
Histological and Immunohistochemical Characterization of Vomeronasal Organ Aging in Mice


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sampling Procedure
2.2. Histochemical Analyses
2.3. Immunohistochemical Analyses
2.4. Statistical Analyses
3. Results
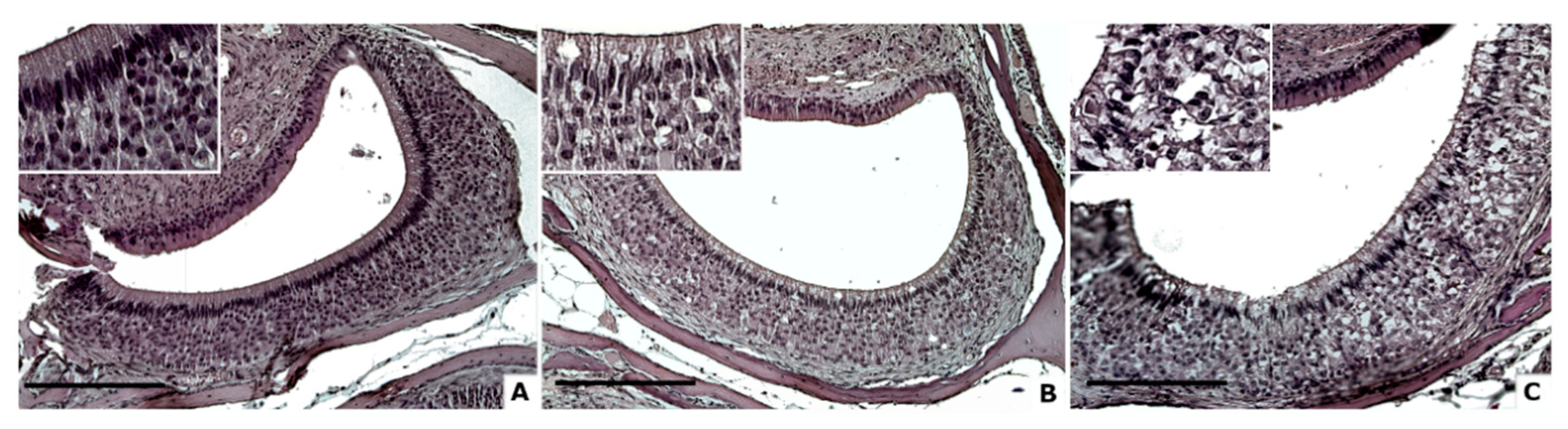
3.1. Cellular Degeneration
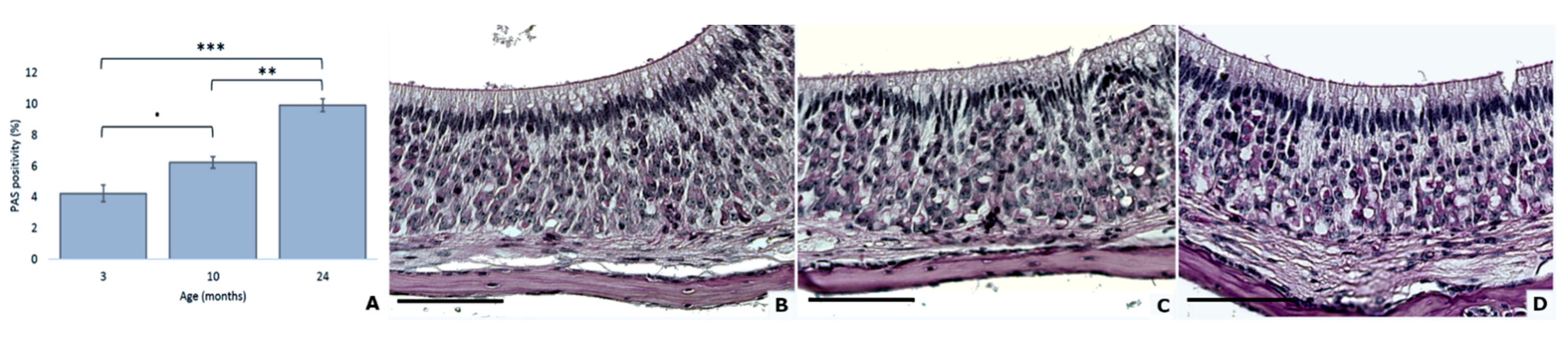
3.2. Increased Quantity of Accumulated Glycogen with Age
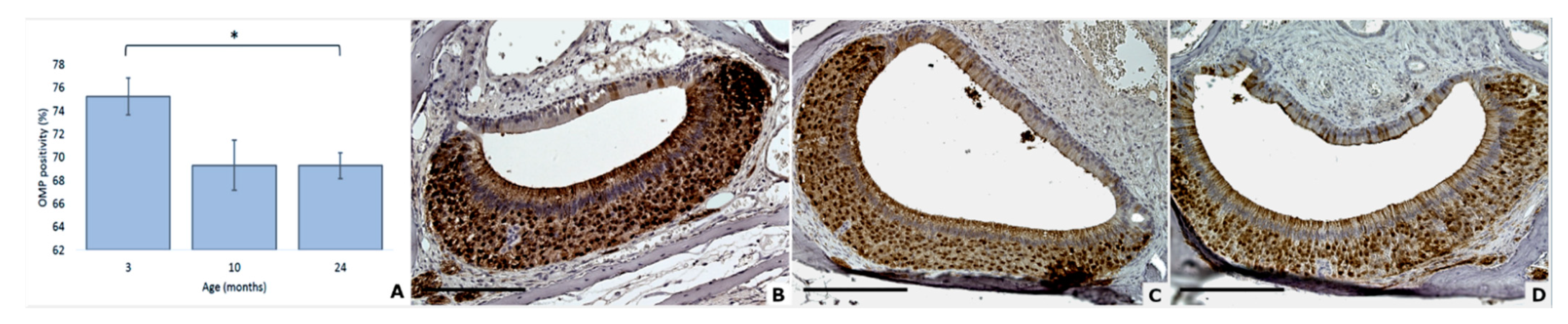
3.3. OMP Expression Decreases with Age
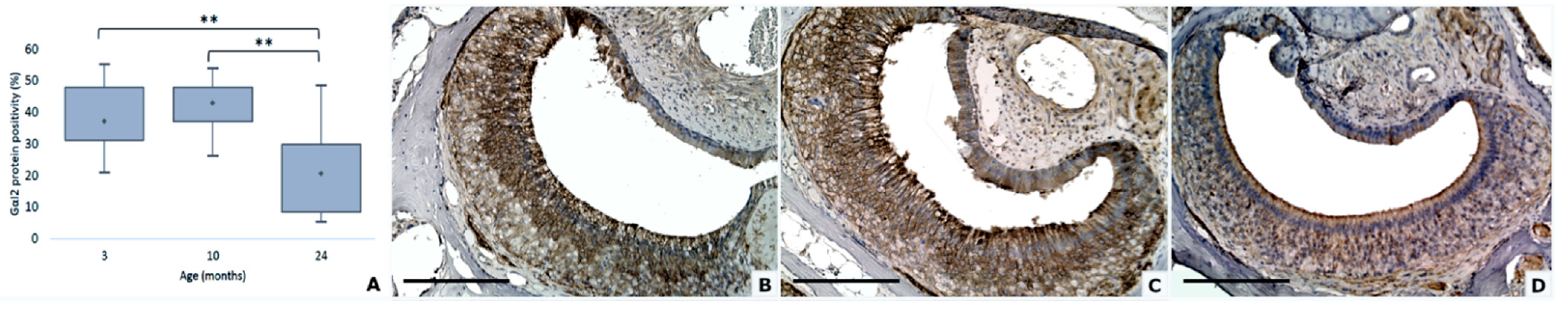
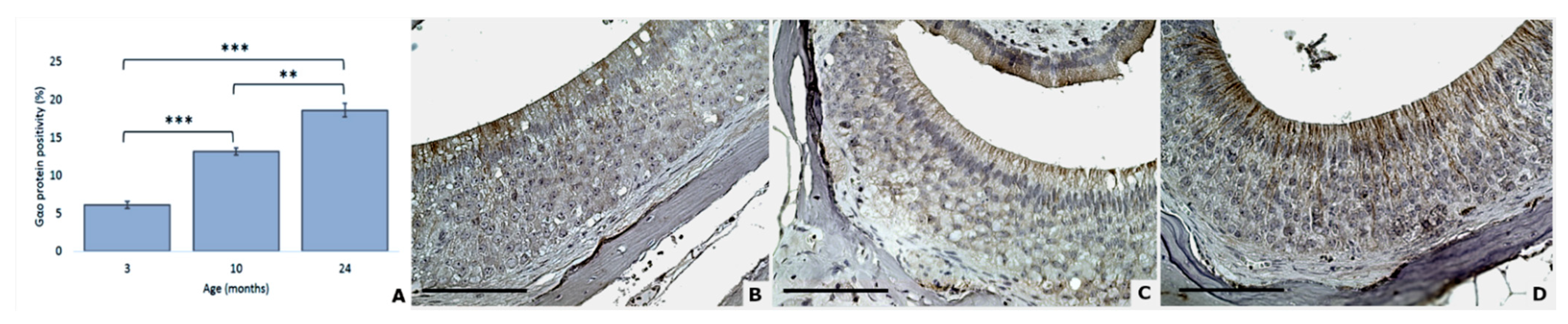
3.4. Gαi2 and Gαo Proteins Modifications
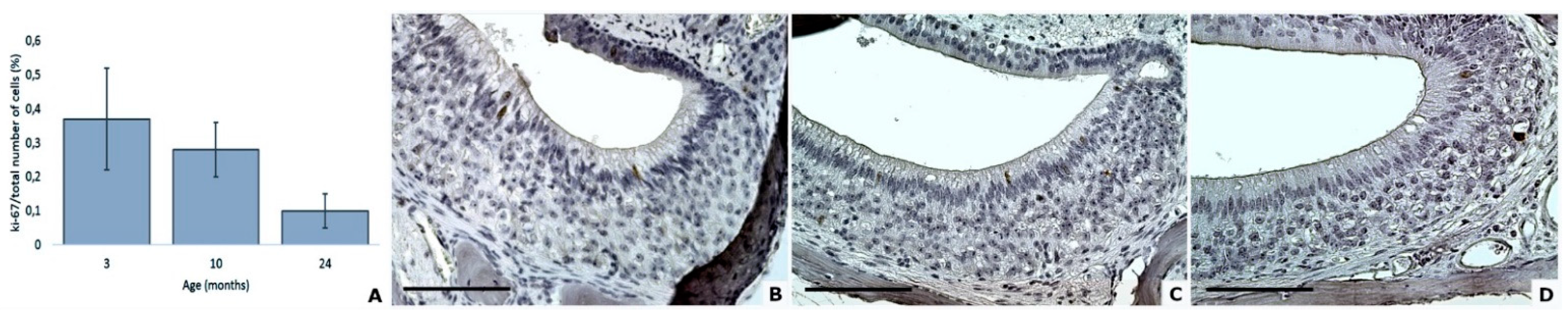
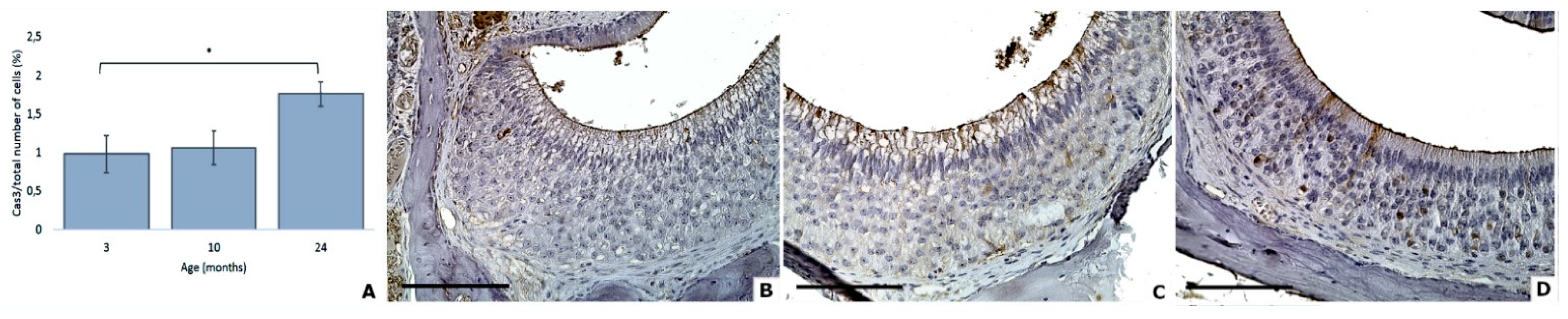
3.5. Proliferation and Apoptosis Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Adams, D.R.; Wiekamp, M.D. The canine vomeronasal organ. J. Anat. 1984, 138, 771–787. [Google Scholar] [PubMed]
- Brossut, R. Phéromones: La Communication Chimique Chez les Animaux; CNRS Éditions: Paris, France, 1996; p. 143. [Google Scholar]
- Salazar, I.; Quinteiro, P.S.; Cifuentes, J.M. The Soft-tissue Components of the Vomeronasal Organ in Pigs, Cows and Horses. Anat. Histol. Embryol. 1997, 26, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Baxi, K.N.; Dorries, K.M.; Eisthen, H.L. Is the vomeronasal system really specialized for detecting pheromones? Trends Neurosci. 2006, 29, 1–7. [Google Scholar] [CrossRef]
- Rivière, S.; Challet, L.; Fluegge, D.; Spehr, M.; Rodriguez, I. Formyl peptide receptor-like proteins are a novel family of vomeronasal chemosensors. Nature 2009, 459, 574–577. [Google Scholar] [CrossRef]
- Isogai, Y.; Si, S.; Pont-Lezica, L.; Tan, T.; Kapoor, V.; Murthy, V.N.; Dulac, C. Molecular Organization of Vomeronasal Chemoreception. Nature 2012, 478, 241–245. [Google Scholar] [CrossRef] [Green Version]
- Zufall, F.; Kelliher, K.R.; Leinders-Zufall, T. Pheromone detection by mammalian vomeronasal neurons. Microsc. Res. Tech. 2002, 58, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Halpern, M.; Martinez-Marcos, A. Structure and function of the vomeronasal system: An update. Prog. Neurobiol. 2003, 70, 245–318. [Google Scholar] [CrossRef]
- Rodriguez, I. Vomeronasal Receptors: V1Rs, V2Rs, and FPRs. In Chemosensory Transduction: The Detection of Odors, Tastes, and Other Chemostimuli, 1st ed.; Zufall, F., Munger, S.D., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 10, pp. 175–190. [Google Scholar]
- Dulac, C.; Axel, R. A novel family of genes encoding putative pheromone receptors in mammals. Cell 1995, 83, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Herrada, G.; Dulac, C. A novel family of putative pheromone receptors in mammals with a topographically organized and sexually dimorphic distribution. Cell 1997, 90, 763–773. [Google Scholar] [CrossRef] [Green Version]
- Pearlman, S.J. Jacobson’s organ: Its anatomy, gross, microscopic and comparative, with some observations as well on its function. Ann. Otol. Rhinol. Laryngol. 1934, 43, 739–768. [Google Scholar] [CrossRef]
- Salazar, I.; Quinteiro, P.S. The risk of extrapolation in neuroanatomy: The case of the mammalian vomeronasal system. Front. Neuroanat. 2009, 3, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyatt, T.D. Pheromones and Animal Behavior: Chemical Signals and Signature Mixes, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Wysocky, C.; Lepri, J. Consequences of removing the vomeronasal organ. J. Steroid Biochem. Mol. Biol. 1991, 39, 661–669. [Google Scholar] [CrossRef]
- Booth, K.K.; Katz, L.S. Role of the vomeronasal organ in neonatal offspring recognition in sheep. Biol. Reprod. 2000, 63, 953–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pankevich, D.; Cherry, J.; Baum, M. Effect of vomeronasal organ removal from male mice on their preference for and neural Fos responses to female urinary odors. Behav. Neurosci. 2006, 120, 925–936. [Google Scholar] [CrossRef] [Green Version]
- Kiyokawa, Y.; Kikusui, T.; Takeuchi, Y.; Mori, Y. Removal of the vomeronasal organ blocks the stress-induced hyperthermia response to alarm pheromone in male rats. Chem. Senses 2007, 32, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booth, K.K.; Webb, E.C. Effect of blockage of the ducts of the vomeronasal organ on LH plasma levels during the “Whitten Effect” in does. Vet. Med. Int. 2010, 2011, 305468. [Google Scholar] [CrossRef] [Green Version]
- Asproni, P.; Cozzi, A.; Verin, R.; Lafont-Lecuelle, C.; Bienboire-Frosini, C.; Poli, A.; Pageat, P. Pathology and behaviour in feline medicine: Investigating the link between vomeronasalitis and aggression. J. Feline Med. Surg. 2016, 18, 997–1002. [Google Scholar] [CrossRef]
- Lawless, C.; Wang, C.; Jurk, D.; Merz, A.; Zglinicki, T.; Passos, J. Quantitative assessment of markers for cell senescence. Exp. Gerontol. 2010, 45, 772–778. [Google Scholar] [CrossRef]
- Martínez-Cué, C.; Rueda, N. Cellular Senescence in Neurodegenerative Diseases. Front. Cell Neurosci. 2020, 14, 16. [Google Scholar] [CrossRef]
- Wilson, K.C.; Raisman, G. Age-related changes in the neurosensory epithelium of the mouse vomeronasal organ: Extended period of postnatal growth size and evidence for rapid cell turnover in the adult. Brain Res. 1980, 185, 103–113. [Google Scholar] [CrossRef]
- Brann, J.; Firestein, S. A lifetime of neurogenesis in the olfactory system. Front. Neurosci. 2014, 8, 182. [Google Scholar] [CrossRef]
- Brann, J.; Firestein, S. Regeneration of New Neurons Is Preserved in Aged Vomeronasal Epithelia. J. Neurosci. 2010, 30, 15686–15694. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.C.; He, J.; Ma, M. Olfactory marker protein is critical for functional maturation of olfactory sensory neurons and development of mother preference. J. Neurosci. 2011, 31, 2974–2982. [Google Scholar] [CrossRef] [Green Version]
- Witt, M.; Thiemer, R.; Meyer, A.; Schmitt, O.; Wree, A. Main Olfactory and Vomeronasal Epithelium Are Differently Affected in Niemann-Pick Disease Type C1. Int. J. Mol. Sci. 2018, 19, 3563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flurkey, K.; Currer, J.M.; Harrisonet, D.E. Mouse Models in Aging Research. In The Mouse in Biomedical Research, 2nd ed.; Fox, J.G., Barthold, S.W., Davisson, M.T., Newcomer, C.E., Quimby, F.W., Smith, A.L., Eds.; Academic Press: Cambridge, MA, USA, 2007; pp. 639–668. [Google Scholar]
- Klatt, E.; Kumar, V. Robbins and Cotran. Review of Pathology, 4th ed.; Saunders: Philadelphia, PA, USA, 2015. [Google Scholar]
- Brown, A.M.; Sickmann, H.M.; Fosgerau, K.; Lund, T.M.; Schousboe, A.; Waagepetersen, H.S.; Ransom, B.R. Astrocyte glycogen metabolism is required for neural activity during a glycemia or intense stimulation in mouse white matter. J. Neurosci. 2005, 79, 74–80. [Google Scholar]
- Suzuki, A.; Stern, S.A.; Bozdagi Huntley, G.W.; Walker, R.H.; Magistretti, P.J.; Alberini, C.M. Astrocyte-neuron lac-tate transport is required for long-term memory formation. Cell 2011, 144, 810–823. [Google Scholar] [CrossRef] [Green Version]
- Duran, J.; Guinovart, J.J. Brain glycogen in health and disease. Mol. Asp. Med. 2015, 46, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Augé, E.; Cabezon, I.; Pelegri, C.; Vilaplana, J. New perspectives on corpora amylacea in the human brain. Nature 2017, 7, 41807. [Google Scholar] [CrossRef] [Green Version]
- Cavanagh, J.B. Corpora-amylacea and the family of polyglucosan diseases. Brain Res. Rev. 1999, 29, 265–295. [Google Scholar] [CrossRef]
- Manich, G.; Cabezib, I.; Augé, E.; Pelegri, C.; Vilaplana, J. Periodic acid-Schiff granules in the brain of aged mice: From amyloid aggregates to degenerative structures containing neo-epitopes. Ageing Res. Rev. 2016, 27, 42–55. [Google Scholar] [CrossRef]
- Dibattista, M.; Reisert, J. The Odorant Receptor-Dependent Role of Olfactory Marker Protein in Olfactory Receptor Neurons. J. Neurosci. 2016, 36, 2995–3006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyatt, T.D. Pheromones and signature mixtures: Defining species-wide signals and variable cues for identity in both invertebrates and vertebrates. J. Comp. Physiol. A 2010, 196, 685–700. [Google Scholar] [CrossRef] [PubMed]
- Trouillet, A.C.; Keller, M.; Weiss, J.; Leinders-Zufall, T.; Birnbaumer, L.; Zufall, F.; Chamero, P. Central role of G protein Gαi2 and Gαi2+ vomeronasal neurons in balancing territorial and infant-directed aggression of male mice. Proc. Natl. Acad. Sci. USA 2019, 116, 5135–5143. [Google Scholar] [CrossRef] [Green Version]
- Norlin, E.M.; Gussing, F.; Berghard, A. Vomeronasal Phenotype and Behavioral Alterations in Gαi2 Mutant Mice. Curr. Biol. 2003, 13, 1214–1219. [Google Scholar] [CrossRef] [Green Version]
- Chamero, P.; Katsoulidou, V.; Hendrix, P.; Bufe, B.; Roberts, R.; Matsunami, H.; Abramowitz, J.; Birnbaumer, L.; Zufall, F.; Leinders-Zufall, T. G protein G(alpha)o is essential for vomeronasal function and aggressive behavior in mice. Proc. Natl. Acad. Sci. USA 2011, 108, 12898–12903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wettschureck, N.; Offermanns, S. Mammalian G proteins and their cell type specific functions. Physiol. Rev. 2005, 85, 1159–1204. [Google Scholar] [CrossRef] [Green Version]
- Loconto, J.; Papes, F.; Chang, E.; Stowers, L.; Jones, E.P.; Takada, T.; Kumánovics, A.; Lindahl, K.F.; Dulac, C. Functional Expression of Murine V2R Pheromone Receptors Involves Selective Association with the M10 and M1 Families of MHC Class Ib Molecules. Cell 2003, 112, 607–618. [Google Scholar] [CrossRef] [Green Version]
- Mangold, C.A.; Masser, D.R.; Stanford, D.R.; Bixler, G.V.; Pisupati, A.; Giles, C.B.; Wren, J.D.; Ford, M.M.; Sonntag, W.E.; Freeman, W.M. CNS-wide Sexually Dimorphic Induction of the Major Histocompatibility Complex 1 Pathway with Aging. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2017, 72, 16–29. [Google Scholar] [CrossRef] [Green Version]
- Barrios, A.W.; Nunez, G.; Quinteiro, P.S.; Salazar, I. Anatomy, histochemistry, and immunohistochemistry of the olfactory subsystems in mice. Front. Neuroanat. 2014, 63, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kovacs, T. Mechanisms of olfactory dysfunction in aging and neurodegenerative disorders. Ageing Res. Rev. 2004, 3, 215–232. [Google Scholar] [CrossRef]
- Doty, R.; Kamath, V. The influences of age on olfaction: A review. Front. Psychol. 2014, 5, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Host | Clonality | Dilution | Reference | Supplier |
|---|---|---|---|---|---|
| Anti-Olfactory Marker Protein (OMP) | Rabbit | Polyclonal | 1:10,000 | O7889 | Sigma, Saint-Louis, MO, USA |
| Anti-Gαi2 protein | Rabbit | Monoclonal | 1:200 | Ab157204 | Abcam, Cambridge, UK |
| Anti-Gαo protein | Goat | Polyclonal | 1:200 | Sc26769 | Santa Cruz Biotechnologies, CA, USA |
| Anti-Cleaved Caspase 3 | Rabbit | Polyclonal | 1:1000 | 9661 | Cell Signaling Technology, Danvers, MA, USA |
| Anti-Ki-67 | Rabbit | Monoclonal | 1:50 | MA514520 | Life Technologies, Carlsbad, CA, USA |
| Antibody | Targeted Cells/Molecules | Quantification Method |
|---|---|---|
| Anti-Olfactory Marker protein (OMP) | Mature neurons quantification | % of positivity signal/VNSE surface |
| Anti-Gαi2 protein | V1R neurons | % of positivity signal/VNSE surface |
| Anti-Gαo protein | V2R neurons | % of positivity signal/VNSE surface |
| Anti-Cleaved Caspase 3 | Apoptosis | Cell number/VNSE surface |
| Anti-Ki-67 | Proliferation activity | Cell number/VNSE surface |
| Studied Parameters | 3 Months (n = 20) | 10 Months (n = 22) | 24 Months (n = 40) |
|---|---|---|---|
| Cellular Degeneration | n (%) | ||
| non-existent | 2 (10.0) | 5 (22.7) | 0 (0.0) |
| weak | 13 (65.0) | 13 (59.1) | 15 (37.5) |
| moderate | 5 (25.0) | 4 (18.2) | 17 (42.5) |
| strong | 0 (0.0) | 0 (0.0) | 8 (20.0) |
| % area (stderr) | |||
| PAS positivity | 4.2 (0.5) | 6.3 (0.4) | 9.9 (0.4) |
| OMP positivity | 75.3 (1.6) | 69.3 (2.2) | 69.3 (1.1) |
| Gαi2 positivity | 39.1 (2.2) | 42.7 (1.6) | 20.8 (2.1) |
| Gαo positivity | 6.1 (0.5) | 13.2 (0.5) | 18.6 (0.9) |
| % positive cells (stderr) | |||
| Proliferative cells | 0.4 (0.2) | 0.3 (0.1) | 0.1 (0.1) |
| Apoptotic cells | 1.0 (0.2) | 1.1 (0.2) | 1.8 (0.2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mechin, V.; Pageat, P.; Teruel, E.; Asproni, P. Histological and Immunohistochemical Characterization of Vomeronasal Organ Aging in Mice. Animals 2021, 11, 1211. https://doi.org/10.3390/ani11051211
Mechin V, Pageat P, Teruel E, Asproni P. Histological and Immunohistochemical Characterization of Vomeronasal Organ Aging in Mice. Animals. 2021; 11(5):1211. https://doi.org/10.3390/ani11051211
Chicago/Turabian StyleMechin, Violaine, Patrick Pageat, Eva Teruel, and Pietro Asproni. 2021. "Histological and Immunohistochemical Characterization of Vomeronasal Organ Aging in Mice" Animals 11, no. 5: 1211. https://doi.org/10.3390/ani11051211