
Evaluation of Dietary Soluble Extract Hydrolysates with or without Supplementation of Inosine Monophosphate Based on Growth, Hematology, Non-Specific Immune Responses and Disease Resistance in Juvenile Nile Tilapia Oreochromis niloticus


 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Experimental Fish
2.3. Sample Collection and Analyses
2.4. Proximate Composition
2.5. Hematological Parameters
2.6. Non-Specific Immune Parameters
2.7. Challenge Test
2.8. Statistical Analysis
3. Results
3.1. Growth Performance and Survival Rate
3.2. Whole-Body Proximate Composition
3.3. Hematological Parameters
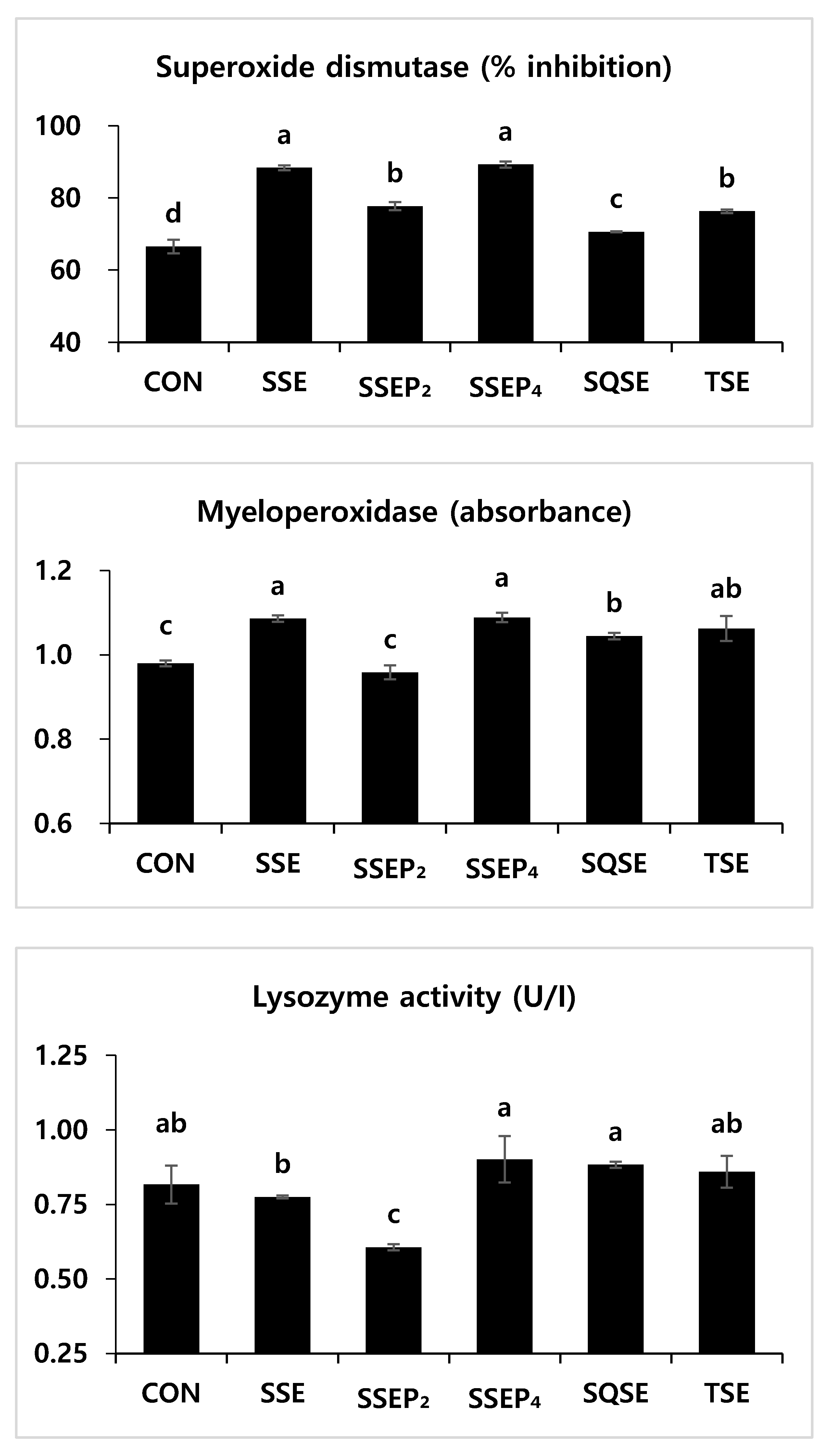
3.4. Non-Specific Immune Responses
3.5. Bacterial Challenge Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Food and Agriculture Organization FISHSTAT Plus; Universal Software for Fishery Statistical Time Series; Food and Agriculture Organization, United Nations: Rome, Italy, 2021. [Google Scholar]
- Moniruzzaman, M.; Bae, J.H.; Won, S.H.; Cho, S.J.; Chang, K.H.; Bai, S.C. Evaluation of solid-state fermented protein concentrates as a fish meal replacer in the diets of juvenile rainbow trout, Oncorhynchus mykiss. Aquacult. Nutr. 2018, 24, 1198–1212. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Damusaru, J.H.; Won, S.H.; Cho, S.J.; Chang, K.H.; Bai, S.C. Effects of partial replacement of dietary fish meal by bioprocessed plant protein concentrates on growth performance, hematology, nutrient digestibility and digestive enzyme activities in juvenile Pacific white shrimp, Litopenaeus Vannamei. J. Sci. Food Agric. 2020, 100, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Herath, S.S.; Haga, Y.; Satoh, S. Potential use of corn co-products in fishmeal-free diets for juvenile Nile tilapia Oreochromis niloticus. Fish. Sci. 2016, 82, 811–818. [Google Scholar] [CrossRef] [Green Version]
- Koch, J.F.; Rawles, S.D.; Webster, C.D.; Cummins, V.; Kobayashi, Y.; Thompson, K.R.; Gannam, A.L.; Twibell, R.G.; Hyde, N.M. Optimizing fish meal-free commercial diets for Nile tilapia, Oreochromis niloticus. Aquaculture 2016, 452, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Romano, N.; Kumar, V.; Yang, G.; Kajbaf, K.; Rubio, M.B.; Overturf, K.; Brezas, A.; Hardy, R. Bile acid metabolism in fish: Disturbances caused by fishmeal alternatives and some mitigating effects from dietary bile inclusions. Rev. Aquacult. 2020, 12, 1792–1817. [Google Scholar] [CrossRef]
- Toften, H.; Jobling, M. Feed intake and growth of Atlantic salmon, Salmo salar L., fed diets supplemented with oxytetracycline and squid extract. Aquacult. Nutr. 1997, 3, 145–151. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Kumar, B.D.; Hemalatha, R.; Jyothirmayi, T. Fish protein hydrolysates: Proximate composition, amino acid composition, antioxidant activities and applications: A review. Food Chem. 2012, 135, 3020–3038. [Google Scholar] [CrossRef]
- Gildberg, A. Enzymic processing of marine raw materials. Process Biochem. 1992, 28, 1–15. [Google Scholar] [CrossRef]
- Leduc, A.; Zatylny-Gaudin, C.; Khosravi, M.; Corre, E.; Le Corguille, G.; Castel, H.; Lefevre-Scelles, A.; Fournier, V.; Gisbert, E.; Andree, K.B.; et al. Dietary aquaculture by-product hydrolysates: Impact on the transcriptomic response of the intestinal mucosa of European seabass (Dicentrarchus labrax) fed low fish meal diets. BMC Genom. 2018, 19, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Plascencia-Jatomea, M.; Olvera-Novoa, M.A.; Arredondo-Figueroa, J.L.; Hall, G.M.; Shirai, K. Feasibility of fishmeal replacement by shrimp head silage protein hydrolysate in Nile tilapia (Oreochromis niloticus L.) diets. J. Sci. Food Agric. 2002, 82, 753–759. [Google Scholar] [CrossRef]
- Jo, H.; Lee, S.; Yun, H.; Hong, J.W.; Moniruzzaman, M.; Bai, S.C.; Park, G.; Chee, S.; Jeon, T.E. Evaluation of dietary fishmeal analogue with addition of shrimp soluble extract on growth and nonspecific immune response of rainbow trout, Oncorhynchus mykiss. J. World Aquacult. Soc. 2017, 48, 583–591. [Google Scholar] [CrossRef]
- Kolkovski, S.; Tandler, A. The use of squid protein hydrolysate as a protein source in microdiets for gilthead seabream Sparus aurata larvae. Aquacult. Nutr. 2000, 6, 11–16. [Google Scholar] [CrossRef]
- Foh, M.B.; Kamara, M.T.; Amadou, I.; Foh, B.M.; Xia, W. Chemical and physicochemical properties of tilapia (Oreochromis niloticus) fish protein hydrolysate and concentrate. Int. J. Biol. Chem. 2011, 5, 21–36. [Google Scholar] [CrossRef]
- Fan, J.; He, J.; Zhuang, Y.; Sun, L. Purification and identification of antioxidant peptides from enzymatic hydrolysates of tilapia (Oreochromis niloticus) frame protein. Molecules 2012, 17, 12836–12850. [Google Scholar] [CrossRef]
- Jobling, M.; Koskela, J.; Savolainen, R. Influence of dietary fat level and increased adiposity on growth and fat deposition in rainbow trout, Oncorhynchus mykiss (Walbaum). Aquacult. Res. 1998, 29, 601–607. [Google Scholar] [CrossRef]
- Lian, P.; Lee, C.M.; Park, E. Characterization of Squid-Processing Byproduct Hydrolysate and Its Potential as Aquaculture Feed Ingredient. J. Agric. Food Chem. 2005, 53, 5587–5592. [Google Scholar] [CrossRef] [PubMed]
- Gildberg, A.; Stenberg, E. A new process for advanced utilisation of shrimp waste. Process Biochem. 2001, 36, 809–812. [Google Scholar] [CrossRef]
- Aksnes, A.; Hope, B.; Jönsson, E.; Björnsson, B.T.; Albrektsen, S. Size-fractionated fish hydrolysate as feed ingredient for rainbow trout (Oncorhynchus mykiss) fed high plant protein diets. I: Growth, growth regulation and feed utilization. Aquaculture 2006, 261, 305–317. [Google Scholar] [CrossRef]
- El-Sayed, A.F. Tilapia Culture, 2nd ed.; Academic Press: Cambridge, MA, USA; Elsevier Publishers: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Abdel-Tawwab, M.; Abdel-Rahman, A.M.; Ismael, N.E. Evaluation of commercial live bakers’ yeast, Saccharomyces cerevisiae as a growth and immunity promoter for Fry Nile tilapia, Oreochromis niloticus (L.) challenged in situ with Aeromonas hydrophila. Aquaculture 2008, 280, 185–189. [Google Scholar] [CrossRef]
- Simon, C.J.; Blyth, D.; Ahmad, F.N.; Suri, S. Microbial biomass (Novacq™) stimulates feeding and improves the growth performance on extruded low to zero-fishmeal diets in tilapia (GIFT strain). Aquaculture 2019, 501, 319–324. [Google Scholar] [CrossRef]
- Li, P.; Gatlin, D.M., III. Nucleotide nutrition in fish: Current knowledge and future applications. Aquaculture 2006, 251, 141–152. [Google Scholar] [CrossRef]
- Carver, J.D. Dietary nucleotides: Cellular immune, intestinal and hepatic system effects. J. Nutr. 1994, 124, 144S–148S. [Google Scholar] [CrossRef]
- Haskó, G.; Kuhel, D.G.; Németh, Z.H.; Mabley, J.G.; Stachlewitz, R.F.; Virág, L.; Lohinai, Z.; Southan, G.J.; Salzman, A.L.; Szabó, C. Inosine inhibits inflammatory cytokine production by a posttranscriptional mechanism and protects against endotoxin-induced shock. J. Immunol. 2000, 164, 1013–1019. [Google Scholar] [CrossRef]
- Lin, Y.H.; Wang, H.; Shiau, S.Y. Dietary nucleotide supplementation enhances growth and immune responses of grouper, Epinephelus malabaricus. Aquacult. Nutr. 2009, 15, 117–122. [Google Scholar] [CrossRef]
- Hossain, M.S.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Sony, N.M.; Ono, S.; Fujieda, T. Comparison of the effects of inosine and inosine monophosphate on growth, immune response, stress resistance and gut morphology of juvenile red sea bream, Pagrus major. Aquaculture 2016, 458, 64–74. [Google Scholar] [CrossRef]
- Song, J.W.; Lim, S.J.; Lee, K.J. Effects of dietary supplementation of inosine monophosphate on growth performance, innate immunity and disease resistance of olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2012, 33, 1050–1054. [Google Scholar] [CrossRef]
- Asaduzzaman, M.; Ikeda, D.; Abol-Munafi, A.B.; Bulbul, M.; Ali, M.E.; Kinoshita, S.; Watabe, S.; Kader, M.A. Dietary supplementation of inosine monophosphate promotes cellular growth of muscle and upregulates growth-related gene expression in Nile tilapia Oreochromis niloticus. Aquaculture 2017, 468, 297–306. [Google Scholar] [CrossRef]
- Kader, M.A.; Bulbul, M.; Abol-Munafi, A.B.; Asaduzzaman, M.; Mian, S.; Noordin, N.B.M.; Ali, M.E.; Hossain, M.S.; Koshio, S. Modulation of growth performance, immunological responses and disease resistance of juvenile Nile tilapia (Oreochromis niloticus) (Linnaeus, 1758) by supplementing dietary inosine monophosphate. Aquacult. Rep. 2018, 10, 23–31. [Google Scholar] [CrossRef]
- Hamidoghli, A.; Won, S.; Farris, N.W.; Bae, J.; Choi, W.; Yun, H.; Bai, S.C. Solid state fermented plant protein sources as fish meal replacers in whiteleg shrimp Litopaeneus vannamei. Anim. Feed Sci. Technol. 2020, 264, 114474. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International; AOAC: Rockville, MD, USA, 2005. [Google Scholar]
- Hultmark, D.; Steiner, H.; Rasmuson, T.; Boman, H.G. Insect immunity. Purification and properties of three inducible bactericidal proteins from hemolymph of immunized pupae of Hyalophora cecropia. Eur. J. Biochem. 1980, 106, 7–16. [Google Scholar] [CrossRef]
- Quade, M.J.; Roth, J.A. A rapid, direct assay to measure degranulation of bovine neutrophil primary granules. Vet. Immunol. Immunopath. 1997, 58, 239–248. [Google Scholar] [CrossRef]
- Hasan, M.T.; Jang, W.J.; Lee, B.J.; Kim, K.W.; Hur, S.W.; Lim, S.G.; Bai, S.C.; Kong, I.S. Heat-killed Bacillus sp. SJ-10 probiotic acts as a growth and humoral innate immunity response enhancer in olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 2019, 88, 424–431. [Google Scholar] [CrossRef]
- Leal, A.L.G.; de Castro, P.F.; de Lima, J.P.V.; de Souza Correia, E.; de Souza Bezerra, R. Use of shrimp protein hydrolysate in Nile tilapia (Oreochromis niloticus, L.) feeds. Aquacult. Int. 2010, 18, 635–646. [Google Scholar] [CrossRef]
- Robert, M.; Zatylny-Gaudin, C.; Fournier, V.; Corre, E.; Le Corguille, G.; Bernay, B.; Henry, J. Transcriptomic and peptidomic analysis of protein hydrolysates from the white shrimp (L. vannamei). J. Biotechnol. 2014, 186, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Robert, M.; Zatylny-Gaudin, C.; Fournier, V.; Corre, E.; Le Corguille, G.; Bernay, B.; Henry, J. Molecular characterization of peptide fractions of a Tilapia (Oreochromis niloticus) by-product hydrolysate and in vitro evaluation of antibacterial activity. Process Biochem. 2015, 50, 487–492. [Google Scholar] [CrossRef]
- Hung, L.T. Shrimp soluble extract-novel feed attractant for aquaculture. Glob. Aquac. Advocate 2014, 17, 81–82. [Google Scholar]
- Khosravi, S.; Rahimnejad, S.; Herault, M.; Fournier, V.; Lee, C.R.; Bui, H.T.D.; Jeong, J.B.; Lee, K.J. Effects of protein hydrolysates supplementation in low fish meal diets on growth performance, innate immunity and disease resistance of red sea bream Pagrus major. Fish Shellfish Immunol. 2015, 45, 858–868. [Google Scholar] [CrossRef]
- Delcroix, J.; Gatesoupe, F.J.; Desbruyeres, E.; Huelvan, C.; Le Delliou, H.; Le Gall, M.M.; Quazuguel, P.; Mazurais, D.; Zambonino-Infante, J.L. The effects of dietary marine protein hydrolysates on the development of sea bass larvae, Dicentrarchus labrax, and associated microbiota. Aquacult. Nutr. 2015, 21, 98–104. [Google Scholar] [CrossRef] [Green Version]
- Burrells, C.; Williams, P.D.; Southgate, P.J.; Wadsworth, S.L. Dietary nucleotides: A novel supplement in fish feeds: 2. Effects on vaccination, salt water transfer, growth rates and physiology of Atlantic salmon (Salmo salar L.). Aquaculture 2001, 199, 171–184. [Google Scholar] [CrossRef]
- Tahmasebi-Kohyani, A.; Keyvanshokooh, S.; Nematollahi, A.; Mahmoudi, N.; Pasha-Zanoosi, H. Dietary administration of nucleotides to enhance growth, humoral immune responses, and disease resistance of the rainbow trout (Oncorhynchus mykiss) fingerlings. Fish Shellfish Immunol. 2011, 30, 189–193. [Google Scholar] [CrossRef]
- Zhang, P.; Fu, L.; Liu, H.; Huda, N.U.; Zhu, X.; Han, D.; Jin, J.; Yang, Y.; Kim, Y.S.; Xie, S. Effects of inosine 5′-monophosphate supplementation in high fishmeal and high soybean diets on growth, immune-related gene expression in gibel carp (Carassius auratus gibelio var. CAS Ⅲ), and its challenge against Aeromonas hydrophila infection. Fish Shellfish Immunol. 2019, 86, 913–921. [Google Scholar] [CrossRef]
- Khosravi, S.; Bui, H.T.D.; Herault, M.; Fournier, V.; Kim, K.D.; Kim, K.W.; Lee, K.J. Supplementation of protein hydrolysates to a low-fishmeal diet improves growth and health status of Juvenile Olive Flounder, Paralichthys olivaceus. J. World Aquacult. Soc. 2018, 49, 897–911. [Google Scholar] [CrossRef]
- Gisbert, E.; Fournier, V.; Solovyev, M.; Skalli, A.; Andree, K.B. Diets containing shrimp protein hydrolysates provided protection to European sea bass (Dicentrarchus labrax) affected by a Vibrio pelagius natural infection outbreak. Aquaculture 2018, 495, 136–143. [Google Scholar] [CrossRef]
- Maita, M. Fish Health Assessment. In Dietary Supplements for the Health and Quality of Cultured Fish; CAB International: Washington, DC, USA, 2007. [Google Scholar]
- Khosravi, S.; Bui, H.T.D.; Rahimnejad, S.; Herault, M.; Fournier, V.; Jeong, J.B.; Lee, K.J. Effect of dietary hydrolysate supplementation on growth performance, non-specific immune response and disease resistance of olive flounder (Paralichthys olivaceus) challenged with Edwardsiella tarda. Aquac. Nutr. 2015, 21, 321–331. [Google Scholar] [CrossRef]
- Sila, A.; Nedjar-Arroume, N.; Hedhili, K.; Chataigne, G.; Balti, R.; Nasri, M.; Dhulster, P.; Bougatef, A. Antibacterial peptides from barbel muscle protein hydrolysates: Activity against some pathogenic bacteria. LWT Food Sci. Technol. 2014, 55, 183–188. [Google Scholar] [CrossRef]
- García-Moreno, P.J.; Batista, I.; Pires, C.; Bandarra, N.M.; Espejo-Carpio, F.J.; Guadix, A.; Guadix, E.M. Antioxidant activity of protein hydrolysates obtained from discarded Mediterranean fish species. Food Res. Int. 2014, 65, 469–476. [Google Scholar] [CrossRef]
- Ktari, N.; Nasri, R.; Mnafgui, K.; Hamden, K.; Belguith, O.; Boudaouara, T.; El Feki, A.; Nasri, M. Antioxidative and ACE inhibitory activities of protein hydrolysates from zebra blenny (Salaria basilisca) in alloxan-induced diabetic rats. Process Biochem. 2014, 49, 890–897. [Google Scholar] [CrossRef]
- Ainsworth, A.J.; Dexiang, C.; Waterstrat, P.R. Changes in peripheral blood leukocyte percentages and function of neutrophils in stressed channel catfish. J. Aquat. Anim. Health 1991, 3, 41–47. [Google Scholar] [CrossRef]
- Bui, H.T.D.; Khosravi, S.; Fournier, V.; Herault, M.; Lee, K.J. Growth performance, feed utilization, innate immunity, digestibility and disease resistance of juvenile red seabream (Pagrus major) fed diets supplemented with protein hydrolysates. Aquaculture 2014, 418, 11–16. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Ingredients (% in Diet) | Diets | |||||
|---|---|---|---|---|---|---|
| CON | SSE | SSEP2 | SSEP4 | SQSE | TSE | |
| Fishmeal (Tuna) 1 | 8.00 | 8.00 | 8.00 | 8.00 | 8.00 | 8.00 |
| Soybean meal 1 | 34.60 | 34.00 | 34.00 | 34.00 | 34.00 | 34.00 |
| Wheat flour 1 | 34.99 | 33.54 | 33.54 | 33.54 | 33.54 | 33.54 |
| Rapeseed meal 1 | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 |
| Meat and bone meal 1 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 |
| Poultry offal meal 1 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 |
| Squid liver meal 1 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 |
| Soybean oil 1 | 1.30 | 1.34 | 1.34 | 1.34 | 1.34 | 1.34 |
| Fish oil 2 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 |
| Other 3 | 3.11 | 3.12 | 3.12 | 3.12 | 3.12 | 3.12 |
| SSE 4 | 2.00 | |||||
| SSE + IMP 5 2% | 2.00 | |||||
| SSE + IMP 4% | 2.00 | |||||
| SQSE 6 | 2.00 | |||||
| TSE 7 | 2.00 | |||||
| Total | 100 | 100 | 100 | 100 | 100 | 100 |
| Proximate Analysis (% of Dry Matter Basis) | ||||||
| Moisture | 9.42 | 9.90 | 9.84 | 9.22 | 9.82 | 9.08 |
| Crude protein | 33.32 | 32.64 | 32.90 | 33.25 | 32.40 | 32.33 |
| Crude lipid | 5.20 | 5.79 | 5.78 | 5.41 | 5.92 | 6.22 |
| Crude ash | 8.02 | 8.08 | 8.08 | 8.28 | 8.16 | 8.36 |
| Diets | ||||||
|---|---|---|---|---|---|---|
| CON | SSE | SSEP2 | SSEP4 | SQSE | TSE | |
| WG 2 | 254.01 ± 3.57 b | 264.24 ± 2.71 a | 264.71 ± 4.67 a | 264.95 ± 2.87 a | 253.49 ± 5.32 b | 256.84 ± 4.13 ab |
| SGR 3 | 2.63 ± 0.02 b | 2.69 ± 0.02 a | 2.70 ± 0.03 a | 2.70 ± 0.02 a | 2.63 ± 0.03 b | 2.65 ± 0.02 ab |
| FE 4 | 76.90 ± 0.93 ns | 71.41 ± 1.79 | 75.04 ± 3.66 | 74.21 ± 1.58 | 72.91 ± 2.19 | 72.62 ± 4.07 |
| FI 5 | 354.70 ± 7.52 b | 397.20 ± 9.12 a | 380.60 ± 5.03 ab | 384.70 ± 7.05 a | 373.40 ± 6.12 ab | 377.80 ± 8.50 ab |
| PER 6 | 2.09 ± 0.03 ns | 1.97 ± 0.05 | 2.06 ± 0.10 | 2.03 ± 0.04 | 2.03 ± 0.06 | 2.04 ± 0.11 |
| Survival 7 | 100.00 ± 0.00 ns | 100.00 ± 0.00 | 100.00 ± 0.00 | 100.00 ± 0.00 | 98.25 ± 2.48 | 98.25 ± 2.48 |
| HSI 8 | 0.69 ± 0.02 ns | 0.74 ± 0.06 | 0.72 ± 0.04 | 0.73 ± 0.10 | 0.71 ± 0.07 | 0.77 ± 0.09 |
| VSI 9 | 6.68 ± 0.04 ns | 6.76 ± 0.01 | 6.64 ± 0.20 | 6.70 ± 0.16 | 6.65 ± 0.14 | 6.75 ± 0.09 |
| CF 10 | 1.52 ± 0.07 ns | 1.55 ± 0.07 | 1.54 ± 0.01 | 1.52 ± 0.03 | 1.55 ± 0.01 | 1.54 ± 0.03 |
| Diets | ||||||
|---|---|---|---|---|---|---|
| CON | SSE | SSEP2 | SSEP4 | SQSE | TSE | |
| Moisture | 72.69 ± 0.35 ns | 71.88 ± 2.14 | 71.67 ± 1.37 | 71.03 ± 0.45 | 72.76 ± 0.65 | 72.66 ± 0.26 |
| Crude protein | 15.77 ± 0.22 ns | 16.20 ± 1.15 | 16.55 ± 0.81 | 16.58 ± 0.18 | 15.99 ± 0.40 | 15.87 ± 0.55 |
| Crude Lipid | 6.63 ± 0.77 ns | 6.29 ± 1.54 | 6.72 ± 0.63 | 7.39 ± 0.51 | 6.87 ± 0.67 | 6.36 ± 1.35 |
| Crude Ash | 4.56 ± 0.17 ns | 4.72 ± 0.20 | 5.08 ± 0.43 | 5.00 ± 0.32 | 4.77 ± 0.81 | 4.55 ± 0.31 |
| Diets | ||||||
|---|---|---|---|---|---|---|
| CON | SSE | SSEP2 | SSEP4 | SQSE | TSE | |
| AST 2 | 76.0 ± 4.6 ns | 77.0 ± 2.6 | 75.0 ± 2.0 | 75.0 ± 3.0 | 75.0 ± 2.0 | 74.7 ± 2.1 |
| ALT 3 | 3.0 ± 1.0 ns | 2.0 ± 0.0 | 3.0 ± 1.0 | 2.7 ± 0.6 | 2.7 ± 0.6 | 2.7 ± 0.6 |
| GLU 4 | 44.3 ± 2.3 ns | 43.0 ± 1.0 | 45.3 ± 1.5 | 44.0 ± 1.0 | 43.0 ± 1.0 | 43.0 ± 1.0 |
| TCHO 5 | 224.3 ± 0.6 ns | 224.0 ± 1.0 | 225.3 ± 1.5 | 224.3 ± 0.6 | 224.0 ± 1.7 | 225.3 ± 1.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, J.; Song, Y.; Moniruzzaman, M.; Hamidoghli, A.; Lee, S.; Je, H.; Choi, W.; Min, T.; Bai, S.C. Evaluation of Dietary Soluble Extract Hydrolysates with or without Supplementation of Inosine Monophosphate Based on Growth, Hematology, Non-Specific Immune Responses and Disease Resistance in Juvenile Nile Tilapia Oreochromis niloticus. Animals 2021, 11, 1107. https://doi.org/10.3390/ani11041107
Bae J, Song Y, Moniruzzaman M, Hamidoghli A, Lee S, Je H, Choi W, Min T, Bai SC. Evaluation of Dietary Soluble Extract Hydrolysates with or without Supplementation of Inosine Monophosphate Based on Growth, Hematology, Non-Specific Immune Responses and Disease Resistance in Juvenile Nile Tilapia Oreochromis niloticus. Animals. 2021; 11(4):1107. https://doi.org/10.3390/ani11041107
Chicago/Turabian StyleBae, Jinho, Yujin Song, Mohammad Moniruzzaman, Ali Hamidoghli, Seunghan Lee, Hyeongwoo Je, Wonsuk Choi, Taesun Min, and Sungchul C. Bai. 2021. "Evaluation of Dietary Soluble Extract Hydrolysates with or without Supplementation of Inosine Monophosphate Based on Growth, Hematology, Non-Specific Immune Responses and Disease Resistance in Juvenile Nile Tilapia Oreochromis niloticus" Animals 11, no. 4: 1107. https://doi.org/10.3390/ani11041107