
Characterization and Comparative Transcriptomic Analysis of Skeletal Muscle in Pekin Duck at Different Growth Stages Using RNA-Seq



Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Ethics Approval and Consent to Participate
2.2. Animal and Tissue Collection
2.3. RNA Isolation, Library Preparation and Sequencing
2.4. Quality Control and Comparative Analysis
2.5. SNP/InDel Analysis and Prediction of AS
2.6. New Genes Analysis
2.7. Quantification and Differential Expression Analysis
2.8. Analysis of GO Enrichment and KEGG Pathway Enrichment
2.9. Protein–Protein Interaction Network
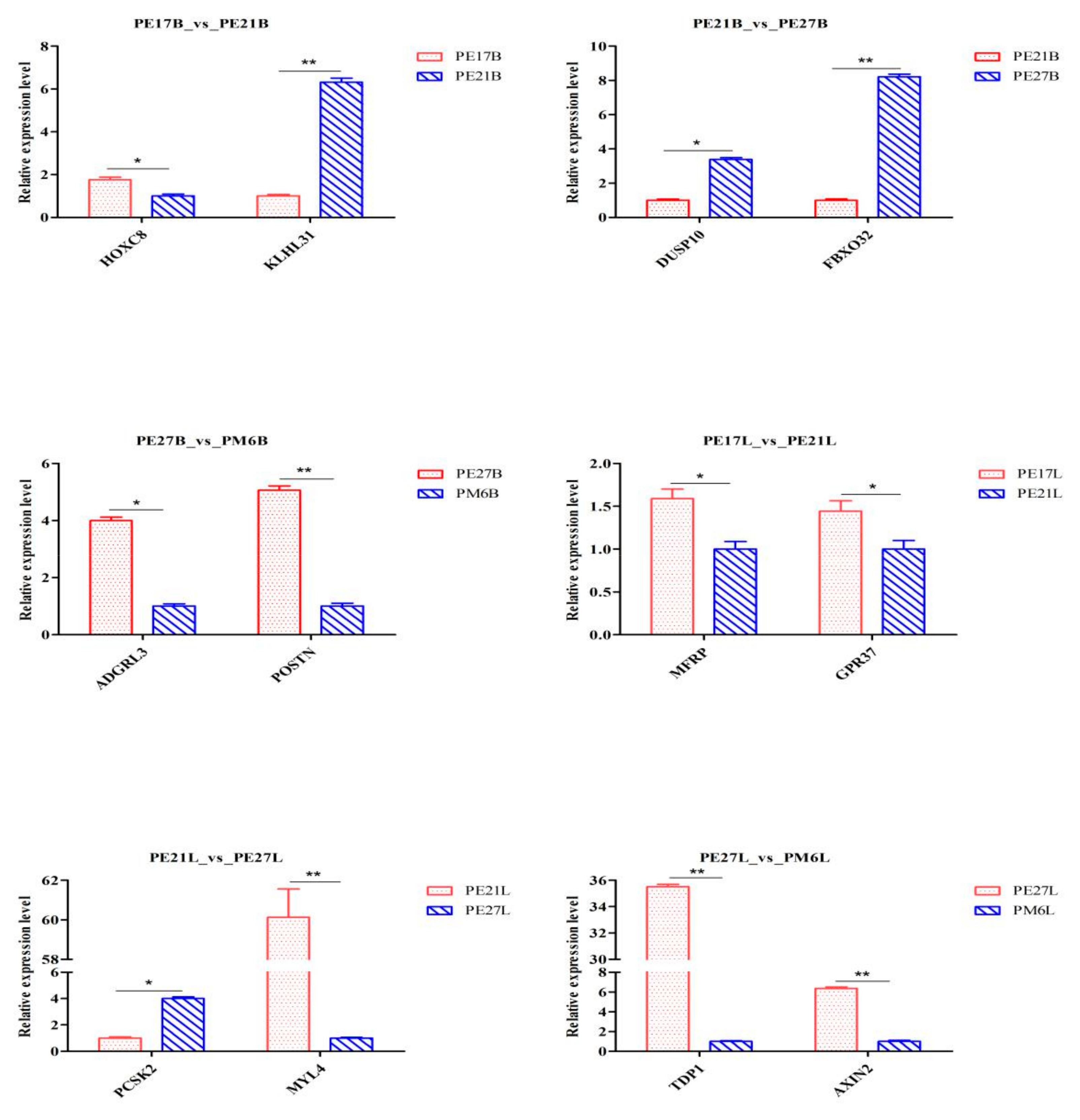
2.10. Verification of Results by qPCR
3. Results
3.1. Evaluation of Transcriptome Profiles
3.2. Annotation and Classification of SNP/InDel
3.3. Prediction of Alternative Splicing
3.4. New Genes Analysis
3.5. Gene Functional Annotation and Classification
3.6. Analysis of Differentially Expressed Genes
3.7. GO Annotation and KEGG Pathway Analysis
3.8. Analysis of Protein–Protein Interaction Network
3.9. Validation of RNA-Seq Results
4. Discussion
4.1. Analysis Summary of Sequencing Data
4.2. Annotation and Classification of SNP/InDel in Skeletal Muscle Developmental Process
4.3. Alternative Splicing Events in Skeletal Muscle Developmental Process
4.4. DEGs Analyzed at All Time Points
4.5. Analysis of GO and KEGG Pathway
4.6. Analysis of Protein–Protein Interaction Network
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Apponi, L.H.; Corbett, A.H.; Pavlath, G.K. RNA-binding proteins and gene regulation in myogenesis. Trends Pharmacol. Sci. 2011, 32, 652–658. [Google Scholar] [CrossRef] [Green Version]
- Buckingham, M.; Rigby, P.J. Gene regulatory networks and transcriptional mechanisms that control myogenesis. Dev. Cell. 2014, 28, 225–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aiello, D.; Patel, K.; Lasagna, E. The myostatin gene: An overview of mechanisms of action and its relevance ti livestock animals. Anim. Genet. 2018, 49, 505–519. [Google Scholar] [CrossRef] [Green Version]
- Abmayr, S.M.; Pavlath, G.K. Myoblast fusion: Lessons from flies and mice. Development 2012, 139, 641–656. [Google Scholar] [CrossRef] [Green Version]
- Scaal, M.; Marcelle, C. Chick muscle development. Int. J. Dev. Biol. 2018, 62, 127–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, J.T.; Li, H.F.; Shan, Y.J.; Xu, W.J.; Chen, W.F.; Song, C.; Song, W.T. Expression profile of IGF-I-calcineurin-NFATc3-dependent pathway genes in skeletal muscle during early development between duck breeds differing in growth rates. Dev. Genes Evol. 2015, 225, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Ba, Y.; Zhuang, Q.F.; Zhong, G.F. RNA-Seq technology and its application in fish transcriptomics. OMICS 2014, 18, 98–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garber, M.; Grabherr, M.G.; Guttman, M.; Trapnell, C. Computational methods for transcriptome annotation and quantification using RNA-seq. Nat. Methods 2011, 8, 469–477. [Google Scholar] [CrossRef] [Green Version]
- Zheng, C.L.; Kawane, S.; Bottomly, D.; Wilmot, B. Analysis considerations for utilizing RNA-seq to characterize the brain transcriptome. Int. Rev. Neurobiol. 2014, 116, 21–54. [Google Scholar] [PubMed]
- Calviello, L.; Ohler, U. Beyond read-counts: Ribo-seq data analysis to understand the functions of the transcriptome. Trends Genet. 2017, 33, 728–744. [Google Scholar] [CrossRef]
- Roux, J.; Rosikiewicz, M.; Robinson-Rechavi, M. What to compare and how: Comparative transcriptomics for Evo-Devo. J. Exp. Zool. B Mol. Dev. Evol. 2015, 324, 372–382. [Google Scholar] [CrossRef] [Green Version]
- Xue, Q.; Zhang, G.X.; Li, T.T.; Ling, J.J.; Zhang, X.Q.; Wang, J.Y. Transcriptomic profile of leg muscle during early growth in chicken. PLoS ONE 2017, 12, e0173824. [Google Scholar] [CrossRef]
- Zhang, Z.R.; Du, H.R.; Yang, C.W.; Li, Q.Y.; Qiu, M.H.; Song, X.Y.; Yu, C.L.; Jiang, X.Y.; Liu, L.; Hu, C.M.; et al. Comparative transcriptome analysis reveals regulators mediating breast muscle growth and development in three chicken breeds. Anim. Biotechnol. 2019, 30, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.X.; Hu, Y.; Ji, G.G.; Li, H.F. Rapid-Sexing Poultries via a New Pair of Universal Primers. J. Agric. Biotechnol. 2014, 22, 1567–1574. (In Chinese) [Google Scholar]
- Humble, E.; Thorne, M.A.S.; Forcada, J.; Hoffman, J.I. Transcriptomic SNP discovery for custom genotyping arrays: Impacts of sequence data, SNP calling method and genotyping technology on the probability of validation success. BMC Res. Notes 2016, 9, 418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piskol, R.; Ramaswami, G.; Li, J.B. Reliable identification of genomic variants from RNA-seq data. Am. J. Hum. Genet. 2013, 93, 641–651. [Google Scholar] [CrossRef] [Green Version]
- Florea, L.; Song, L.; Salzberg, S.L. Thousands of exon skipping events differentiate among splicing patterns in sixteen human tissues. F1000Res 2013, 2, 188. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.L.; Liu, H.L.; Cheng, Y.S. A new DNA marker of the TMIGD1 gene used to identify high fertilization rates in Tsaiya ducks (Anas platyrhynchos). J. Reprod. Dev. 2018, 64, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Sultana, H.; Seo, D.; Choi, N.R.; Bhuiyan, M.S.A.; Lee, S.H.; Heo, K.N.; Lee, J.H. Identification of polymorphisms in MITF and DCT genes and their associations with plumage colors in Asian duck breeds. Asian-Aust. J. Anim. Sci. 2018, 31, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Lee, E.A.; Kim, J.M.; Lim, K.S.; Ryu, Y.C.; Jeon, W.M.; Hong, K.C. Effects of variation in porcine MYOD1 gene on muscle fiber characteristics, lean meat production, and meat quality traits. Meat Sci. 2012, 92, 36–43. [Google Scholar] [CrossRef]
- Wu, Y.; Pan, A.L.; Pi, J.S.; Pu, Y.J.; Du, J.P.; Liang, Z.H.; Shen, J. One novel SNP of growth hormone gene and its associations with growth and carcass traits in ducks. Mol. Biol. Rep. 2012, 39, 8027–8033. [Google Scholar] [CrossRef]
- Djari, A.; Esquerré, D.; Weiss, B.; Martins, F.; Meersseman, C.; Boussaha, M.; Klopp, C.; Rocha, D. Gene-based single nucleotide polymorphism discovery in bovine muscle using next-generation transcriptomic sequencing. BMC Genomics 2013, 14, 307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piórkowska, K.; Żukowski, K.; Ropka-Molik, K.; Tyra, M. Detection of genetic variants between different Polish Landrace and Puławska pigs by means of RNA-seq analysis. Anim. Genet. 2018, 49, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Blanca, J.; Esteras, C.; Ziarsolo, P.; Perez, D.; Fernandez-Pedrosa, V.; Collado, C.; de Pablos, G.R.R.; Ballester, A.; Roig, C.; Cañizares, J.; et al. Transcriptome sequencing for SNP discovery across Cucumis melo. BMC Genomics 2012, 13, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell. Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Clevenger, J.; Chavarro, C.; Pearl, S.A.; Ozias-Akins, P.; Jackson, S.A. Single nucleotide polymorphism identification in polyploids: A review, example, and recommendations. Mol. Plant. 2015, 8, 831–846. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.X.; Xu, Y.O.; Lin, Y.Q. Transcriptome analyses reveal genes of alternative splicing associated with muscle development in chickens. Gene 2018, 676, 146–155. [Google Scholar] [CrossRef]
- Li, B.J.; Zhu, Z.X.; Qin, H.; Meng, Z.N.; Lin, H.R.; Xia, J.H. Genome-wide characterization of alternative splicing events and their responses to cold stress in tilapia. Front. Genet. 2020, 11, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, B.; Jin, J.P. Troponin T isoforms and posttranscriptional modifications: Evolution, regulation and function. Arch. Biochem. Biophys. 2011, 505, 144–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sebastian, S.; Faralli, H.; Yao, Z.Z.; Rakopoulos, P.; Palii, C.; Cao, Y.; Singh, K.; Liu, Q.C.; Chu, A.; Aziz, A.; et al. Tissue-specific splicing of a ubiquitously expressed transcription factor is essential for muscle differentiation. Gene Dev. 2013, 27, 1247–1259. [Google Scholar] [CrossRef] [Green Version]
- Spletter, M.L.; Schnorrer, F. Transcriptional regulation and alternative splicing cooperate in muscle fiber-type specification in flies and mammals. Exp. Cell. Res. 2014, 321, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.R.; Suh, Y.; Shin, S.; Woodfint, R.M.; Hwang, S.; Lee, K. Exogenous expression of an alternative splicing variant of myostatin prompts leg muscle fiber hyperplasia in Japanese quail. Int. J. Mol. Sci. 2019, 20, 4617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.L.; Wang, G.Z.; Wang, J.M.; Ji, Z.B.; Liu, Z.H.; Pi, X.S.; Chen, C.X. Characterization and comparative analyses of muscle transcriptomes in dorper and small-tailed han sheep using RNA-seq technique. PLoS ONE 2013, 30, e72686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.J.; Xi, Y.; Liu, G.R.; Zhao, Y.Q.; Li, J.; Lei, M.G. Comparative transcriptomic analysis of skeletal muscle tissue during prenatal stages in Tongcheng and Yorkshire pig using RNA-seq. Funct. Integr. Genomics 2018, 18, 195–209. [Google Scholar] [CrossRef]
- Hao, Y.; Feng, Y.; Yang, P.; Cui, Y.; Liu, J.; Yang, C.; Gu, X. Transcriptome analysis reveals that constant heat stress modifies the metabolism and structure of the porcine longissimus dorsi skeletal muscle. Mol. Gen. Genomics 2016, 291, 2101–2115. [Google Scholar] [CrossRef]
- Leppek, K.; Das, R.; Barna, M. Functional 5′ UTR mRNA structures in eukaryotic translation regulation and how to find them. Nat. Rev. Mol. Cell Biol. 2018, 19, 158–174. [Google Scholar] [CrossRef]
- Schiaffino, S.; Rossi, A.C.; Smerdu, V.S.; Leinwand, L.A.; Reggiani, C. Developmental myosins: Expression patterns and functional significance. Skelet Muscle 2015, 5, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.F.; Pan, Z.X.; Zhang, K.; Yu, M.L.; Yu, D.B.; Lu, Y.L.; Wang, J.T.; Zhang, J.; Zhang, K.N.; Du, W.X. Identification of the differentially expressed genes of muscle growth and intramuscular fat metabolism in the development stage of yellow broilers. Genes 2020, 11, 244. [Google Scholar] [CrossRef] [Green Version]
- Ye, M.S.; Ye, F.; He, L.T.; Luo, B.; Yang, F.L.; Cui, C.; Zhao, X.L.; Yin, H.D.; Li, D.Y.; Xu, H.Y.; et al. Transcriptomic analysis of chicken Myozenin 3 regulation reveals its potential role in cell proliferation. PLoS ONE 2017, 12, e0189476. [Google Scholar] [CrossRef] [Green Version]
- Ciecierska, A.; Motyl, T.; Sadkowski, T. Transcriptomic profile of primary culture of skeletal muscle cells isolated from semitendinosus muscle of beef and dairy bulls. Int. J. Mol. Sci. 2020, 21, 4794. [Google Scholar] [CrossRef]
- Huang, X.W.; Zhang, H.; Guo, X.R.; Zhu, Z.X.; Cai, H.B.; Kong, X.Y. Insulin-like growth factor 2 mRNA-binding protein 1 (IGF2BP1) in cancer. J. Hematol. Oncol. 2018, 11, 88. [Google Scholar] [CrossRef] [Green Version]
- Stöhr, N.; Köhn, M.; Lederer, M.; Glass, M.; Reinke, C.; Singer, R.H.; Hüttelmaier, S. IGF2BP1 promotes cell migration by regulating MK5 and PTEN signaling. Genes Dev. 2012, 26, 176–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahaira, L.G.; Katsara, O.; Pappou, E.; Iliopoulou, E.G.; Fortis, S.; Antsaklis, A.; Fotinopoulos, P.; Baxevanis, C.N.; Papamichail, M.; Perez, S.A. IGF2BP1 expression in human mesenchymal stem cells significantly affects their proliferation and is under the epigenetic control of TET1/2 Demethylases. Stem Cell Dev. 2014, 23, 2501–2512. [Google Scholar] [CrossRef]
- Huang, H.L.; Weng, H.Y.; Sun, W.S.; Qin, X.; Shi, H.L.; Wu, H.Z.; Zhao, B.X.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N6-methyladenosine by IGF2BP Proteins Enhances mRNA Stability and Translation. Nat. Cell Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef]
- Liu, Z.J.; Wu, G.Y.; Lin, C.; Guo, H.L.; Xu, J.J.; Zhao, T.L. IGF2BP1 over-expression in skin squamous cell carcinoma cells is essential for cell growth. Biochem. Biophys. Res. Commun. 2018, 501, 731–738. [Google Scholar] [CrossRef]
- Hansen, T.V.O.; Hammer, N.A.; Nielsen, J.; Madsen, M.; Dalbaeck, C.; Wewer, U.M.; Christiansen, J.; Nielsen, F.C. Dwarfism and impaired gut development in insulin-like growth factor II mRNA-binding protein 1-deficient mice. Mol. Cell Biol. 2004, 24, 4448–4464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arber, S.; Caroni, P. Specificity of single LIM motifs in targeting and LIM/LIM interactions in situ. Genes Dev. 1996, 10, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.H.; Zhao, Y. The diverse biofunctions of LIM domain proteins: Determined by subcellular localization and protein-protein interaction. Biol. Cell. 2007, 99, 489–502. [Google Scholar] [CrossRef]
- Vafiadaki, E.; Arvanitis, D.A.; Sanoudou, D. Muscle Lim Protein: Master regulator of cardiac and skeletal muscle function. Gene 2015, 566, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Arber, S.; Halder, G.; Caroni, P. Muscle LIM protein, a novel essential regulator of myogenesis, promotes myogenic differentiation. Cell 1994, 79, 221–231. [Google Scholar] [CrossRef]
- Kong, Y.; Flick, M.J.; Kudla, A.J.; Konieczny, S.F. LIM protein promotes myogenesis by enhancing the activity of MyoD. Mol. Cell. Biol. 1997, 17, 4750–4760. [Google Scholar] [CrossRef] [Green Version]
- Rashid, M.M.; Runci, A.; Russo, M.A.; Tafani, M. Muscle Lim Protein (MLP)/CSRP3 at the crossroad between mechanotransduction and autophagy. Cell. Death. Dis. 2015, 6, e1940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, C.; Han, S.S.; Tang, S.Y.; He, H.R.; Shen, X.X.; Zhao, J.; Chen, Y.Q.; Wei, Y.H.; Wang, Y.; Zhu, Q.; et al. The autophagy regulatory molecule CSRP3 interacts with LC3 and protects against muscular dystrophy. Int. J. Mol. Sci. 2020, 21, 749. [Google Scholar] [CrossRef] [Green Version]
- Lim, W.; Jeong, W.; Kim, J.; Ka, H.; Bazer, F.W.; Han, J.Y.; Song, G. Differential expression of secreted phosphoprotein 1 in response to estradiol-17β and in ovarian tumors in chickens. Biochem. Biophys. Res. Commun. 2012, 422, 494–500. [Google Scholar] [CrossRef]
- La, Y.F.; Zhang, X.X.; Li, F.D.; Zhang, D.Y.; Li, C.; Mo, F.T.; Wang, W.M. Molecular characterization and expression of SPP1, LAP3 and LCORL and their association with growth traits in sheep. Genes 2019, 10, 616. [Google Scholar] [CrossRef] [Green Version]
- Uaesoontrachoon, K.; Yoo, H.J.; Tudor, E.M.; Pike, R.N.; Mackie, E.J.; Pagel, C.N. Osteopontin and skeletal muscle myoblasts: Association with muscle regeneration and regulation of myoblast function in vitro. Int. J. Biochem. Cell. Biol. 2008, 40, 2303–2314. [Google Scholar] [CrossRef] [PubMed]
- Nghiem, P.P.; Kornegay, J.N.; Uaesoontrachoon, K.; Bello, L.; Yin, Y.; Kesari, A.; Mittal, P.; Schatzberg, S.J.; Many, G.M.; Lee, N.H.; et al. OPN may interact with AKT1/ MSTN/FoxO1 to modify normal and dystrophic muscle. Muscle Nerve 2017, 56, 1119–1127. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, H.; Kohara, R.; Sugi, M.; Usui, A.; Oyama, K.; Mannen, H.; Sasazaki, S. The non-synonymous mutation in bovine SPP1 gene influences carcass weight. Heliyon 2019, 51, e03006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenberg, C.C.; Connelly, P.S.; Daniels, M.P.; Horowits, R. Krp1 (Sarcosin) promotes lateral fusion of myofibril assembly intermediates in cultured mouse cardiomyocytes. Exp. Cell. Res. 2008, 314, 1177–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, W.S.; Li, Y.Q.; Zhou, X.J.; Deng, Y.; Wang, Z.Q.; Yuan, W.Z.; Li, D.L.; Zhu, C.B.; Zhao, X.Y.; Mo, X.Y.; et al. A novel human BTB-kelch protein KLHL31, strongly expressed in muscle and heart, inhibits transcriptional activities of TRE and SRE. Mol. Cell. 2008, 26, 443–453. [Google Scholar]
- Papizan, J.B.; Garry, G.A.; Brezprozvannaya, S.; McAnally, J.R.; Bassel-Duby, R.; Liu, N.; Olson, E.N. Deficiency in Kelch protein Klhl31 causes congenital myopathy in mice. J. Clin. Investig. 2017, 127, 3730–3740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abou-Elhamd, A.; Cooper, O.; Münsterberg, A. Klhl31 is associated with skeletal myogenesis and its expression is regulated by myogenic signals and Myf-5. Mech. Dev. 2009, 126, 852–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abou-Elhamd, A.; Alrefaei, A.F.; Mok, G.F.; Garcia-Morales, C.; Abu-Elmagd, M.; Wheeler, G.N.; Münsterberg, A.E. Klhl31 attenuates β-catenin dependent Wnt signaling and regulates embryo myogenesis. Dev. Biol. 2015, 402, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Onteru, S.K.; Gorbach, D.M.; Young, J.M.; Garrick, D.J.; Dekkers, J.C.M.; Rothschild, M.F. Whole Genome Association Studies of Residual Feed Intake and Related Traits in the Pig. PLoS ONE 2013, 8, e61756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hargreaves, M.; Spriet, L.L. Exercise metabolism: Fuels for the fire. Cold Spring Harb. Perspect. Med. 2018, 8, a029744. [Google Scholar] [CrossRef]
- Hawley, J.A.; Hargreaves, M.; Joyner, M.J.; Zierath, J.R. Integrative biology of exercise. Cell 2014, 159, 738–749. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Kim, S.H.; Lee, J.S.; Yang, Y.H.; Nam, J.M.; Kim, B.W.; Ko, Y.G. Mitochondrial oxidative phosphorylation complexes exist in the sarcolemma of skeletal muscle. BMB Rep. 2016, 49, 116–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freedman, B.R.; Rodriguez, A.B.; Leiphart, R.J.; Newton, J.B.; Ban, E.; Sarver, J.J.; Mauck, R.L.; Shenoy, V.B.; Soslowsky, L.J. Dynamic loading and tendon healing affect multiscale tendon properties and ECM stress transmission. Sci. Rep. 2018, 8, 10854. [Google Scholar] [CrossRef]
- Ridley, A.J.; Hall, A. The small GTP-binding protein rho regulates the assembly of focal adhesions and actin stress fibers in response to growth factors. Cell 1992, 70, 389–399. [Google Scholar] [CrossRef]
- Lassiter, D.G.; Nylén, C.; Sjögren, R.J.O.; Chibalin, A.V.; Wallberg-Henriksson, H.; Näslund, E.; Krook, A.; Zierath, J.R. FAK tyrosine phosphorylation is regulated by AMPK and controls metabolism in human skeletal muscle. Diabetologia 2018, 61, 424–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romer, L.H.; Birukov, K.G.; Garcia, J.G.N. Focal adhesions: Paradigm for a signaling nexus. Circ. Res. 2006, 98, 606–616. [Google Scholar] [CrossRef] [Green Version]
- Geers, C.; Gros, G. Carbon dioxide transport and carbonic anhydrase in blood and muscle. Physiol. Rev. 2000, 80, 681–715. [Google Scholar] [CrossRef] [Green Version]
- Iresjö, B.M.; Landin, A.; Ohlsson, C.; Lundholm, K. Estrogen biosynthesis in cultured skeletal muscle cells (L6) induced by amino acids. Genes Nutr. 2019, 14, 29. [Google Scholar] [CrossRef] [PubMed]
- Gawlik, K.I.; Körner, Z.; Oliveira, B.M.; Durbeej, M. Early skeletal muscle pathology and disease progress in the dy3K/dy3K mouse model of congenital muscular dystrophy with laminin α2 chain-defciency. Sci. Rep. 2019, 9, 14324. [Google Scholar] [CrossRef] [PubMed]
- Maselli, R.A.; Ng, J.J.; Anderson, J.A.; Cagney, O.; Arredondo, J.; Williams, C.; Wessel, H.B.; Abdel-Hamid, H.; Wollmann, R.L. Mutations in LAMB2 causing a severe form of synaptic congenital myasthenic syndrome. J. Med. Genet. 2009, 46, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Schwander, M.; Shirasaki, R.; Pfaff, S.L.; Müller, U. β1 Integrins in muscle, but not in motor neurons, are required for skeletal muscle innervation. J. Neurosci. 2004, 23, 8181–8191. [Google Scholar] [CrossRef] [PubMed]
- Pang, Y.S.; Zhang, Z.H.; Wang, Z.; Wang, Y.X.; Yan, Y.Q.; Li, S.F.; Tong, H.L. Platelet endothelial aggregation receptor-1 regulates bovine muscle satellite cell migration and differentiation via integrin beta-1 and focal adhesion kinase. Cell Adh. Migr. 2019, 13, 192–202. [Google Scholar] [CrossRef] [Green Version]
- Han, J.W.; Lee, H.J.; Bae, G.U.; Kang, J.S. Promyogenic function of Integrin/FAK signaling is mediated by Cdo, Cdc42 and MyoD. Cell Signal. 2011, 23, 1162–1169. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Primer Name | Primer Sequence (5′-3′) | Size | Regulated |
|---|---|---|---|---|
| β-actin | F: CCCTGTATGCCTCTGGTCG | 194 bp | ||
| R: CTCGGCTGTGGTGGTGAAG | ||||
| GAPDH | F: CTGGTGCTGAATACGTTGTGG | 198 bp | ||
| R: CTGGTGCTGAATACGTTGTGG | ||||
| PE17B_vs_ PE21B | HOXC8 | F:ACTACTGCTCCCACCTTCCA | 168 bp | DOWN |
| R:AAGCGATTGCCTCGGTAGAG | ||||
| KLHL31 | F: AACCAGTGCGTGACAGTGAT | 171 bp | UP | |
| R: GCTGAAGTGGGTACGCTTCT | ||||
| PE21B_vs_ PE27B | DUSP10 | F:GGCAAGATCACCGTCTTGGA | 173 bp | UP |
| R:TCCTTGCCTTCTCGCTTGAG | ||||
| FBXO32 | F: GTCGGCAAATCTGTCCTGGT | 196 bp | UP | |
| R: GGCTAACCAGGTCTCTCCCA | ||||
| PE27B_vs_ PM6B | ADGRL3 | F:CCAATGCTCTGCTTCGTCCT | 143 bp | DOWN |
| R:GGCATTGTTCAGAAGCCCCT | ||||
| POSTN | F: GCAGGGAGCTGGAACTGAG | 148 bp | DOWN | |
| R: TGTTGCTCCTCCTTGTGTCC | ||||
| PE17L_vs_ PE21L | MFRP | F:AGTTCTGCAACCCCGTCTTC | 151 bp | DOWN |
| R:CAGGTGAACCTACAGTCGGC | ||||
| GPR37 | F: CGCCAGTCCTCCTTTTCTGT | 175 bp | DOWN | |
| R: ATTTCACGACGGATGGTGCT | ||||
| PE21L_vs_ PE27L | PCSK2 | F:TGTAGCTGAAGCATGGGAGC | 128 bp | UP |
| R:TGAAGTCGTAGCTGGCTTGG | ||||
| MYL4 | F:CCTGACCCCAAAAAGGATGC | 116 bp | DOWN | |
| R:AACTCTTCGATCTGCTCGGC | ||||
| PE27L_vs_ PM6L | TDP1 | F:GCTTGGTTCTACCCCTGGAC | 132 bp | DOWN |
| R:AACTGTCCAACAACAGGCCA | ||||
| AXIN2 | F: GCTACCAAGACCTACATAAG | 222 bp | DOWN | |
| R: GAGATAGCCACAGACAACT |
| Samples | Clean Reads | Clean Bases | GC Content | Q30 Value |
|---|---|---|---|---|
| PE17B1 | 21,975,680 | 6,554,209,528 | 51.62% | 93.01% |
| PE17B2 | 24,902,534 | 7,429,488,988 | 50.72% | 92.96% |
| PE17B3 | 27,675,557 | 8,261,513,254 | 51.50% | 93.12% |
| PE17L1 | 29,364,381 | 8,772,703,244 | 51.20% | 92.69% |
| PE17L2 | 31,153,547 | 9,309,596,992 | 51.11% | 93.22% |
| PE17L3 | 24,302,063 | 7,263,420,576 | 50.37% | 92.66% |
| PE21B1 | 22,936,235 | 6,848,636,870 | 51.28% | 92.96% |
| PE21B2 | 25,545,188 | 7,617,899,452 | 51.47% | 92.59% |
| PE21B3 | 23,616,893 | 7,047,370,582 | 50.76% | 93.11% |
| PE21L1 | 23,419,211 | 7,007,727,954 | 51.72% | 93.02% |
| PE21L2 | 21,134,566 | 6,316,620,646 | 51.41% | 93.05% |
| PE21L3 | 25,503,895 | 7,612,558,182 | 51.26% | 93.11% |
| PE27B1 | 23,860,738 | 7,133,252,550 | 50.70% | 92.86% |
| PE27B2 | 23,257,098 | 6,947,415,186 | 50.96% | 92.74% |
| PE27B3 | 21,883,412 | 6,534,290,346 | 51.66% | 92.93% |
| PE27L1 | 24,872,668 | 7,420,346,180 | 51.81% | 92.88% |
| PE27L2 | 23,784,499 | 7,108,311,764 | 51.94% | 92.91% |
| PE27L3 | 27,447,436 | 8,197,076,086 | 51.74% | 92.71% |
| PM6B1 | 27,026,755 | 8,072,713,512 | 53.75% | 92.78% |
| PM6B2 | 24,555,447 | 7,338,843,490 | 53.63% | 92.87% |
| PM6B3 | 22,180,858 | 6,629,939,152 | 54.51% | 93.24% |
| PM6L1 | 27,342,141 | 8,173,286,426 | 53.45% | 93.48% |
| PM6L2 | 25,702,190 | 7,668,643,964 | 53.48% | 93.34% |
| PM6L3 | 21,228,238 | 6,343,509,968 | 53.24% | 92.93% |
| Samples | AS Number | Samples | AS Number |
|---|---|---|---|
| PE17B1 | 42,619 | PE17L1 | 45,818 |
| PE17B2 | 43,505 | PE17L2 | 47,889 |
| PE17B3 | 45,395 | PE17L3 | 47,567 |
| PE21B1 | 41,937 | PE21L1 | 36,118 |
| PE21B2 | 42,880 | PE21L2 | 39,346 |
| PE21B3 | 43,413 | PE21L3 | 44,668 |
| PE27B1 | 43,374 | PE27L1 | 36,949 |
| PE27B2 | 45,707 | PE27L2 | 35,841 |
| PE27B3 | 38,412 | PE27L3 | 37,077 |
| PM6B1 | 40,585 | PM6L1 | 39,332 |
| PM6B2 | 35,912 | PM6L2 | 38,184 |
| PM6B3 | 32,256 | PM6L3 | 36,346 |
| Annotated Databases | New Gene Number |
|---|---|
| COG | 26 |
| GO | 97 |
| KEGG | 55 |
| KOG | 77 |
| Pfam | 68 |
| Swiss-Prot | 55 |
| eggNOG | 154 |
| Nr | 297 |
| All | 299 |
| DEG Set | Total | COG | GO | KEGG | KOG | NR | Pfam | Swiss-Prot | eggNOG |
|---|---|---|---|---|---|---|---|---|---|
| PE17B_vs_ PE21B | 358 | 123 | 279 | 225 | 247 | 357 | 324 | 263 | 344 |
| PE21B_vs_ PE27B | 1511 | 495 | 1206 | 1021 | 1070 | 1503 | 1354 | 1090 | 1446 |
| PE27B_vs_ PM6B | 3643 | 1289 | 2924 | 2448 | 2656 | 3630 | 3342 | 2635 | 3519 |
| PE17L_vs_ PE21L | 624 | 238 | 492 | 417 | 443 | 621 | 561 | 463 | 598 |
| PE21L_vs_ PE27L | 274 | 82 | 207 | 171 | 167 | 274 | 233 | 186 | 258 |
| PE27L_vs_ PM6L | 2323 | 841 | 1896 | 1614 | 1691 | 2316 | 2152 | 1744 | 2265 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Z.; Cao, J.; Ge, L.; Zhang, J.; Zhang, H.; Liu, X. Characterization and Comparative Transcriptomic Analysis of Skeletal Muscle in Pekin Duck at Different Growth Stages Using RNA-Seq. Animals 2021, 11, 834. https://doi.org/10.3390/ani11030834
Hu Z, Cao J, Ge L, Zhang J, Zhang H, Liu X. Characterization and Comparative Transcriptomic Analysis of Skeletal Muscle in Pekin Duck at Different Growth Stages Using RNA-Seq. Animals. 2021; 11(3):834. https://doi.org/10.3390/ani11030834
Chicago/Turabian StyleHu, Zhigang, Junting Cao, Liyan Ge, Jianqin Zhang, Huilin Zhang, and Xiaolin Liu. 2021. "Characterization and Comparative Transcriptomic Analysis of Skeletal Muscle in Pekin Duck at Different Growth Stages Using RNA-Seq" Animals 11, no. 3: 834. https://doi.org/10.3390/ani11030834