
A Moderate Reduction of Dietary Crude Protein Provide Comparable Growth Performance and Improve Metabolism via Changing Intestinal Microbiota in Sushan Nursery Pigs


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design, Animals, and Diets
2.2. Measurement and Sampling
2.3. Chemical Analysis
2.4. Fecal Bacterial DNA Isolation, MiSeq Sequencing, and Bioinformatics Analysis
2.5. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Apparent Total Tract Digestibility
3.3. Serum Parameters
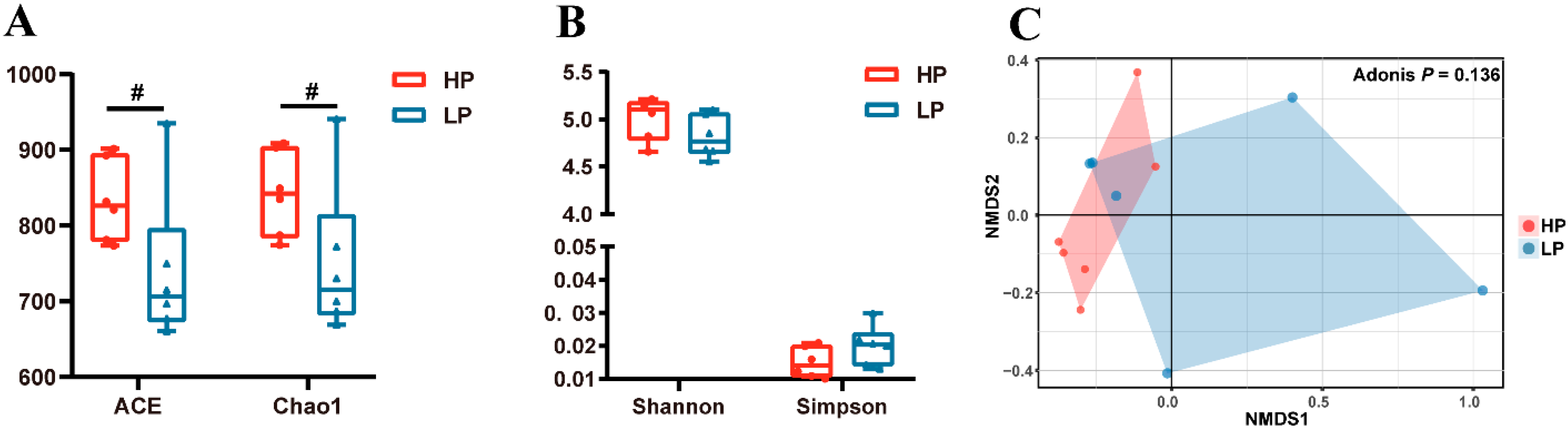
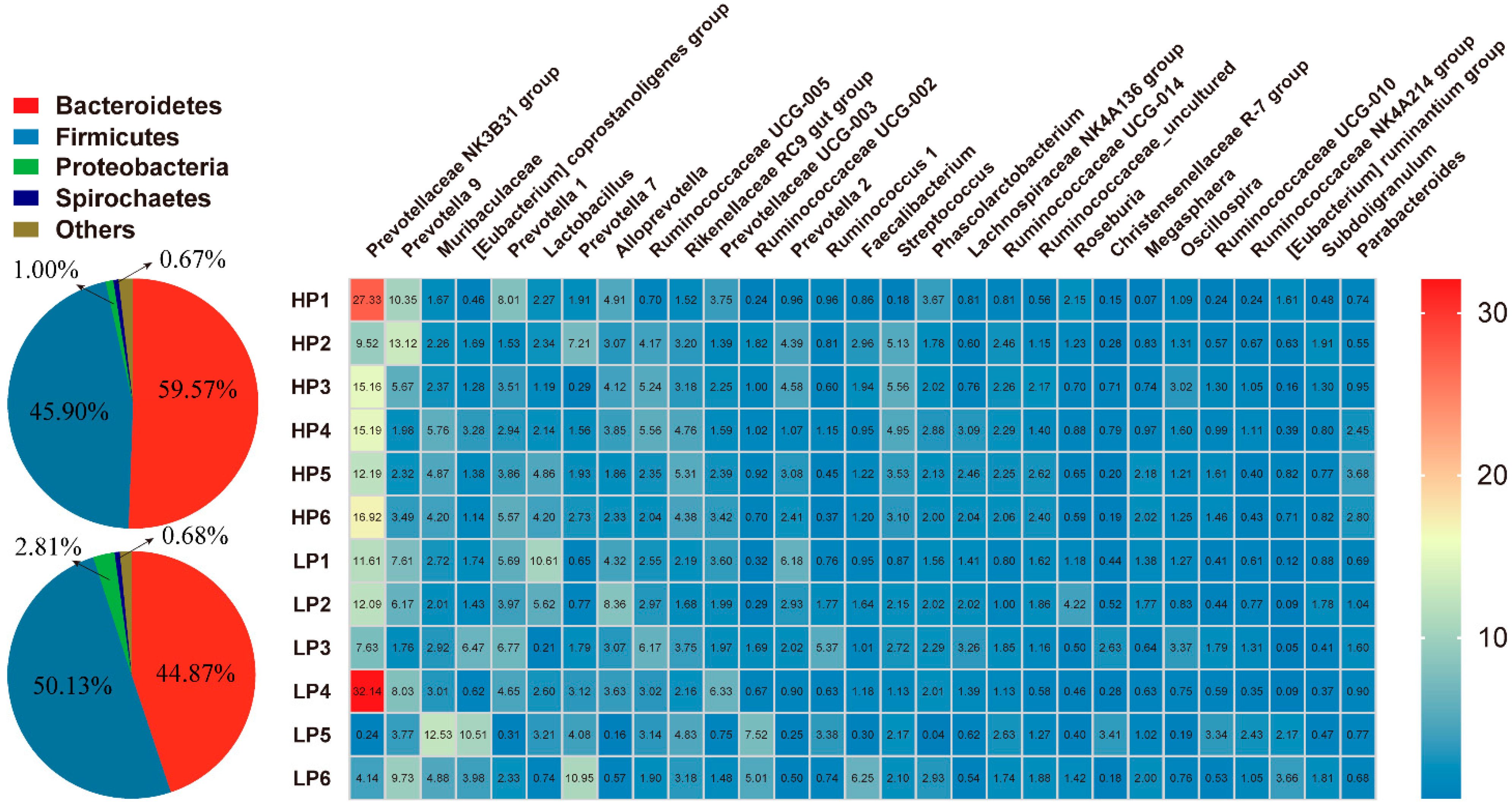
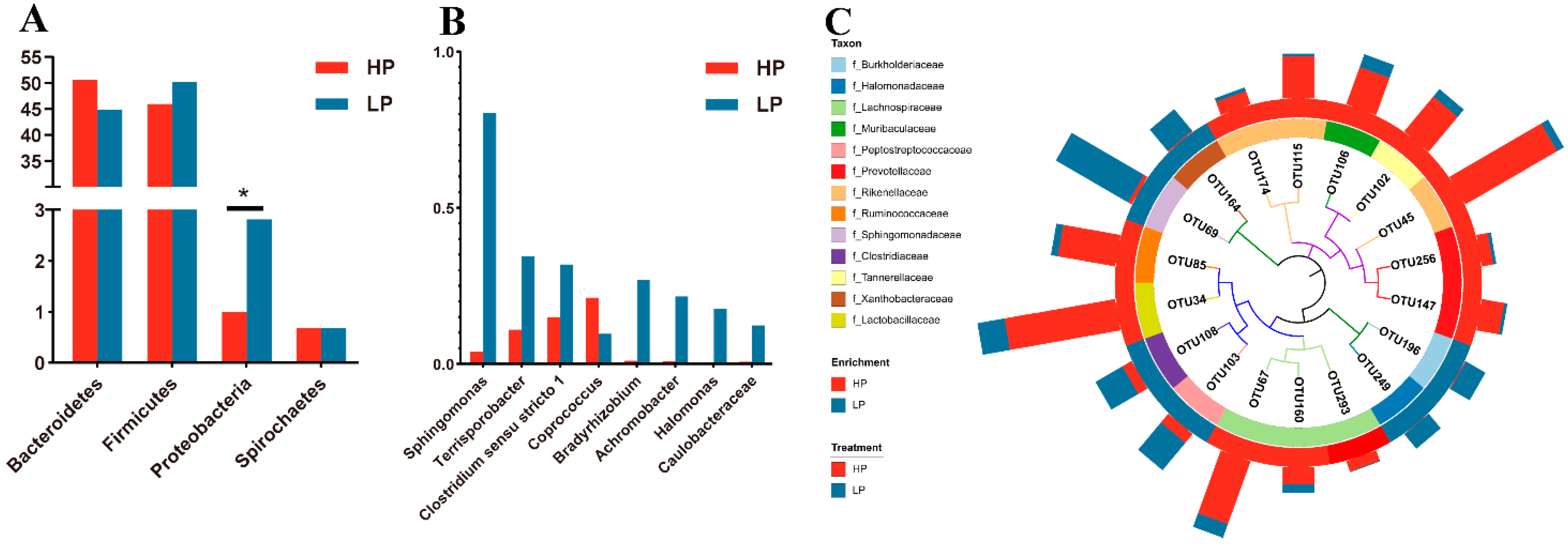
3.4. Fecal Microbial Community
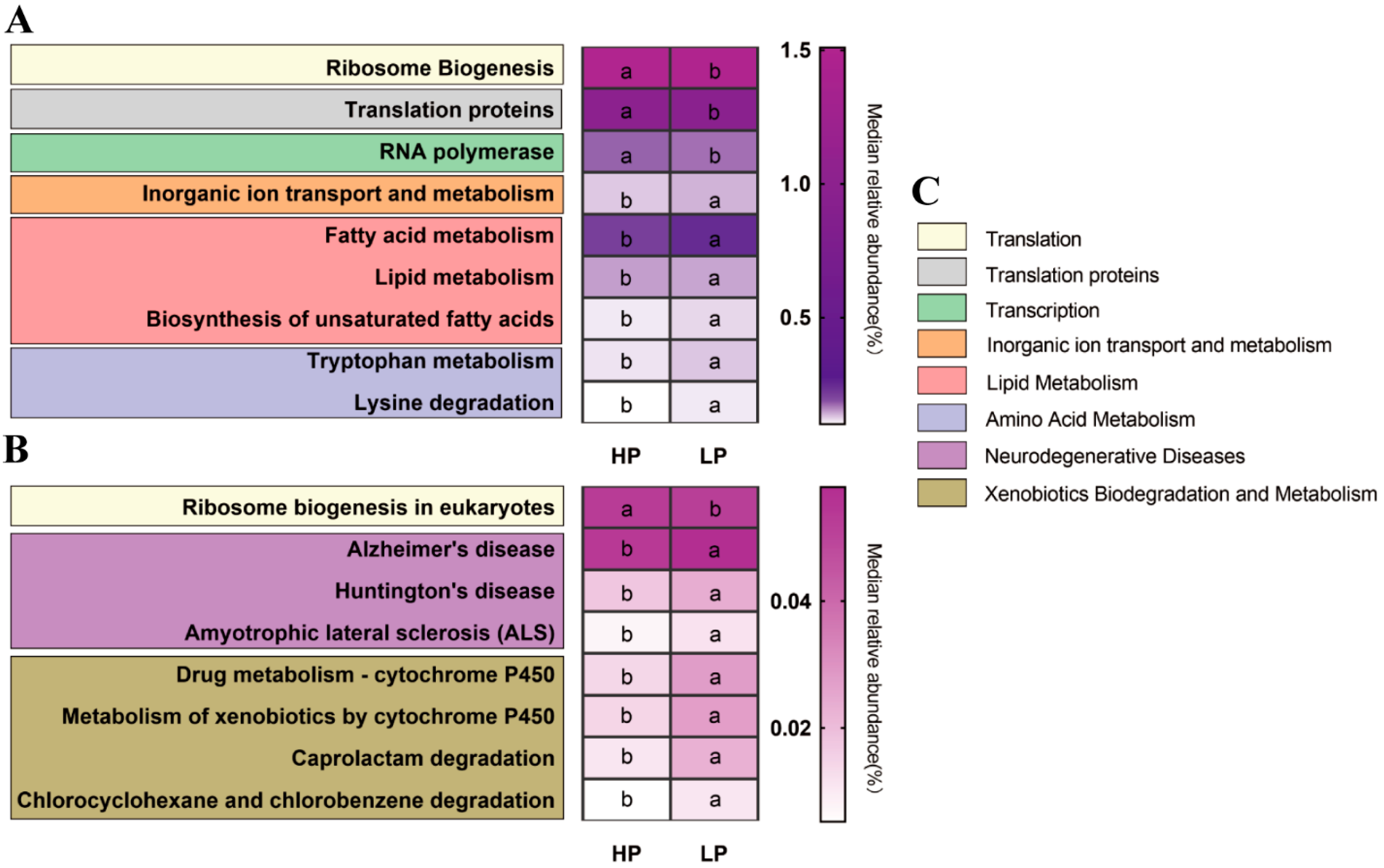
3.5. The Predicted Microbial Gene Functions of the Fecal Microbiota
3.6. Fecal SCFA Concentrations
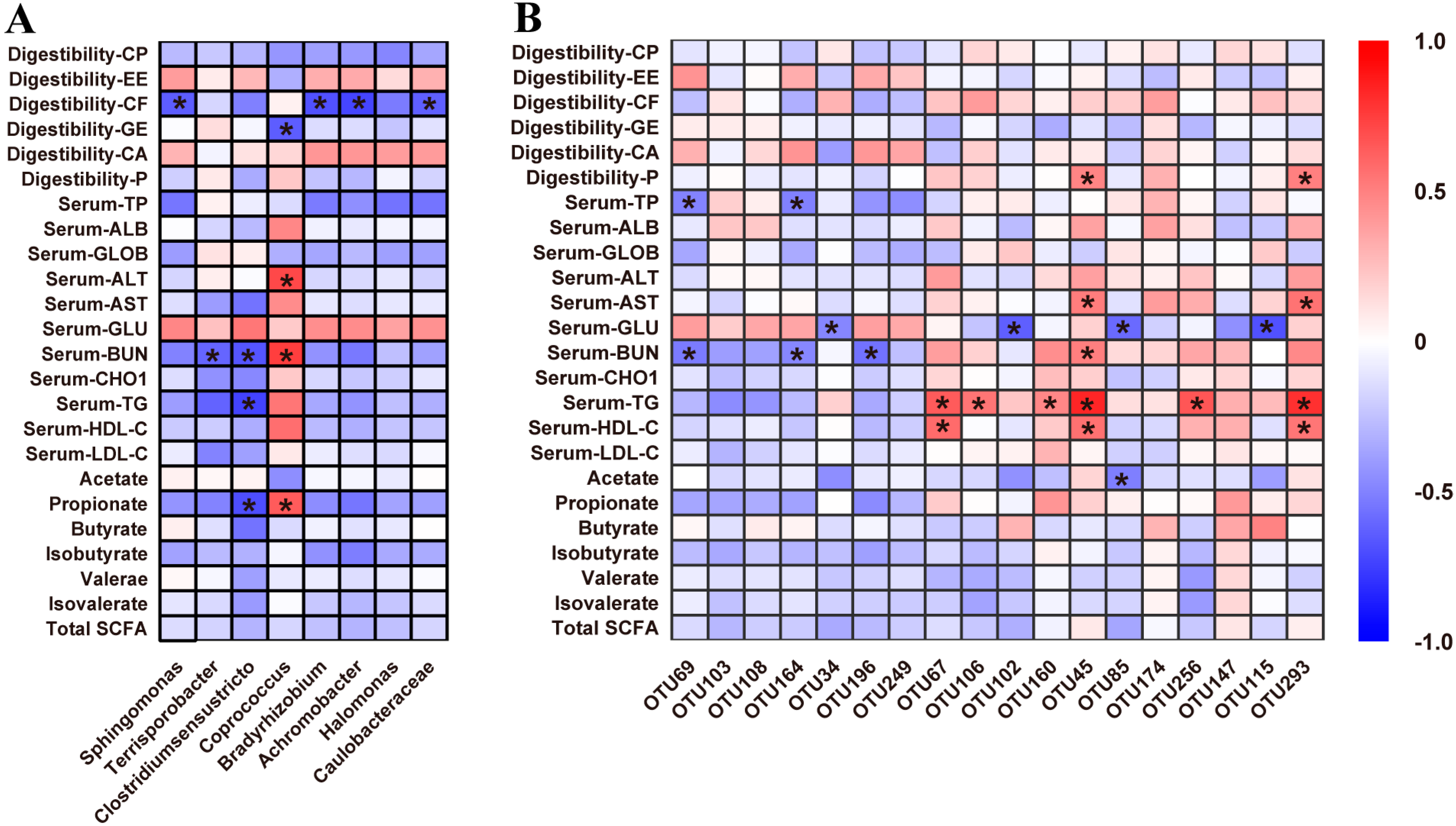
3.7. Correlations between Fecal Microbiota and Metabolic Profiles
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Orlando, U.A.D.; Oliveira, R.F.M.d.; Donzele, J.L.; Silva, F.C.D.O.; Generoso, R.A.R.; Siqueira, J.C.D. Dietary crude protein levels and amino acid supplementation for gilts from 30 to 60 kg maintained in a high environmental temperature. R. Bras. Zootec. 2007, 36, 1573–1578. [Google Scholar] [CrossRef] [Green Version]
- Webb, J.; Broomfield, M.; Jones, S.; Donovan, B. Ammonia and odour emissions from UK pig farms and nitrogen leaching from outdoor pig production. A review. Sci. Total Environ. 2014, 470, 865–875. [Google Scholar] [CrossRef]
- Portejoie, S.; Dourmad, J.Y.; Martinez, J.; Lebreton, Y. Effect of lowering dietary crude protein on nitrogen excretion, manure composition and ammonia emission from fattening pigs. Livest. Prod. Sci. 2004, 91, 45–55. [Google Scholar] [CrossRef]
- Heo, J.M.; Kim, J.C.; Hansen, C.F.; Mullan, B.P.; Hampson, D.J.; Pluske, J.R. Effects of dietary protein level and zinc oxide supplementation on performance responses and gastrointestinal tract characteristics in weaner pigs challenged with an enterotoxigenic strain of Escherichia coli. Anim. Prod. Sci. 2010, 50, 827–836. [Google Scholar] [CrossRef]
- Portune, K.J.; Beaumont, M.; Davila, A.M.; Tomé, D.; Blachier, F.; Sanz, Y. Gut microbiota role in dietary protein metabolism and health-related outcomes: The two sides of the coin. Trends Food Sci. Technol. 2016, 213–232. [Google Scholar] [CrossRef] [Green Version]
- Round, J.L.; Mazmanian, S.K. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 2009, 9, 600. [Google Scholar] [CrossRef]
- Rist, V.T.S.; Weiss, E.; Eklund, M.; Mosenthin, R. Impact of dietary protein on microbiota composition and activity in the gastrointestinal tract of piglets in relation to gut health: A review. Animal 2013, 7, 1067–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ball, R.O.; Aherne, F.X. Influence of dietary nutrient density, level of feed intake and weaning age on young pigs. II. Apparent nutrient digestibility and incidence and severity of diarrhea. Can. J. Anim. Sci. 1987, 67, 1105–1115. [Google Scholar] [CrossRef]
- Heo, J.M.; Kim, J.C.; Hansen, C.F.; Mullan, B.P.; Hampson, D.J.; Pluske, J.R. Effects of feeding low protein diets to piglets on plasma urea nitrogen, faecal ammonia nitrogen, the incidence of diarrhoea and performance after weaning. Arch. Anim. Nutr. 2008, 62, 343–358. [Google Scholar] [CrossRef]
- Acciaioli, A.; Sirtori, F.; Pianaccioli, L.; Campodoni, G.; Pugliese, C.; Bozzi, R.; Franci, O. Comparison of total tract digestibility and nitrogen balance between Cinta Senese and Large White pigs fed on different levels of dietary crude protein. Anim. Feed Sci. Technol. 2011, 169, 134–139. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Fang, L.; Sun, Y.; Su, Y.; Zhu, W. Effects of the dietary protein level on the microbial composition and metabolomic profile in the hindgut of the pig. Anaerobe 2016, 38, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Opapeju, F.O.; Rademacher, M.; Blank, G.; Nyachoti, C.M. Effect of low-protein amino acid-supplemented diets on the growth performance, gut morphology, organ weights and digesta characteristics of weaned pigs. Animal 2008, 2, 1457–1464. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Zhu, W.; Hang, S. Effects of low-protein diet on the intestinal morphology, digestive enzyme activity, blood urea nitrogen, and gut microbiota and metabolites in weaned pigs. Arch. Anim. Nutr. 2019, 73, 287–305. [Google Scholar] [CrossRef]
- Nyachoti, C.; Omogbenigun, F.; Rademacher, M.; Blank, G. Performance responses and indicators of gastrointestinal health in early-weaned pigs fed low-protein amino acid-supplemented diets. J. Anim. Sci. 2006, 84, 125–134. [Google Scholar] [CrossRef]
- Gu, X.; Li, D. Effect of dietary crude protein level on villous morphology, immune status and histochemistry parameters of digestive tract in weaning piglets. Anim. Feed Sci. Technol. 2004, 114, 113–126. [Google Scholar] [CrossRef]
- Zhou, L.; Zhao, W.; Fu, Y.; Fang, X.; Ren, S.; Ren, J. Genome-wide detection of genetic loci and candidate genes for teat number and body conformation traits at birth in Chinese Sushan pigs. Anim. Genet. 2019, 50, 753–756. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Nutrient Requirements of Swine, 11th ed.; The National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- AOAC. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2007. [Google Scholar]
- Hess, M.; Sczyrba, A.; Egan, R.; Kim, T.W.; Chokhawala, H.; Schroth, G.; Luo, S.; Clark, D.S.; Chen, F.; Zhang, T. Metagenomic discovery of biomass-degrading genes and genomes from cow rumen. Science 2011, 331, 463–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, W. Determination of rumen volatile fatty acids by means of gas chromatography. J. Nanjing Agric. College 1982, 4, 110–116. [Google Scholar]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas, M.N.; Burton, O.T.; Wise, P.; Zhang, Y.; Hobson, S.A.; Lloret, M.G.; Chehoud, C.; Kuczynski, J.; DeSantis, T.; Warrington, J. A microbiota signature associated with experimental food allergy promotes allergic sensitization and anaphylaxis. J. Allergy Clin. Immunol. 2013, 131, 201–212. [Google Scholar] [CrossRef]
- He, J.; Zheng, W.; Tao, C.; Guo, H.; Xue, Y.; Zhao, R.; Yao, W. Heat stress during late gestation disrupts maternal microbial transmission with altered offspring’s gut microbial colonization and serum metabolites in a pig model. Environ. Pollut. 2020, 266, 115111. [Google Scholar] [CrossRef]
- Cheng, J.; Ringel-Kulka, T.; Heikamp-de Jong, I.; Ringel, Y.; Carroll, I.; de Vos, W.M.; Salojärvi, J.; Satokari, R. Discordant temporal development of bacterial phyla and the emergence of core in the fecal microbiota of young children. ISME J. 2016, 10, 1002–1014. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods. 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Gallo, L.; Dalla Montà, G.; Carraro, L.; Cecchinato, A.; Carnier, P.; Schiavon, S. Growth performance of heavy pigs fed restrictively diets with decreasing crude protein and indispensable amino acids content. Livest. Sci. 2014, 161, 130–138. [Google Scholar] [CrossRef]
- Yin, Y.; Tan, B. Manipulation of dietary nitrogen, amino acids and phosphorus to reduce environmental impact of swine production and enhance animal health. J. Food Agric. Environ. 2010, 8, 447–462. [Google Scholar]
- Wu, G. Dietary requirements of synthesizable amino acids by animals: A paradigm shift in protein nutrition. J. Anim. Sci. Biotechnol. 2014, 5, 34. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.T.; Ma, W.F.; Zeng, X.F.; Xie, C.Y.; Thacker, P.A.; Htoo, J.K.; Qiao, S.Y. Estimation of the standardized ileal digestible valine to lysine ratio required for 25- to 120- kilogram pigs fed low crude protein diets supplemented with crystalline amino acids. J. Anim. Sci. 2015, 93, 4761–4773. [Google Scholar] [CrossRef]
- Li, Y.H.; Li, F.N.; Wu, L.; Liu, Y.Y.; Wei, H.K.; Li, T.J.; Tan, B.E.; Kong, X.F.; Wu, F.; Duan, Y.H.; et al. Reduced dietary protein level influences the free amino acid and gene expression profiles of selected amino acid transceptors in skeletal muscle of growing pigs. J. Anim. Physiol. Anim. Nutr. 2017, 101, 96–104. [Google Scholar] [CrossRef]
- Coma, J.; Carrion, D.; Zimmerman, D.R. Use of plasma urea nitrogen as a rapid response criterion to determine the lysine requirement of pigs. J. Anim. Sci. 1995, 73, 472–481. [Google Scholar] [CrossRef] [Green Version]
- Kohn, R.A.; Dinneen, M.M.; Russek-Cohen, E. Using blood urea nitrogen to predict nitrogen excretion and efficiency of nitrogen utilization in cattle, sheep, goats, horses, pigs, and rats. J. Anim. Sci. 2005, 83, 879–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stojanov, S.; Berlec, A.; Trukelj, B. The Influence of Probiotics on the Firmicutes/Bacteroidetes Ratio in the Treatment of Obesity and Inflammatory Bowel disease. Microorganisms 2020, 8, 1715. [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Jeong, M.Y.; Jang, H.M.; Kim, D.H. High-fat diet causes psychiatric disorders in mice by increasing Proteobacteria population. Neurosci. Lett. 2019, 698, 51–57. [Google Scholar] [CrossRef]
- Yun, L.; Yu, Z.; Li, Y.; Luo, P.; Jiang, X.; Tian, Y.; Ding, X. Ammonia nitrogen and nitrite removal by a heterotrophic Sphingomonas sp. strain LPN080 and its potential application in aquaculture. Aquaculture 2019, 500, 477–484. [Google Scholar] [CrossRef]
- Cho, H.U.; Kim, Y.M.; Park, J.M. Changes in microbial communities during volatile fatty acid production from cyanobacterial biomass harvested from a cyanobacterial bloom in a river. Chemosphere 2018, 202, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.G.; Sakamoto, K.; Seo, S.U.; Pickard, J.M.; Gillilland, M.G.; Pudlo, N.A.; Hoostal, M.; Li, X.; Wang, T.D.; Feehley, T.; et al. Neonatal acquisition of Clostridia species protects against colonization by bacterial pathogens. Science 2017, 356, 315–319. [Google Scholar] [CrossRef] [Green Version]
- Fan, P.; Liu, P.; Song, P.; Chen, X.; Ma, X. Moderate dietary protein restriction alters the composition of gut microbiota and improves ileal barrier function in adult pig model. Sci. Rep. 2017, 7, 43412. [Google Scholar] [CrossRef]
- Mesbah, N.M. Natronovirga. Bergey’s Man. Syst. Archaea Bact. 2019, 1–4. [Google Scholar] [CrossRef]
- Le Sciellour, M.; Zemb, O.; Serviento, A.M.; Renaudeau, D. Transient effect of single or repeated acute deoxynivalenol and zearalenone dietary challenge on fecal microbiota composition in female finishing pigs. Animal 2020, 14, 2277–2287. [Google Scholar] [CrossRef]
- Wilson, I.D.; Nicholson, J.K. Gut microbiome interactions with drug metabolism, efficacy, and toxicity. Transl. Res. 2017, 179, 204–222. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Zhou, G.; Wang, C.; Xu, X.; Li, C. Temporal changes in gut microbiota and signaling molecules of the gut-brain axis in mice fed meat protein diets. bioRxiv 2018, 329953. [Google Scholar] [CrossRef]
- Zhao, Y.; Tian, G.; Chen, D.; Zheng, P.; Yu, J.; He, J.; Mao, X.; Huang, Z.; Luo, Y.; Luo, J.; et al. Dietary protein levels and amino acid supplementation patterns alter the composition and functions of colonic microbiota in pigs. Anim. Nutr. 2020, 6, 143–151. [Google Scholar] [CrossRef]
- Spring, S.; Premathilake, H.; DeSilva, U.; Shili, C.; Carter, S.; Pezeshki, A. Low protein-high carbohydrate diets alter energy balance, gut microbiota composition and blood metabolomics profile in young pigs. Sci. Rep. 2020, 10, 3318. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Li, S.; Na, L.; Feng, R.; Liu, L.; Ying, L.; Sun, C.; Shirihai, O.S. Mangiferin decreases plasma free fatty acids through promoting its catabolism in liver by activation of AMPK. PLoS ONE 2012, 7, e30782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.H.; Wang, Y.C.; Chiu, C.C.; Lee, Y.P.; Hung, S.W.; Huang, C.C.; Chiu, C.F.; Chen, T.H.; Huang, W.C.; Chuang, H.L. Housing condition-associated changes in gut microbiota further affect the host response to diet-induced nonalcoholic fatty liver. J. Nutr. Biochem. 2020, 79, 108362. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Ishaq, S.L.; Zhao, F.Q.; Wright, A.D.G. Colonic inflammation accompanies an increase of β-catenin signaling and Lachnospiraceae/Streptococcaceae bacteria in the hind gut of high-fat diet-fed mice. J. Nutr. Biochem. 2016, 35, 30–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Dietary Protein Level 1 | |
|---|---|---|
| HP | LP | |
| Ingredients, g | ||
| Corn | 597.5245 | 672.7734 |
| Soybean meal | 257.9598 | 160.7904 |
| Wheat shorts | 50.00 | 50.00 |
| Rice bran meal | 30.00 | 30.00 |
| Corn germ meal | 30.00 | 30.00 |
| Dicalcium phosphate | 4.00 | 17.7298 |
| Limestone | 10.00 | 10.00 |
| Salt | 5.00 | 5.00 |
| Premix 2 | 13.275 | 13.275 |
| L-Lysine | 1.3385 | 4.6934 |
| L-Threonine | 0.4087 | 1.9448 |
| DL-Methionine | 0.3907 | 1.3430 |
| L-Tryptophan | 0.1028 | 0.5796 |
| Valine | 0.00 | 1.8706 |
| Total | 1000.00 | 1000.00 |
| Analyzed nutrient levels, % | ||
| Crude protein | 18.89 | 15.87 |
| Ether extract | 2.63 | 2.80 |
| Acid detergent fiber | 11.20 | 5.91 |
| Neutral detergent fiber | 9.43 | 5.17 |
| Crude fiber | 3.79 | 3.48 |
| Calcium | 0.61 | 0.61 |
| Phosphorus | 0.53 | 0.67 |
| Lysine | 1.22 | 1.23 |
| Methionine | 0.28 | 0.31 |
| Tryptophan | 0.21 | 0.22 |
| Threonine | 0.84 | 0.87 |
| Valine | 1.01 | 1.03 |
| Calculated nutrient levels 3, % | ||
| NE 4 MJ/kg | 10.19 | 10.21 |
| Items | Diets | p-Value | |
|---|---|---|---|
| HP | LP | ||
| BW, kg | |||
| day 0 | 19.10 ± 0.35 | 20.03 ± 0.31 | 0.152 |
| day 14 | 26.00 ± 0.50 | 26.87 ± 0.52 | 0.230 |
| day 28 | 33.79 ± 0.70 | 34.97 ± 0.63 | 0.158 |
| 0–14 day | |||
| ADFI, kg | 1.01 ± 0.04 | 1.03 ± 0.03 | 0.736 |
| ADG, kg | 0.49 ± 0.02 | 0.49 ± 0.02 | 0.899 |
| F/G | 2.08 ± 0.07 | 2.11 ± 0.13 | 0.781 |
| 15–28 d | |||
| ADFI, kg | 1.26 ± 0.05 | 1.27 ± 0.05 | 0.842 |
| ADG, kg | 0.53 ± 0.03 | 0.58 ± 0.03 | 0.215 |
| F/G | 2.29 ± 0.18 | 2.24 ± 0.12 | 0.828 |
| 0–28 day | |||
| ADFI, kg | 1.13 ± 0.04 | 1.15 ± 0.04 | 0.765 |
| ADG, kg | 0.51 ± 0.02 | 0.53 ± 0.02 | 0.359 |
| F/G | 2.22 ± 0.06 | 2.16 ± 0.03 | 0.421 |
| Items | Diets | p-Value | |
|---|---|---|---|
| HP | LP | ||
| Gross energy | 86.75 ± 0.56 | 88.13 ± 0.49 | 0.083 |
| Crude protein | 80.97 ± 0.94 | 81.24 ± 1.21 | 0.865 |
| Ether extract | 70.74 ± 1.40 | 75.45 ± 1.03 | 0.015 |
| Crude fiber | 51.01 ± 5.03 | 45.01 ± 3.39 | 0.336 |
| Calcium | 48.57 ± 3.16 | 37.61 ± 3.40 | 0.030 |
| Phosphorus | 33.28 ± 3.47 | 27.46 ± 3.17 | 0.233 |
| Items | Diets | p-Value | |
|---|---|---|---|
| HP | LP | ||
| Total protein, g/L | 68.07 ± 1.26 | 67.83 ± 1.69 | 0.911 |
| Albumin, g/L | 28.58 ± 1.30 | 26.50 ± 1.04 | 0.227 |
| Globulin, g/L | 39.49 ± 1.55 | 41.33 ± 2.41 | 0.529 |
| ALT, U/L | 46.67 ± 4.28 | 45.17 ± 3.61 | 0.835 |
| AST, U/L | 48.83 ± 2.34 | 49.17 ± 3.34 | 0.929 |
| Glucose, mmol/L | 4.84 ± 0.31 | 5.33 ± 0.16 | 0.182 |
| Urea nitrogen, mmol/L | 4.54 ± 0.31 | 2.80 ± 0.17 | <0.001 |
| Cholesterol, mmol/L | 2.49 ± 0.10 | 2.58 ± 0.14 | 0.633 |
| Triglyceride, mmol/L | 0.68 ± 0.07 | 0.53 ± 0.04 | 0.087 |
| HDLC, mmol/L | 0.84 ± 0.06 | 0.85 ± 0.03 | 0.928 |
| LDLC, mmol/L | 1.21 ± 0.05 | 1.27 ± 0.08 | 0.476 |
| Items | Diets | p-Value | |
|---|---|---|---|
| HP | LP | ||
| Acetate | 39.54 ± 2.62 | 46.72 ± 2.03 | 0.046 |
| Propionate | 18.87 ± 0.77 | 16.83 ± 1.27 | 0.191 |
| Butyrate | 8.91 ± 0.54 | 9.01 ± 1.05 | 0.937 |
| Isobutyrate | 2.16 ± 0.42 | 2.56 ± 1.74 | 0.558 |
| Valerate | 2.90 ± 0.28 | 3.30 ± 0.59 | 0.538 |
| Isovalerate | 2.59 ± 0.31 | 2.45 ± 0.34 | 0.764 |
| Total SCFA | 74.50 ± 3.34 | 80.90 ± 4.79 | 0.290 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, R.; He, J.; Ji, X.; Zheng, W.; Yao, W. A Moderate Reduction of Dietary Crude Protein Provide Comparable Growth Performance and Improve Metabolism via Changing Intestinal Microbiota in Sushan Nursery Pigs. Animals 2021, 11, 1166. https://doi.org/10.3390/ani11041166
Liu R, He J, Ji X, Zheng W, Yao W. A Moderate Reduction of Dietary Crude Protein Provide Comparable Growth Performance and Improve Metabolism via Changing Intestinal Microbiota in Sushan Nursery Pigs. Animals. 2021; 11(4):1166. https://doi.org/10.3390/ani11041166
Chicago/Turabian StyleLiu, Riliang, Jianwen He, Xu Ji, Weijiang Zheng, and Wen Yao. 2021. "A Moderate Reduction of Dietary Crude Protein Provide Comparable Growth Performance and Improve Metabolism via Changing Intestinal Microbiota in Sushan Nursery Pigs" Animals 11, no. 4: 1166. https://doi.org/10.3390/ani11041166