tissue and monocytes [
47].
The utilized doses of boswellic acid in the current study were tested previously as reported by Sami et al. [
21] in which they found that boswellic acids can ameliorate doxorubicin-induced nephrotoxicity in mice, they used different doses of boswellic acid (125 mg/kg), (250 mg/kg), and (500 mg/kg), and they found that the effect of the high dose of BAs (500 mg/kg) was different (more ameliorative) from that observed with the lowest dose (125 mg/kg). In the same line, Nusier et al. [
20] studied the effect of two different doses of boswellic acid: 250 and 500 mg/kg. They reported variation in their impact on the reproductive system of the rat. Barakat et al. [
22,
23] investigated the protective effect of boswellic acid in different doses 250 and 500 in doxorubicin-induced hepatic damaged. The anti-aggregatory effect of boswellic acid in high-fat fed rats found a variation in the boswellic acid in a dose-dependant manner. Al-Yahya et al. [
24] reported that the boswellic acid is safe up to 1000 mg/kg in rats. Still, this dose is relatively high considering the amount of extract consumed by humans. So, in this study, we try to investigate the ameliorative effect of boswellic acid on the toxic impact of fipronil in a dose–response manner.
The impact of FBN and BA on fertility tests is presented in
Table 2. Almost all females recorded positive sperm in all other groups other than the FBN group; moreover, the pregnancy index (%) was highest in the control and BA groups. Still, BA decreases this adverse effect, while the control group and BA groups (250 mg and 500 mg) were the highest in the number of litters. The obtained results were inconsistent with [
20]. They reported that oral administration of boswellic acid increased the fertility in rats and the number of implantations as well as increased spermatogenesis due to its antioxidant activity. In the same line, our work was in harmony with [
20], in which they reported non-significant improvement in the fertility parameters in BA (500 mg/kg) dose concerning (250 mg/kg). The concomitant administration of both doses of BA with FPN significantly increases testosterone and GSH with a significant decrease in MDA, interleukin-6, and TNF-α concentrations concerning the FBN-treated group, as shown in
Table 5. with non-significant improvement in the BA higher dose concerning the lower dose of BA. The obtained result was in the same line with Sami et al. [
21] in which they reported the nephroprotective effect of BA with a non-significant improvement of the higher dose of BA (500 mg/kg) due to the antioxidant and antiapoptotic effect of boswellic acid.
The results showed that FPN-administered rats showed a significant downregulation in steroidogenic and fertility-related gene expression, including
CYP17A1 and
cytochrome P450 17A1,
KISS1,
kisspeptin,
STAR,
Cyp11a1, cholesterol side-chain cleavage enzyme mRNA (
P450SCC)
Hsd3b1, 3-beta-hydroxysteroid dehydrogenase/delta-5-delta-4 isomerase type I,
Cyp19, and cytochrome P450 aromatase compared with other administered groups and the control one. However, co-administration of BA (both doses) with FPN showed significant upregulation and stabilization of the steroidogenic gene expression. Steroid hormones are produced from cholesterol through various steroid cytochrome P450 hydroxylases-induced reactions [
48,
49]. Cholesterol transfer from the external to the mitochondria’s inner membrane by steroidogenic acute regulating protein (StAR) involves a rate-restricted steroidogenesis phase [
50]. Then, steroidogenesis begins with the transition of P450 (P450scc/CYP11A1/Cyp11a1) cholesterol to pregnenolone, which is an essential molecule in developing the body steroid hormones [
48].
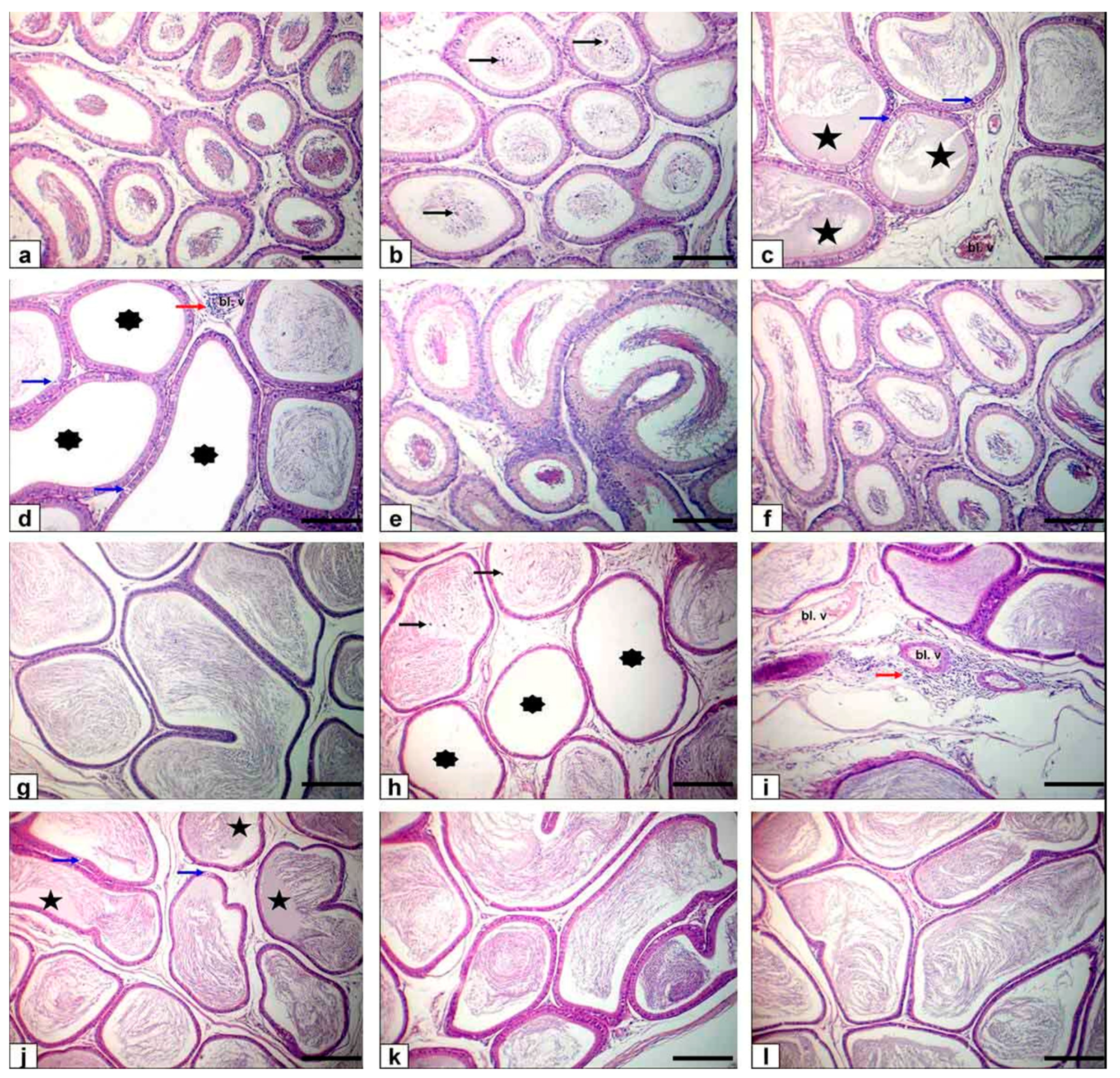
Light microscope examination of FPN-administered rats testes showed degenerated and exfoliated germ cells in the seminiferous tubular lumen, which was attributed by the main effect of the cell-to-cell junction of Sertoli and germ cells or microtubular deterioration that lead to Sertoli cell damage [
51,
52]. In the current work, fragmentation or necrosis of tubular epithelium with giant cell formations in the seminiferous tubular lumen, germ cell loss, and luminous hyalinization dwindled, collapsed, and incoherent seminiferous tubules were reported. FPN caused interstitial edema because of increased vascular permeability and mononuclear cell infiltration that secrete cytokines, causing congested interstitial blood vessels. Another important finding in atrophied seminiferous tubules was noticed in our study due to the cytotoxic effect of FPN. The epididymal lesions were sloughing off some germinal epithelial in the lumen of some ducts of the epididymis, which indicate testicular malfunction [
53,
54] and interstitial congestion of blood vessel with perivascular inflammatory cell infiltrations; besides, most epididymal ducts seemed to have no or low sperm quantities in their lumen, which reflected the cessation of spermatogenesis. The prostates and seminal vesicle lesions experienced necrosis and desquamation of some glandular epithelial cells with low luminal discharges and severe interstitial leukocytes infiltration [
53]. These alterations may be due to reduced testosterone, which needs differentiation, development, and the preservation of epithelial cells of accessory sex glands [
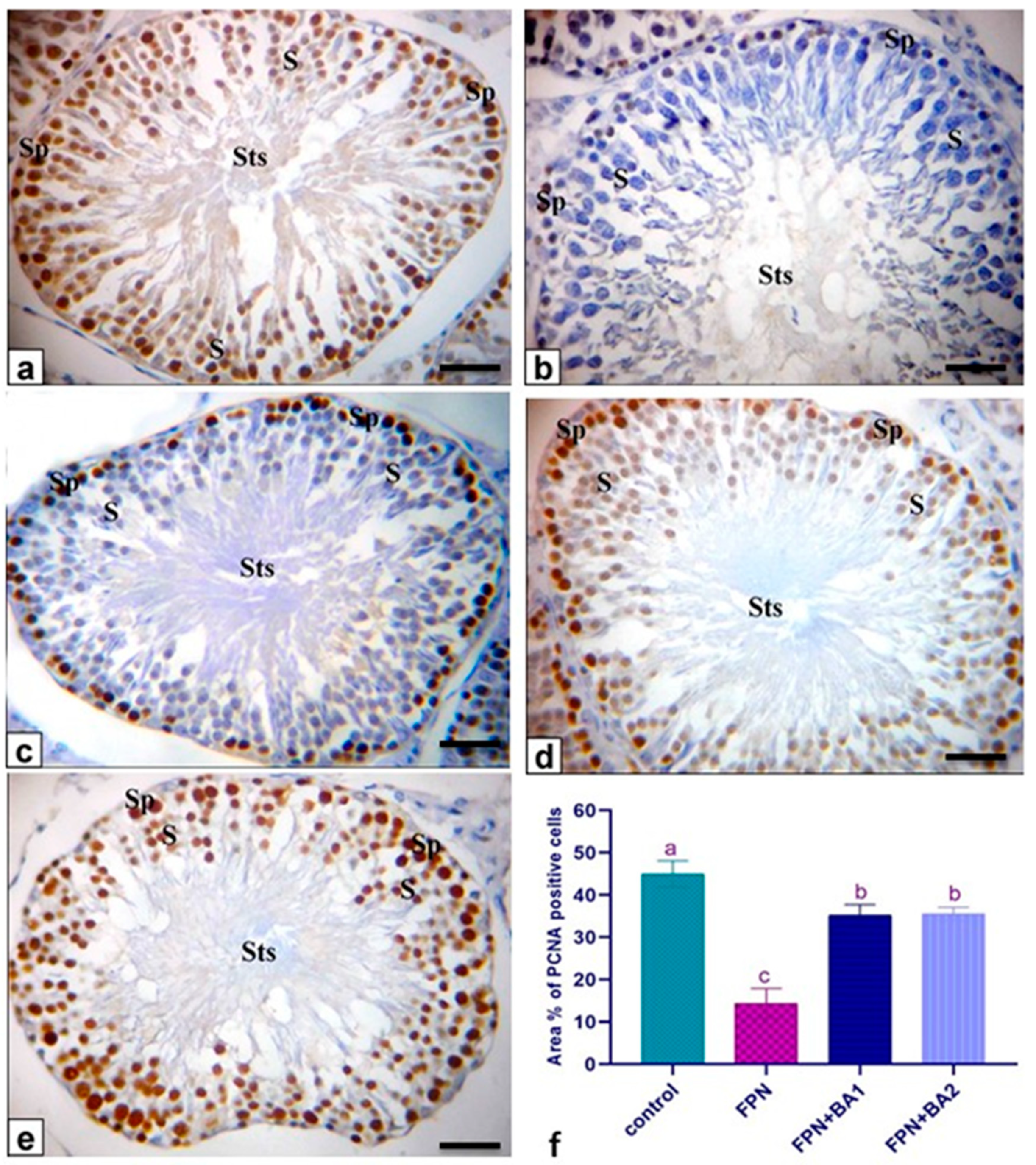
55]. To estimate germ cell kinetics and an indication of DNA synthesis deterioration, PCNA is considered a valuable molecular marker [
56]. Similarly, it measures the cell proliferation and spermatogenic role of studies in male infertility. In spermatogonia and the proliferating control rat spermatocytes, many positive brown nuclear reactions have been found, while a reduced countenance and area percentage of PCNA antibody was detected in FPN-administered rats compared with the control group. There were no histological architecture differences among the control and two levels of BA (250–500 mg/kg) groups in the examined tissues. Conversely, rats that received FPN plus BA (250 and 500 mg/kg) showed significant enhancement of most seminiferous tubules’ spermatogenesis by including elongated spermatids and spermatozoa. These results were inconsistent with [
21] in which they reported that the most convenient restoration of these parameters was achieved in the doxorubicin + BA 500 mg/kg group. Supporting our result, data obtained by Kruger et al. [
57] have reported antioxidants and chelate metals of BAs in oxidative injury pathways.
The co-administration of BA significantly improved the evaluated parameters such as the number of pregnant rats, pregnancy index, and the number of litters, reproductive organs weights, normalized testosterone levels, and sperm quality, which were attributed to the increasing number of spermatocytes and normal spermatogenesis as found by Nusier et al. [
20], which reported the promising effect of boswellia on adult male rat fertility due to its antioxidant activity [
19]. BA treatment (250 and 500 mg/kg) normalizes the apoptotic effect of FPN, which is cleared by our result, in which FPN significantly upregulates Bax, heat shock protein, and gene expression with significant downregulation to Bcl-2. These results show inconsistency with [
21]; they revealed the antioxidant and antiapoptotic effects of BA. BA’s prospective role in the scavenging of FPN ROS was linked in histological architecture to testes, the epididymis, and sexual accessory glands. Upon our result, the molecular mechanism of FBN that induced infertility was summarized in the downregulation in steroidogenic and fertility-related gene expression, including
CYP17A1 and
cytochrome P450 17A1,
KISS1,
kisspeptin,
STAR,
Cyp11a1, cholesterol side-chain cleavage enzyme mRNA (
P450SCC)
Hsd3b1, 3-beta-hydroxysteroid dehydrogenase/delta-5-delta-4 isomerase type I,
Cyp19, and cytochrome P450 aromatase. In addition, FPN-treated rats showing few PCNA immunoreactions in the nuclei of the spermatogenic cells and negative spermatocytes FPN caused interstitial edema because of increased vascular permeability and mononuclear cell infiltration that secrete cytokines
IL-6 and
TNF-α, causing congested interstitial blood vessels. Another important finding regarding atrophied seminiferous tubules was noticed in our study due to the cytotoxic effect of FPN blood serum testosterone, and testicular levels of GSH in FBN-treated group were appreciably (
p ≤ 0.05) decreased.
Many studies are concerned with the significant mechanism driving the protective actions in which boswellic acid-treated rats showed reduced serum expression of
TNF-α and
IL-6 and hepatic iNOS. At the cellular level, also, boswellic acids increased the expression in the white adipose tissue of thermogenesis associated mitochondrial uncoupling protein-1 and carnitine palmitoyl transferase-1 [
58].
Gayathri et al. [
59] showed that boswellic acid has anti-inflammatory effects in human peripheral mononuclear blood cells and mouse macrophages by inhibiting
tumor necrosis factor-α (
TNF-α),
IL-1β,
nitric oxide, and mitogenic protein kinases. Several clinical trials highlighted boswellic acid as a potentially effective anti-inflammatory drug [
60]. In addition, BA extract inhibited the
TGF-ß-induced fibrosis (
p = 0.01) and 5-lipooxygenase activity levels that prevent fibrosis, as mentioned by Ali and Mansour [
61]. In the same line, Sharma, et al. [
62] reported the anti-inflammatory role of the boswellic acid through the inflammatory mediators
TNF-α and
IL-6. The inhibition of
NF-κB activity by the boswellic acid family may be considered an alternative therapy for chronic inflammatory disorders [
63].


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}