
A Model for Dental Age Verification Using Ultrastructural Imaging for Modern and Fossil Representatives of the Rhinocerotidae Family



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Animal and Tooth Analyzed
2.2. Specimen Preparation
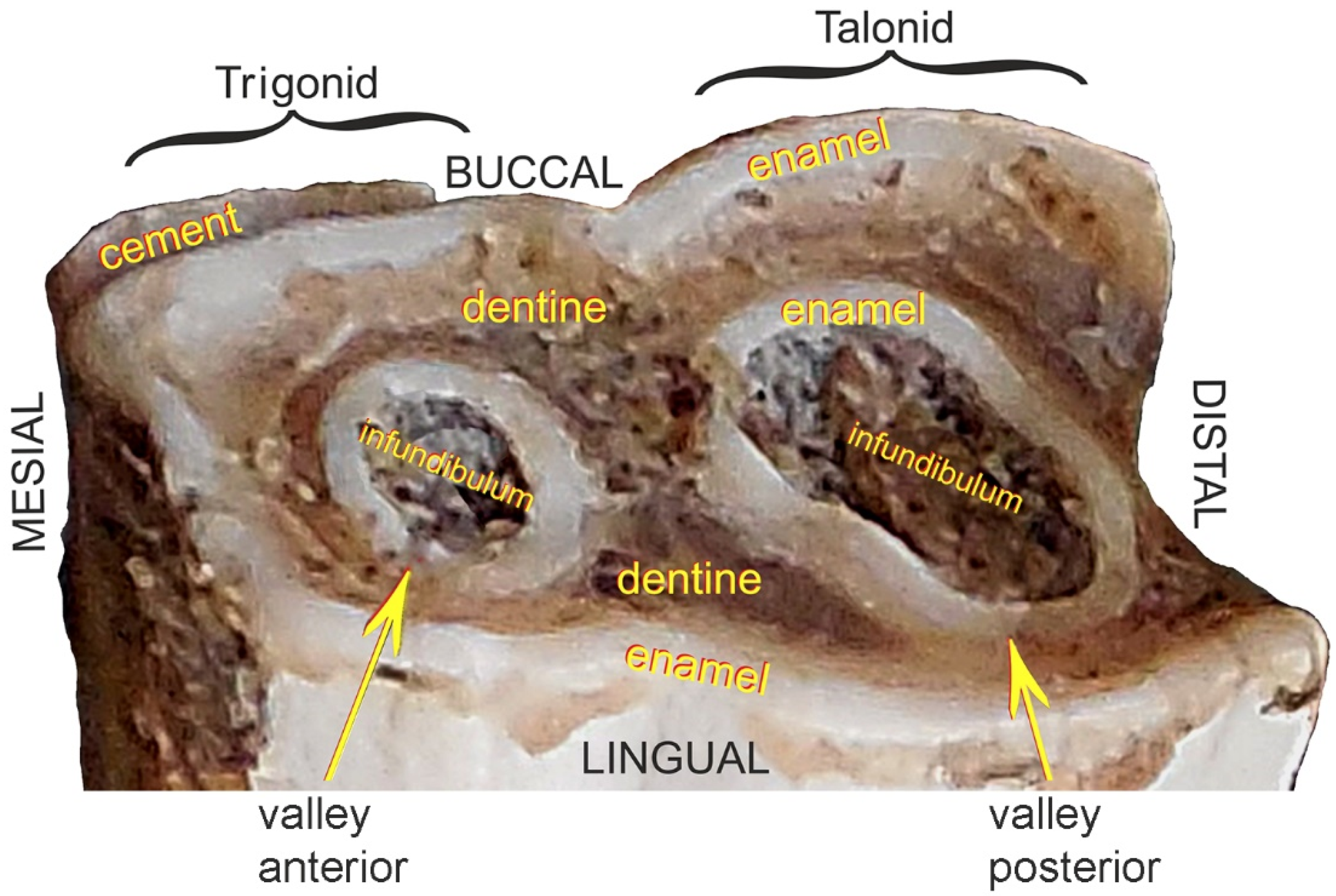
2.3. Analyzed Tooth Sections and Type of Equipment Used
- (a)
- horizontal in a mesial-distal plane through the tooth crown—the cutting plane positioned at a height of approximately 30 mm from the occlusal surface;
- (b)
- horizontal (axial, transverse) sections in a mesial-distal plane through the coronal portion (upper part) of the root—cutting planes positioned on the rostral and caudal root, at the height of the bifurcation of the tooth roots;
- (c)
- longitudinal sections in a lingual-buccal plane through the crown and the root—cutting planes routed through the middle of the rostral and caudal roots.
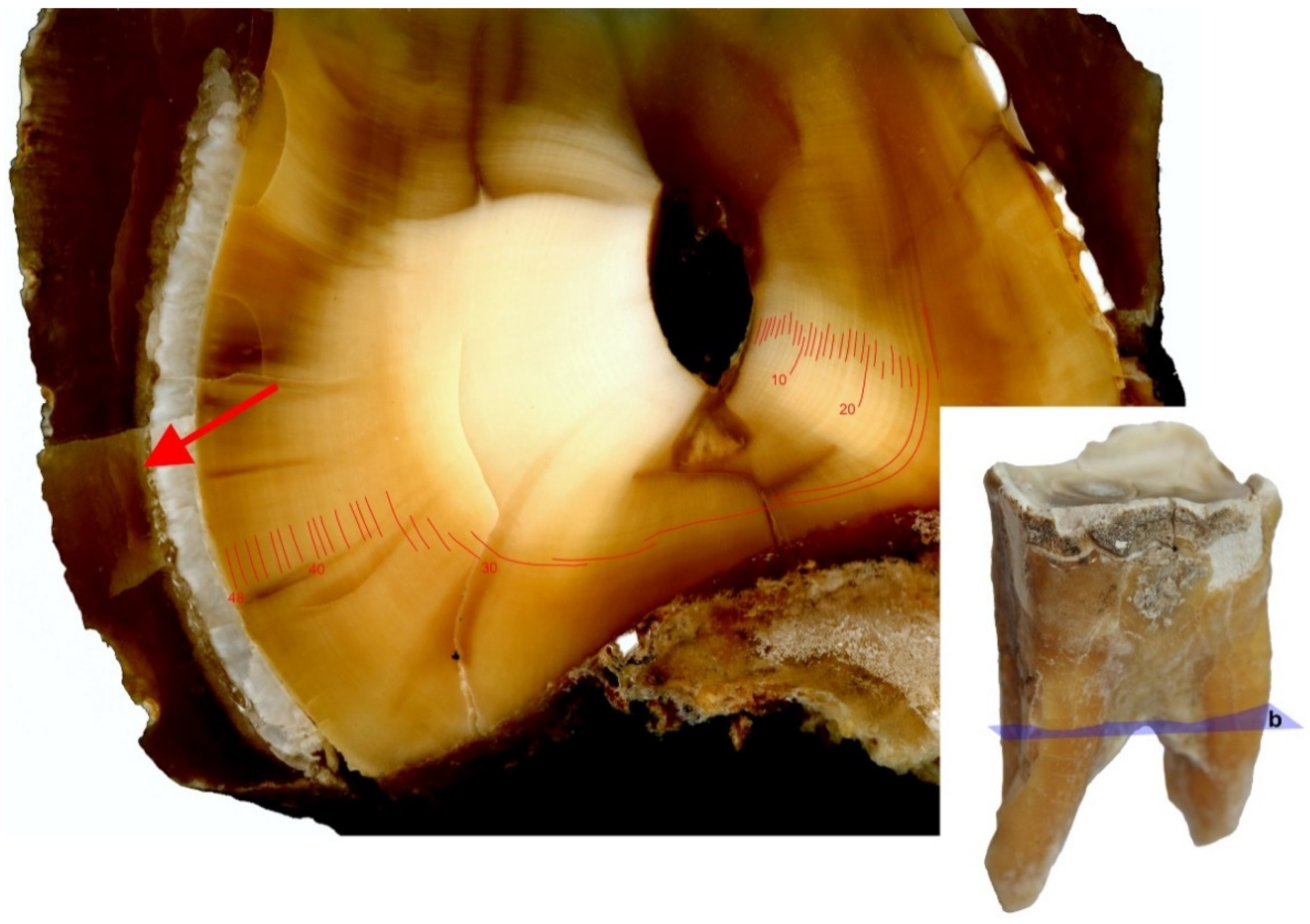
2.4. Annual Season Analysis Based on Cement Lines
3. Results
3.1. Analysis of Tooth Sections
3.2. Analysis of Seasonality Based on Cement Lines
3.3. X-ray Imaging of the Mandible
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Reitz, E.; Wing, E. Zooarchaeology, 2nd ed.; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Gifford-Gonzalez, D. An Introduction to Zooarchaeology; Springer International Publishing: New York, NY, USA, 2018; p. 604. [Google Scholar]
- Foote, M.; Miller, A.I.; Raup, D.M.; Stanley, S.M. Principles of Paleontology; W.H. Freeman: New York, NY, USA, 2007. [Google Scholar]
- Bartosiewicz, L.; Gal, E. Pathological lesions in working animals. In Shuffling Nags, Lame Ducks. The Archaeology of Animals Disease; Oxbow Books: Oxford, UK, 2013; pp. 130–154. [Google Scholar]
- Bartosiewicz, L.; Gal, E. Care or Neglect? Evidence of Animal Disease in Archaeology. In Proceedings of the 6th Meeting of the Animal Palaeopathology Working Group of the International Council for Archaeozoology (ICAZ), Budapest, Hungary, 26–29 May 2016; Oxbow Books: Oxford, UK, 2018; p. 288. [Google Scholar]
- Lasota-Moskalewska, A. Archeozoologia, Ssaki; Wydawnictwa Uniwersytetu Warszawskiego: Warsaw, Poland, 2008; p. 321. [Google Scholar]
- Pasicka, E.; Onar, V.; Dixon, P.M. Supernumerary cheek tooth in a Byzantine horse from Theodosius Harbour, Istanbul, Turkey. Equine Vet. Educ. 2015, 29, 266–269. [Google Scholar] [CrossRef] [Green Version]
- Nowakowski, D. Frequency of appearance of transverse (Harris) lines reflects living conditions of the Pleistocene bear—Ursus ingressus—(Sudety Mts., Poland). PLoS ONE 2018, 13, e0196342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowakowski, D.; Stefaniak, K. Pathological Changes of the Cranium of a Young Female Cave Bear (Ursus spelaeus R.)-A Case Study (the Sudety Mts, Poland): Pathological Changes of a Cave Bear Cranium. Int. J. Osteoarchaeol. 2015, 25, 119–125. [Google Scholar] [CrossRef]
- Janeczek, M.; Chrószcz, A.; Henklewski, R.; Czerski, A.; Onar, V.; Żygadło, L.; Krupska, A.; Pasicka, E. Vertebral disease in excavated canine in Lower Silesia, Poland. Int. J. Paleopathol. 2015, 10, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Chrószcz, A.; Janeczek, M.; Pasicka, E.; Klećkowska-Nawrot, J. Height at the withers estimation in the horses based on the internal dimension of cranial cavity. Folia Morphol. 2014, 73, 143–148. [Google Scholar] [CrossRef] [Green Version]
- Onar, V.; Pazvant, G.; Pasicka, E.; Armutak, A.; Alpak, H. Byzantine horse skeletons of Theodosius harbour: 2. Withers height estimation. Rev. Med. Vet. 2015, 166, 30–42. [Google Scholar]
- Pawłowska, K.; Stefaniak, K.; Nowakowski, D. Healed antler fracture from a giant deer (Megaloceros giganteus) from the Pleistocene in Poland. Palaeontol. Electron. 2014, 17, 1–9. [Google Scholar] [CrossRef]
- Pasicka, E.; Chrószcz, A.; Janeczek, M.; Mucha, A. Craniometric analysis of Early Medieval horses Equus przewalskii f. caballus (Linnaeus, 1758) from chosen areas in Poland. Turk. J. Vet. Anim. Sci. 2012, 36, 688–697. [Google Scholar]
- Renfrew, C.; Bahn, P. Archeologia.Teorie, Metody, Praktyka; Prószyński i S-ka: Warsaw, Poland, 2002. [Google Scholar]
- Morris, P. The Use of Teeth for Estimating the Age of Wild Mammals. In Development, Function and Evolution of Teeth; Butler, P.M., Joysey, K.A., Eds.; London Academic Press: London, UK, 1978; pp. 483–494. [Google Scholar]
- Matson, G.M. Workbook for Cementum Analysis; Matson’s Laboratory: Milltown, MT, USA, 1981. [Google Scholar]
- Abelova, M. Dental cementum of bears (Ursidae)—Possibilities of microscopic study. Neue Forschungenzum Höhlenbären in Europa. Nat. Ges. Nürnberg. Abh. Band. 2005, 45, 11–18. [Google Scholar]
- Veitschegger, K.; Kolb, C.; Amson, E.; Sánchez-Villagra, M.R. Longevity and life history of cave bears—A review and novel data from tooth cementum and relative emergence of permanent dentition. Hist. Biol. 2018, 31, 510–516. [Google Scholar] [CrossRef]
- Beasley, M.J.; Brown, W.A.B.; Legge, A.J. Incremental banding in dental cementum: Methods of preparation of teeth from archeological sites and for modern comparative specimens. Int. J. Osteoarchaeol. 1992, 2, 37–50. [Google Scholar] [CrossRef]
- Burke, A.M. Observation of incremental growth structures in dental cementum using the scanning electron microscope. Ar-chaeozoologia 1993, 5, 41–54. [Google Scholar]
- Debeljak, I. A simple preparation technique of cave bear teeth for age determination by cementum increments. Rev. Paleobiol. 1966, 15, 105–108. [Google Scholar]
- Debeljak, I. Dental cementum in the cave bear; comparison of different preparation techniques. Geol. Zb. 2000, 15, 25–40. [Google Scholar]
- Fancy, S.G. Preparation of Mammalia for the age determination by cementum layers: A review. Wild. Soc. Bull. 1980, 8, 242–248. [Google Scholar]
- Stallibrass, S. The use of cement layers for absolute ageing of mammalian teeth: A selective review of the literature, with suggestions for future research and alternative applications. In Ageing and Sexing Animal Bones from Archaeological Sites; Wilson, B., Grigson, C., Payne, S., Eds.; Taylor Francis: Abingdon, UK, 1982; Volume 109, pp. 109–126. [Google Scholar]
- Waugh, D.A.; Suydam, R.S.; Ortiz, J.D.; Thewissen, J.G.M. Validation of Growth Layer Group (GLG) depositional rate using daily incremental growth lines in the dentin of beluga (Delphinapterus leucas (Pallas, 1776)) teeth. PLoS ONE 2018, 13, e0190498. [Google Scholar] [CrossRef]
- Ruscillo, D. Zooarchaeology: Methods of Collecting Age and Sex Data. In Encyclopedia of Global Archaeology; Smith, C., Ed.; Springer New York: New York, NY, USA, 2014; pp. 8000–8010. [Google Scholar]
- Silberman, M.S.; Fulton, R.B. Medical problems of captive and wild rhinoceros—A review of the literature and personal experiences. J. Zoo An. Med. 1979, 10, 6–16. [Google Scholar] [CrossRef]
- Van der Made, J. The rhinos from the Middle Pleistoceneof Neumark-Nord (Saxony-Anhalt). Veröffentlichungen Landesamtes Denkmalpfl. Archäologie 2010, 62, 433–500. [Google Scholar]
- Deng, T.; Wang, X.; Fortelius, M.; Li, Q.; Wang, Y.; Tseng, Z.J.; Takeuchi, G.T.; Saylor, J.E.; Säilä, L.K.; Xie, G. Out of Tibet: Pliocene Woolly Rhino Suggests High-Plateau Origin of Ice Age Megaherbivores. Science 2011, 333, 1285–1288. [Google Scholar] [CrossRef]
- Pandolfi, L. Evolutionary history of Rhinocerotina (Mammalia, Perissodactyla). Foss. Rep. Palaeontol. 2018, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Stefaniak, K.; Stachowicz-Rybka, R.; Borówka, R.K.; Hrynowiecka, A.; Sobczyk, A.; Moskal-del Hoyo, M.; Kotowski, A.; Nowakowski, D.; Krajcarz, M.T.; Billia, E.M.E.; et al. Browsers, grazers or mix-feeders? Study of the diet of extinct Pleistocene Eurasian forest rhinoceros Stephanorhinus kirchbergensis (Jäger, 1839) and woolly rhinoceros Coelodonta antiquitatis (Blumenbach, 1799). Quat. Int. 2020. [Google Scholar] [CrossRef]
- Brehm, A. Życie Zwierząt. Ssaki; Państwowe Wydawnictwo Naukowe: Warsaw, Poland, 1963. [Google Scholar]
- Groves, C.P.; Fernando, P.; Robovský, J. The Sixth Rhino: A Taxonomic Re-Assessment of the Critically Endangered Northern White Rhinoceros. PLoS ONE 2010, 5, e9703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, L.A.; Müller, D.W.; Schwitzer, C.; Kaiser, T.M.; Codron, D.; Schulz, E.; Clauss, M. Tooth wear in captive rhinoceroses (Diceros, Rhinoceros, Ceratotherium: Perissodactyla) differs from that of free-ranging conspecifics. Contrib. Zool. 2014, 83, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Fortelius, M. Ungulate cheek teeth: Developmental, functional, and evoutionary interrelations. Acta Zool. Fenn. 1985, 180, 1–76. [Google Scholar]
- Prat, F. Podstawy Oznaczania Szczątków Ważniejszych Ssaków Plejstoceńskich; Student Textbooks No.554. Text; Uniwersytet Jagieloński: Cracow, Poland, 1987; pp. 2–57. [Google Scholar]
- Antoine, P.-O.; Downing, K.F.; Crochet, J.-Y.; Duranthon, F.; Flynn, L.J.; Marivaux, L.; Métais, G.; Rajpar, A.R.; Roohi, G. A revision of Aceratherium blanfordi Lydekker, 1884 (Mammalia: Rhinocerotidae) from the Early Miocene of Pakistan: Postcranials as a key. Zool. J. Linn. Soc. 2010, 160, 139–194. [Google Scholar] [CrossRef] [Green Version]
- Böhmer, C.; Rössner, G.E. Dental paleopathology in fossil rhinoceroses: Etiology and implications. J. Zool. 2017, 304, 3–12. [Google Scholar] [CrossRef]
- Konig, H.E.; Liebich, H.-G. Veterinary Anatomy of Domestic Mammals. Textbook and Colour Atlas, 1st ed.; Schattauer: Stuttgart, Germany, 2012. [Google Scholar]
- Hillson, S. Teeth, 2nd ed.; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Dietz, O.; Huskamp, B. Praktyka kliniczna: Konie; Galaktyka, Sp. z.o.o: Łódź, Poland, 2005. [Google Scholar]
- Rensberger, J.M.; Koenigswald, W.V. Functional and phylogenetic interpretation of enamel microstructure in rhinoceroses. Paleobiology 1980, 6, 477–495. [Google Scholar] [CrossRef]
- Tafforeau, P.; Bentaleb, I.; Jaeger, J.J.; Martin, C. Nature of laminations and mineralization in rhinoceros enamel usinghistology and X-ray synchrotron microtomography: Potential implications for palaeoenvironmental isotopic studies. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007, 246, 206–227. [Google Scholar] [CrossRef]
- Leiss-Holzinger, E.; Wiesauer, K.; Stephani, H.; Heise, B.; Stifter, D.; Kriechbaumer, B.; Spachinger, S.J.; Gusenbauer, C.; Withalm, G. Imaging of the inner structure of cave bear teeth by novel non-destructive techniques. Palaeontol. Electron. 2015, 18, 1–15. [Google Scholar] [CrossRef]
- Smith, K.G.; Strother, K.A.; Rose, J.C.; Savelle, J.M. Chemical Ultrastructure of Cementum Growth-Layers of Teeth of Black Bears. J. Mammal. 1994, 75, 406–409. [Google Scholar] [CrossRef]
- Kirillova, I.V.; Chernova, O.F.; Van Der Made, J.; Kukarskih, V.V.; Shapiro, B.; Van Der Plicht, J.; Shidlovskiy, F.K.; Heintzman, P.D.; Van Kolfschoten, T.; Zanina, O.G. Discovery of the skull of Stephanorhinus kirchbergensis (Jäger, 1839) above the Arctic Circle. Quat. Res. 2017, 88, 537–550. [Google Scholar] [CrossRef] [Green Version]
- Hillman-Smith, A.K.K.; Owen-Smith, N.; Anderson, J.L.; Hall-Martin, A.J.; Selaladi, J.P. Age estimation of the White rhinoceros (Ceratotherium simum). J. Zool. 1986, 210, 355–377. [Google Scholar] [CrossRef]
- Burke, A.; Castanet, J. Histological Observations of Cementum Growth in Horse Teeth and their Application to Archaeology. J. Archaeol. Sci. 1995, 22, 479–493. [Google Scholar] [CrossRef]
- Sten, S.A. A method of age determination on archaeological and modern cattle (Bos taurus) by counting of tooth annuli in the cementum. Laborativ Arkeol. 1989, 5, 195–199. [Google Scholar]
- Haynes, G. Tooth Wear Rate in Northern Bison. J. Mammal. 1984, 65, 487–491. [Google Scholar] [CrossRef]
- Rudge, M.R. Ageing domestic sheep (Ovis aries L.) from growth lines in the cementum of the first incisor. New Zealand J. Zool. 1976, 3, 421–424. [Google Scholar] [CrossRef]
- Debeljak, I. Ontogenetic development of dentition in the cave bear. Geologija 1996, 39, 13–77. [Google Scholar] [CrossRef]
- Mitchell, B. Growth Layers in Dental Cement for Determining the Age of Red Deer (Cervus elaphus L.). J. Anim. Ecol. 1967, 36, 279. [Google Scholar] [CrossRef]
- Bromage, T.G. Enamel incremental periodicity in the pig-tailed macaque: A polychrome fluorescent labeling study of dental hard tissues. Am. J. Phys. Anthr. 1991, 86, 205–214. [Google Scholar] [CrossRef]
- Schour, I.; Hoffman, M.M. Studies in Tooth Development: I. The 16 Microns Calcification Rhythm in the Enamel and Dentin from Fish to Man. J. Dent. Res. 1939, 18, 91–102. [Google Scholar] [CrossRef]
- Schour, I.; Hoffman, M.M. Studies in Tooth Development: II. The rate of apposition of enamel and dentin in man and other mammals. J. Dent. Res. 1939, 18, 161–175. [Google Scholar] [CrossRef]
- Rosenberg, G.D.; Simmons, D.J. Rhythmic dentinogenesis in the rabbit incisor: Circadian, ultradian, and infradian periods. Calcif. Tissue Int. 1980, 32, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, S.; Newman, H.; Poole, D. Diurnal periodicity of von Ebner growth lines in pig dentine. Arch. Oral Biol. 1977, 22, 511–513. [Google Scholar] [CrossRef]
- Chhem, R.K.; Brothwell, D.R. Paleoradiology—Imaging Mummies and Fossils; Springer-Verlag Berlin Heidelberg: Heidelberg, Germany, 2008; p. 162. [Google Scholar]














Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pasicka, E.; Nowakowski, D.; Bendrey, R.; Melnyk, O.P. A Model for Dental Age Verification Using Ultrastructural Imaging for Modern and Fossil Representatives of the Rhinocerotidae Family. Animals 2021, 11, 910. https://doi.org/10.3390/ani11030910
Pasicka E, Nowakowski D, Bendrey R, Melnyk OP. A Model for Dental Age Verification Using Ultrastructural Imaging for Modern and Fossil Representatives of the Rhinocerotidae Family. Animals. 2021; 11(3):910. https://doi.org/10.3390/ani11030910
Chicago/Turabian StylePasicka, Edyta, Dariusz Nowakowski, Robin Bendrey, and Oleg P. Melnyk. 2021. "A Model for Dental Age Verification Using Ultrastructural Imaging for Modern and Fossil Representatives of the Rhinocerotidae Family" Animals 11, no. 3: 910. https://doi.org/10.3390/ani11030910