
Revealing New Candidate Genes for Teat Number Relevant Traits in Duroc Pigs Using Genome-Wide Association Studies

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Phenotypic Data
2.3. Genotypic Data
2.4. Quality Filtering
2.5. Statistical Models
2.6. Annotation of Candidate Genes
3. Results
3.1. Basic Description Statistics of Each Trait
3.2. Genome-Wide Association Study and Gene Annotation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hirooka, H.; De Koning, D.J.; Harlizius, B.; Van Arendonk, J.A.M.; Rattink, A.P.; Groenen, M.A.M.; Brascamp, E.W.; Bovenhuis, H. A whole-genome scan for quantitative trait loci affecting teat number in pigs. J. Anim. Sci. 2001, 79, 2320–2326. [Google Scholar] [CrossRef] [Green Version]
- Verardo, L.L.; Silva, F.F.; Lopes, M.S.; Madsen, O.; Bastiaansen, J.W.; Knol, E.F.; Kelly, M.; Varona, L.; Lopes, P.S.; Guimarães, S.E. Revealing new candidate genes for reproductive traits in pigs: Combining Bayesian GWAS and functional pathways. Genet. Sel. Evol. 2016, 48, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidanel, J.P.; Rosendo, A.; Iannuccelli, N.; Riquet, J.; Gilbert, H.; Caritez, J.C.; Billon, Y.; Amigues, Y.; Prunier, A.; Milan, D. Detection of quantitative trait loci for teat number and female reproductive traits in Meishan× Large White F2 pigs. Animal 2008, 2, 813–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toro, M.A.; Dobao, M.T.; Rodrigáñez, J.; Silió, L. Heritability of a canalized trait: Teat number in Iberian pigs. Genet. Sel. Evol. 1986, 18, 173–184. [Google Scholar] [CrossRef]
- Chalkias, H.; Rydhmer, L.; Lundeheim, N. Genetic analysis of functional and non-functional teats in a population of Yorkshire pigs. Livestock Sci. 2013, 152, 127–134. [Google Scholar] [CrossRef]
- Arakawa, A.; Okumura, N.; Taniguchi, M.; Hayashi, T.; Hirose, K.; Fukawa, K.; Ito, T.; Matsumoto, T.; Uenishi, H.; Mikawa, S. Genome-wide association QTL mapping for teat number in a purebred population of Duroc pigs. Anim. Genet. 2015, 46, 571–575. [Google Scholar] [CrossRef]
- Balzani, A.; Cordell, H.J.; Sutcliffe, E.; Edwards, S.A. Heritability of udder morphology and colostrum quality traits in swine. J. Anim. Sci. 2016, 94, 3636–3644. [Google Scholar] [CrossRef] [Green Version]
- Rohrer, G.A.; Nonneman, D.J. Genetic analysis of teat number in pigs reveals some developmental pathways independent of vertebra number and several loci which only affect a specific side. Genet. Sel. Evol. 2017, 49, 4. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, Z.; Ding, R.; Peng, L.; Wu, J.; Ye, Y.; Zhou, S.; Wang, X.; Quan, J.; Zheng, E.; Cai, G.; et al. Genome-wide association analyses identify known and novel loci for teat number in Duroc pigs using single-locus and multi-locus models. BMC Genom. 2020, 21, 344. [Google Scholar] [CrossRef]
- Tang, J.; Zhang, Z.; Yang, B.; Guo, Y.; Ai, H.; Long, Y.; Su, Y.; Cui, L.; Zhou, L.; Wang, X.; et al. Identification of loci affecting teat number by genome-wide association studies on three pig populations. Asian-Australas J. Anim. Sci. 2017, 30, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.; Da, Y.; Wu, Z.; Liu, D.; He, X.; Li, N.; Hu, X. Genome-wide association study and accuracy of genomic prediction for teat number in Duroc pigs using genotyping-by-sequencing. Genet. Sel. Evol. 2017, 49, 35. [Google Scholar] [CrossRef] [Green Version]
- Merks JW, M.; Mathur, P.K.; Knol, E.F. New phenotypes for new breeding goals in pigs. Animal 2012, 6, 535–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, I.L.; Naevdal, E.; Boe, K.E. Maternal investment, sibling competition, and offspring survival with increasing litter size and parity in pigs (Sus scrofa). Behav. Ecol. Sociobiol. 2011, 65, 1159–1167. [Google Scholar] [CrossRef] [Green Version]
- Gourley, K.M.; Calderon, H.I.; Woodworth, J.C.; DeRouchey, J.M.; Tokach, M.D.; Dritz, S.S.; Goodband, R.D. Sow and piglet traits associated with piglet survival at birth and to weaning. J. Anim. Sci. 2020, 98, skaa187. [Google Scholar] [CrossRef]
- Nielsen, B.; Su, G.; Lund, M.S.; Madsen, P. Selection for increased number of piglets at d 5 after farrowing has increased litter size and reduced piglet mortality. J Anim. Sci. 2013, 91, 2575–2582. [Google Scholar] [CrossRef] [PubMed]
- Wiegert, J.G.; Knauer, M.T. 98 Sow Functional Teat Number Impacts Colostrum Intake and Piglet Throughput. J. Anim. Sci. 2018, 96 (Suppl. 2), 51–52. [Google Scholar] [CrossRef]
- Hens, J.R.; Wysolmerski, J.J. Key stages of mammary gland development: Molecular mechanisms involved in the formation of the embryonic mammary gland. Breast. Cancer Res. 2005, 7, 220–224. [Google Scholar] [CrossRef] [Green Version]
- Lopes, M.S.; Bastiaansen, J.W.; Harlizius, B.; Knol, E.F.; Bovenhuis, H. A genome-wide association study reveals dominance effects on number of teats in pigs. PLoS ONE 2014, 9, e105867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. Gigascience 2015, 4, 7. [Google Scholar] [CrossRef]
- Yang, J.; Lee, S.H.; Goddard, M.E.; Visscher, P.M. GCTA: A tool for genome-wide complex trait analysis. Am. J. Hum Genet. 2011, 88, 76–82. [Google Scholar] [CrossRef] [Green Version]
- Aulchenko, Y.S.; Ripke, S.; Isaacs, A.; Van Duijn, C.M. GenABEL: An R library for genome-wide association analysis. Bioinformatics 2007, 23, 1294–1296. [Google Scholar] [CrossRef] [Green Version]
- Verardo, L.L.; Silva, F.F.; Varona, L.; Resende, M.D.V.; Bastiaansen, J.W.M.; Lopes, P.S.; Guimarães, S.E.F. Bayesian GWAS and network analysis revealed new candidate genes for number of teats in pigs. J. Appl. Genet. 2015, 56, 123–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, D.R.; Ren, J.; Ruan, G.F.; Guo, Y.M.; Wu, L.H.; Yang, G.C.; Zhou, L.H.; Li, L.; Zhang, Z.Y.; Huang, L.S. Mapping and fine mapping of quantitative trait loci for the number of vertebrae in a White Duroc× Chinese Erhualian intercross resource population. Anim. Genet. 2012, 43, 545–551. [Google Scholar] [CrossRef] [PubMed]
- McKay, R.M.; Rahnefeld, G.W. Rahnefeld. Heritability of Teat Number in Swine. Can. J. Anim. Sci. 1990, 70, 425–430. [Google Scholar] [CrossRef]
- Borchers, N.; Reinsch, N.; Kalm, E. Teat number, hairiness and set of ears in a Piétrain cross: Variation and effects on performance traits. Arch. Anim. Breed. 2002, 45, 465–480. [Google Scholar] [CrossRef] [Green Version]
- Moscatelli, G.; Dall’Olio, S.; Bovo, S.; Schiavo, G.; Kazemi, H.; Ribani, A.; Zambonelli, P.; Tinarelli, S.; Gallo, M.; Bertolini, F.; et al. Genome-wide association studies for the number of teats and teat asymmetry patterns in Large White pigs. Anim. Genet. 2020, 51, 595–600. [Google Scholar] [CrossRef]
- Le, T.H.; Christensen, O.F.; Nielsen, B.; Sahana, G. Genome-wide association study for conformation traits in three Danish pig breeds. Genet. Sel. Evol. 2017, 49, 12. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, M.; Yuan, J.; Zhou, X.; Xu, S.; Liu, B. Association of polymorphisms in NR6A1, PLAG1 and VRTN with the number of vertebrae in Chinese Tongcheng x Large White crossbred pigs. Anim. Genet. 2018, 49, 353–354. [Google Scholar] [CrossRef]
- Duan, Y.; Zhang, H.; Zhang, Z.; Gao, J.; Yang, J.; Wu, Z.; Fan, Y.; Xing, Y.; Li, L.; Xiao, S.; et al. VRTN is Required for the Development of Thoracic Vertebrae in Mammals. Int. J. Biol. Sci. 2018, 14, 667–681. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Liu, C.; Lan, T.; Zhang, Q.; Cao, Y.; Pu, G.; Niu, P.; Zhang, Z.; Li, Q.; Zhou, J.; et al. Polymorphism of VRTN Gene g.20311_20312ins291 Was Associated with the Number of Ribs, Carcass Diagonal Length and Cannon Bone Circumference in Suhuai Pigs. Animals 2020, 10, 484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Xing, Y.; Zhang, Z.; Ai, H.; Ouyang, Z.; Ouyang, J.; Yang, M.; Li, P.; Chen, Y.; Gao, J.; et al. A further look at porcine chromosome 7 reveals VRTN variants associated with vertebral number in Chinese and Western pigs. PLoS ONE 2013, 8, e62534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikawa, S.; Sato, S.; Nii, M.; Morozumi, T.; Yoshioka, G.; Imaeda, N.; Yamaguchi, T.; Hayashi, T.; Awata, T. Identification of a second gene associated with variation in vertebral number in domestic pigs. BMC Genet. 2011, 12, 5. [Google Scholar] [CrossRef] [Green Version]
- van Son, M.; Lopes, M.S.; Martell, H.J.; Derks, M.F.L.; Gangsei, L.E.; Kongsro, J.; Wass, M.N.; Grindflek, E.H.; Harlizius, B. A QTL for Number of Teats Shows Breed Specific Effects on Number of Vertebrae in Pigs: Bridging the Gap Between Molecular and Quantitative Genetics. Front. Genet. 2019, 10, 272. [Google Scholar] [CrossRef] [PubMed]
- Duijvesteijn, N.; Veltmaat, J.M.; Knol, E.F.; Harlizius, B. High-resolution association mapping of number of teats in pigs reveals regions controlling vertebral development. BMC Genom. 2014, 15, 542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, S.J.; Halaoui, R.; Deagle, R.C.; Rejon, C.; McCaffrey, L. Numb regulates cell tension required for mammary duct elongation. Biol. Open 2019, 8, bio042341. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.H.; Dai, W.G.; Xu, X.D.; Yu, Q.H.; Zhang, B.; Li, J.; Li, H.P. Downregulation of DPF3 promotes the proliferation and motility of breast cancer cells through activating JAK2/STAT3 signaling. Biochem. Biophys. Res. Commun. 2019, 514, 639–644. [Google Scholar] [CrossRef]
- Kerros, C.; Tripathi, S.C.; Zha, D.; Mehrens, J.M.; Sergeeva, A.; Philips, A.V.; Qiao, N.; Peters, H.L.; Katayama, H.; Sukhumalchandra, P.; et al. Neuropilin-1 mediates neutrophil elastase uptake and cross-presentation in breast cancer cells. J. Biol. Chem. 2017, 292, 10295–10305. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Traits | Mean ± sd | Rang | Pedigree-Based Heritability (SE) | Genomic-Based Heritability (SE) |
|---|---|---|---|---|
| LTN | 6.55 ± 0.67 | 5–9 | 0.178 (0.060) | 0.148 (0.044) |
| RTN | 6.61 ± 0.68 | 5–8 | 0.228 (0.067) | 0.177 (0.045) |
| TTN | 13.17 ± 1.12 | 10–16 | 0.321 (0.077) | 0.289 (0.053) |
| MAX | 6.83 ± 0.63 | 5–9 | 0.255 (0.072) | 0.220 (0.051) |
| LR | 0.062 ± 0.76 | −2–2 | 0.000 (0.028) | 0.000 (0.024) |
| ALR | 0.49 ± 0.59 | 0–2 | 0.017 (0.032) | 0.054 (0.034) |
| SLR | - | - | 0.010 (0.030) | 0.032 (0.028) |
| Trait | Chromosome | SNP | Position (bp) | p Value | Gene |
|---|---|---|---|---|---|
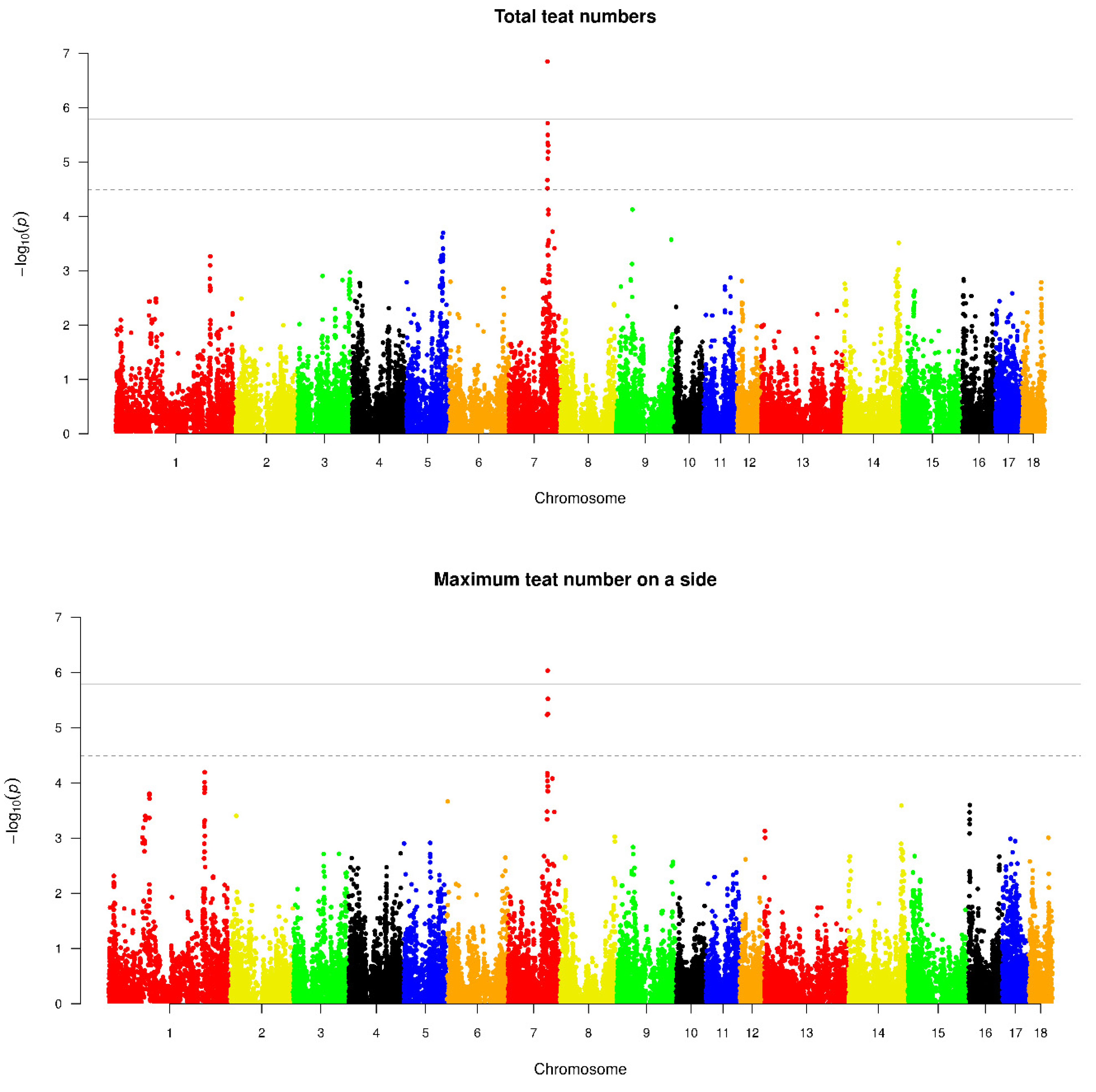
| RTN | 7 | H3GA0022664 | 98066911 | 5.03 × 10−7 | YLPM1, PROX2(within), DLST, RPS6KL1 |
| 7 | ASGA0035527 | 98089286 | 5.66 × 10−7 | YLPM1, PROX2, DLST, RPS6KL1 | |
| 7 | INRA0027601 | 96278617 | 6.55 × 10−7 | DPF3, ZFYVE1 | |
| 7 | MARC0038565 | 97652632 | 7.17 × 10−7 | VRTN, SYNDIG1L | |
| 7 | MARC0048752 | 97946666 | 4.06 × 10−6 | AREL1, FCF1(within), YLPM1 | |
| 7 | M1GA0010654 | 97954258 | 6.52 × 10−6 | AREL1, FCF1(within), YLPM1 | |
| 7 | ALGA0043962 | 97973860 | 6.52 × 10−6 | AREL1, FCF1, YLPM1 | |
| 7 | INRA0027603 | 97048513 | 7.06 ×10−6 | DNAL1(within), MIDEAS | |
| 7 | MARC0033479 | 96786713 | 2.39 × 10−5 | NUMB(within), HEATR4, RIOX1 | |
| 7 | DRGA0008025 | 96731837 | 1.87 × 10−5 | NUMB(within) | |
| 7 | ALGA0043926 | 96806775 | 2.28 × 10−5 | NUMB, HEATR4, RIOX1 | |
| LTN | 7 | MARC0038565 | 97652632 | 3.66 × 10−6 | VRTN, SYNDIG1L |
| MAX | 7 | MARC0038565 | 97652632 | 9.28 × 10−7 | VRTN, SYNDIG1L |
| 7 | H3GA0022664 | 98066911 | 5.67 × 10−6 | YLPM1, PROX2(within), DLST, RPS6KL1 | |
| 7 | ASGA0035527 | 98089286 | 2.99 × 10−6 | YLPM1, PROX2, DLST, RPS6KL1 | |
| 7 | INRA0027601 | 96278617 | 5.86 × 10−6 | DPF3, ZFYVE1 | |
| TTN | 7 | MARC0038565 | 97652632 | 8.67 × 10−9 | VRTN, SYNDIG1L |
| 7 | H3GA0022664 | 98066911 | 6.40 × 10−8 | YLPM1, PROX2(within), DLST, RPS6KL1 | |
| 7 | ASGA0035527 | 98089286 | 5.22 × 10−8 | YLPM1, PROX2, DLST, RPS6KL1 | |
| 7 | INRA0027601 | 96278617 | 1.41 × 10−7 | DPF3, ZFYVE1 | |
| 7 | MARC0033479 | 96786713 | 4.44 × 10−6 | NUMB(within), HEATR4, RIOX1 | |
| 7 | DRGA0008025 | 96731837 | 3.19 × 10−6 | NUMB(within) | |
| 7 | ALGA0043926 | 96806775 | 1.93 × 10−6 | NUMB, HEATR4, RIOX1 | |
| 7 | MARC0048752 | 97946666 | 4.93 × 10−6 | AREL1, FCF1(within), YLPM1 | |
| 7 | M1GA0010654 | 97954258 | 6.52 × 10−6 | AREL1, FCF1(within), YLPM1 | |
| 7 | ALGA0043962 | 97973860 | 6.52 × 10−6 | AREL1, FCF1, YLPM1 | |
| 7 | INRA0027603 | 97048513 | 8.66 × 10−6 | DNAL1(within), MIDEAS | |
| 7 | M1GA0010637 | 96101509 | 2.16 × 10−5 | DPF3(within) | |
| 7 | ALGA0043904 | 96128654 | 3.06 × 10−5 | DPF3(within) | |
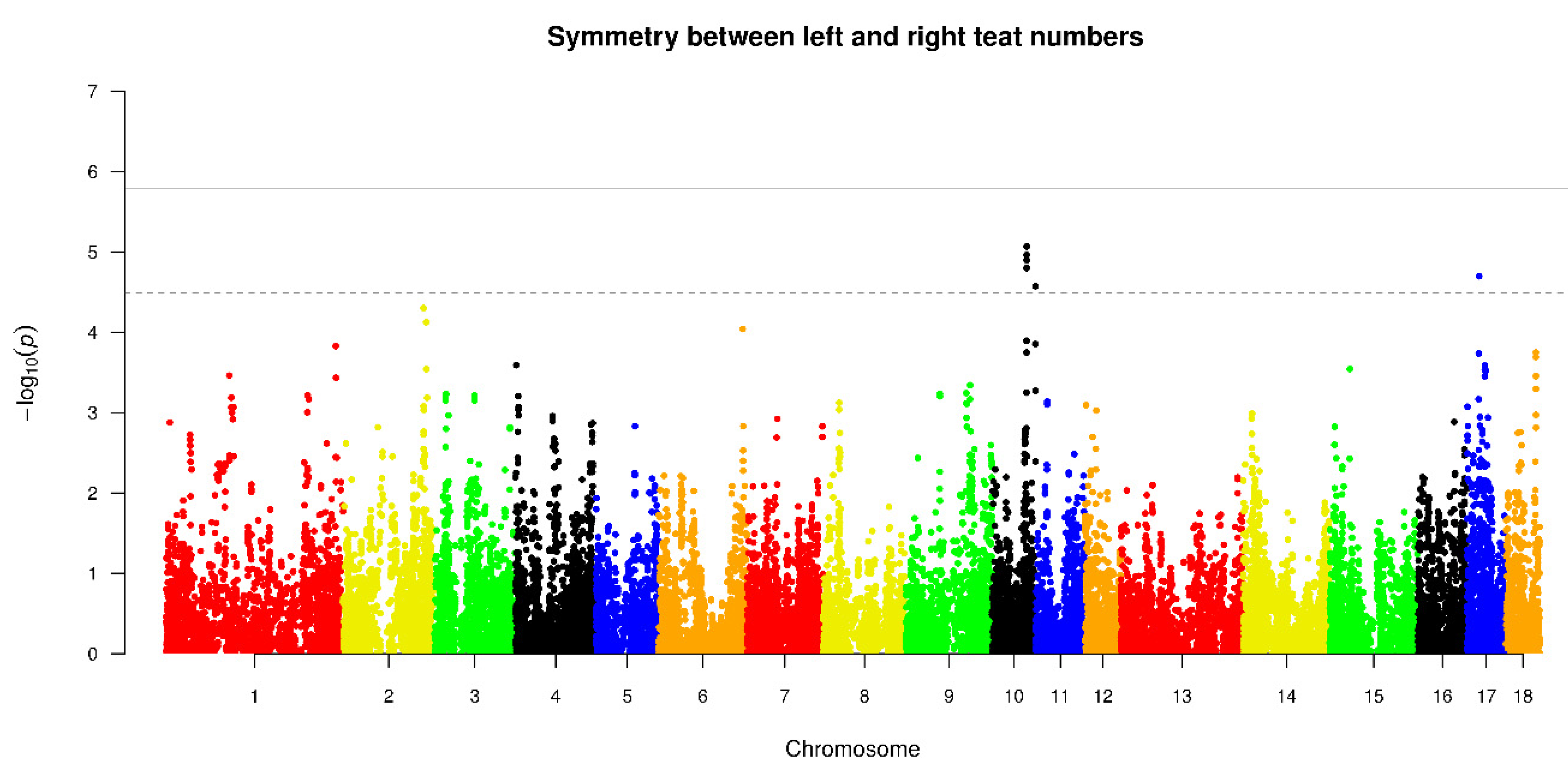
| SLR | 10 | MARC0112026 | 56303425 | 8.58 × 10−6 | NRP1(within) |
| 10 | ALGA0102459 | 56421545 | 8.58 × 10−6 | NRP1(within) | |
| 10 | ASGA0090893 | 56211944 | 1.09 × 10−5 | ITBG1 | |
| 10 | MARC0010633 | 56282915 | 1.27 × 10−5 | NRP1(within) | |
| 10 | MARC0052641 | 56340616 | 1.27 × 10−5 | NRP1(within) | |
| 17 | H3GA0048042 | 19474175 | 2.01 × 10−5 | - | |
| 10 | ASGA0089631 | 56237188 | 1.59 × 10−5 | NRP1 | |
| 10 | ASGA0098692 | 56259450 | 1.59 × 10−5 | NRP1 | |
| 10 | MARC0068931 | 55591111 | 2.67 × 10−5 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Pu, L.; Shi, L.; Gao, H.; Zhang, P.; Wang, L.; Zhao, F. Revealing New Candidate Genes for Teat Number Relevant Traits in Duroc Pigs Using Genome-Wide Association Studies. Animals 2021, 11, 806. https://doi.org/10.3390/ani11030806
Li Y, Pu L, Shi L, Gao H, Zhang P, Wang L, Zhao F. Revealing New Candidate Genes for Teat Number Relevant Traits in Duroc Pigs Using Genome-Wide Association Studies. Animals. 2021; 11(3):806. https://doi.org/10.3390/ani11030806
Chicago/Turabian StyleLi, Yang, Lei Pu, Liangyu Shi, Hongding Gao, Pengfei Zhang, Lixian Wang, and Fuping Zhao. 2021. "Revealing New Candidate Genes for Teat Number Relevant Traits in Duroc Pigs Using Genome-Wide Association Studies" Animals 11, no. 3: 806. https://doi.org/10.3390/ani11030806