
Hormonal Carcinogenesis in Canine Mammary Cancer: Molecular Mechanisms of Estradiol Involved in Malignant Progression


{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Canine Mammary Cancer
2. E2 and Mammary Cancer
2.1. Genomic Effects Associated with Estradiol
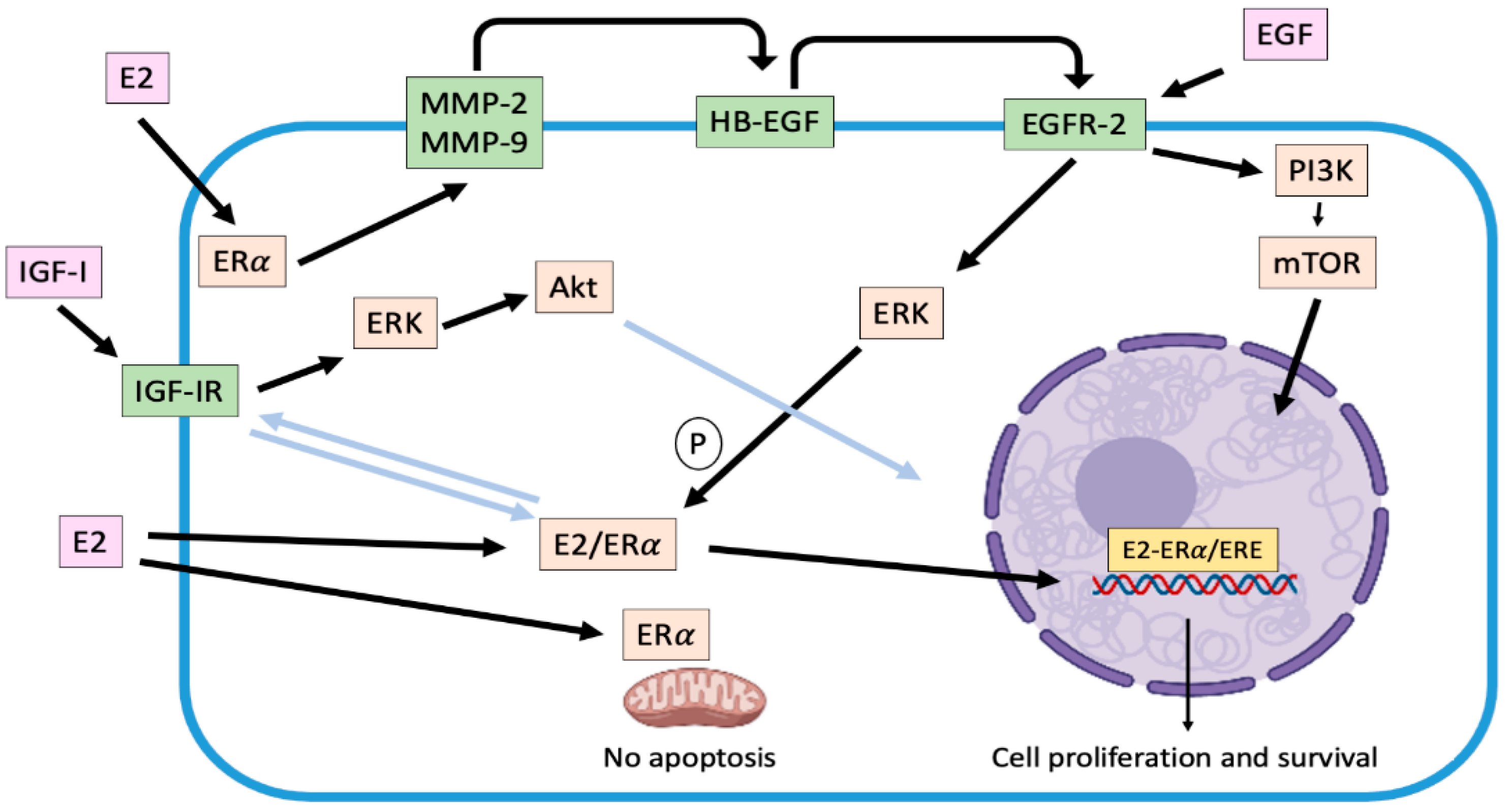
2.2. Non-Genomic Effects Associated with Estradiol
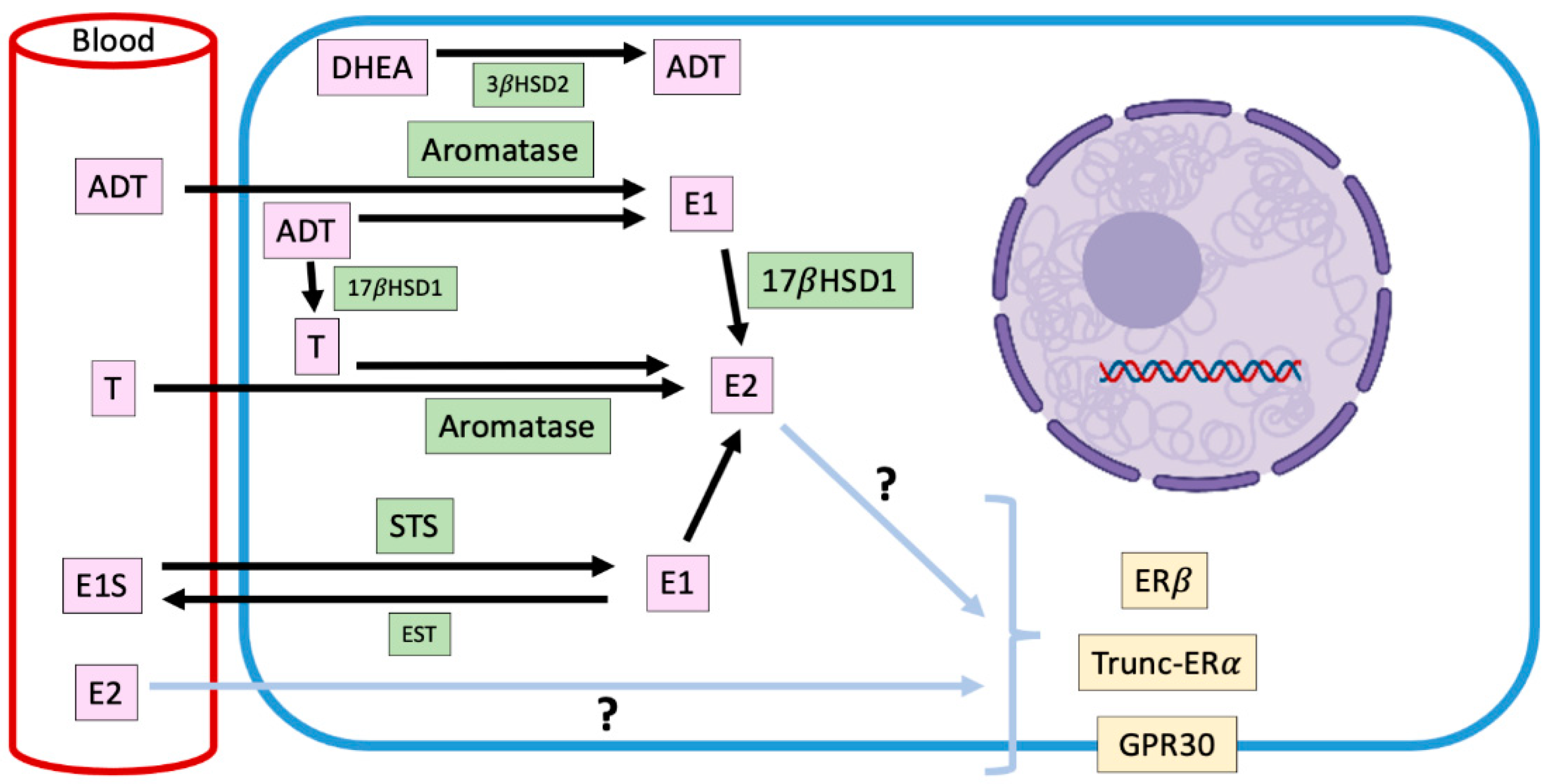
2.3. Intracrinology of Estradiol in Canine Mammary Tumors
3. Future Perspectives
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pang, L.Y.; Cervantes-Arias, A.; Else, R.W.; Argyle, D.J. Canine mammary cancer stem cells are radio- and chemo-resistant and exhibit an epithelial-mesenchymal transition phenotype. Cancers 2011, 3, 1744–1762. [Google Scholar] [CrossRef] [PubMed]
- Egenvall, A.; Bonnett, B.N.; Öhagen, P.; Olson, P.; Hedhammar, A.; Von Euler, H. Incidence of and survival after mammary tumors in a population of over 80.000 insured female dogs in Sweden from 1995 to 2002. Prev. Vet. Med. 2005, 69, 109–127. [Google Scholar] [CrossRef]
- Merlo, D.F.; Rossi, L.; Pellegrino, C.; Ceppi, M.; Cardellino, U.; Capurro, C.; Ratto, A.; Sambucco, P.L.; Sestito, V.; Tanara, G.; et al. Cancer incidence in pet dogs: Findings of the Animal Tumor Registry of Genoa, Italy. J. Vet. Intern. Med. 2008, 22, 976–984. [Google Scholar] [CrossRef]
- Burrai, G.P.; Gabrieli, A.; Moccia, V.; Zappulli, V.; Porcellato, I.; Brachelente, C.; Pirino, S.; Polinas, M.; Antuofermo, E. A Statistical Analysis of Risk Factors and Biological Behavior in Canine Mammary Tumors: A Multicenter Study. Animals 2020, 10, 1687. [Google Scholar] [CrossRef] [PubMed]
- Rivera, P.; Melin, M.; Biagi, T.; Fall, T.; Haggstrom, J.; Lindblad-Toh, K.; von Euler, H. Mammary tumor development in dogs is associated with BRCA1 and BRCA2. Cancer Res. 2009, 69, 8770–8774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canadas, A.; Santos, M.; Pinto, R.; Medeiros, R.; Dias-Pereira, P. Catechol-o-methyltransferase genotypes are associated with progression and biological behaviour of canine mammary tumours. Vet. Comp. Oncol. 2018, 16, 664–669. [Google Scholar] [CrossRef]
- Sorenmo, K.U.; Rasotto, R.; Zappulli, V.; Goldschmidt, M.H. Development, anatomy, histology, lymphatic drainage, clinical features, and cell differentiation markers of canine mammary gland neoplasms. Vet. Pathol. 2011, 48, 85–97. [Google Scholar] [CrossRef] [Green Version]
- Schneider, R.; Dorn, C.R.; Taylor, D.O.N. Factors influencing canine mammary cancer development and postsurgical survival. J. Natl. Cancer Inst. 1969, 43, 1249–1261. [Google Scholar]
- Klopfleisch, R.; von Euler, H.; Sarli, G.; Pinho, S.S.; Gärtner, F.; Gruber, A.D. Molecular carcinogenesis of canine mammary tumors: News from an old disease. Vet. Pathol. 2011, 48, 98–116. [Google Scholar] [CrossRef]
- Kristiansen, V.M.; Peña, L.; Díez Córdova, L.; Illera, J.C.; Skjerve, E.; Breen, A.M.; Cofone, M.A.; Langeland, M.; Teige, J.; Goldschmidt, M.; et al. Effect of Ovariohysterectomy at the Time of Tumor Removal in Dogs with Mammary Carcinomas: A Randomized Controlled Trial. J. Vet. Intern. Med. 2016, 30, 230–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enginler, S.O.; Akis, I.; Toydemir, T.S.; Oztabak, K.; Haktanir, D.; Gunduz, M.C.; Kirsan, I.; Firat, I. Genetic variations of BRCA1 and BRCA2 genes in dogs with mammary tumours. Vet. Res. Commun. 2014, 38, 21–27. [Google Scholar] [CrossRef]
- Thumser-Henner, P.; Nytko, K.J.; Rohrer Bley, C. Mutations of BRCA2 in canine mammary tumors and their targeting potential in clinical therapy. BMC Vet. Res. 2020, 16, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.M.; Yang, I.S.; Seung, B.J.; Lee, S.; Kim, D.; Ha, Y.J.; Seo, M.K.; Kim, K.K.; Kim, H.S.; Cheong, J.H.; et al. Cross-species oncogenic signatures of breast cancer in canine mammary tumors. Nat. Commun. 2020, 11, 3616. [Google Scholar] [CrossRef]
- Canadas-Sousa, A.; Santos, M.; Leal, B.; Medeiros, R.; Dias-Pereira, P. Estrogen receptors genotypes and canine mammary neoplasia. BMC Vet. Res. 2019, 15, 325. [Google Scholar] [CrossRef]
- Vascellari, M.; Capello, K.; Carminato, A.; Zanardelo, C.; Baioni, E.; Mutinelli, F. Incidence of mammary tumors in the canine population living in the Veneto region (Northeastern Italy): Risk factors and similarities to human breast cancer. Prev. Vet. Med. 2016, 126, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Salas, Y.; Márquez, A.; Diaz, D.; Romero, L. Epidemiological Study of Mammary Tumors in Female Dogs Diagnosed during the Period 2002–2012: A Growing Animal Health Problem. PLoS ONE 2015, 10, e0127381. [Google Scholar] [CrossRef] [Green Version]
- Litterine-Kaufman, J.; Casale, S.A.; Mouser, P.J. Prevalence of malignancy in masses from the mammary gland region of dogs with single or multiple masses. J. Am. Vet. Med. Assoc. 2019, 255, 817–820. [Google Scholar] [CrossRef] [PubMed]
- Goldschmidt, M.; Peña, L.; Rasotto, R.; Zappulli, V. Classification and grading of canine mammary tumors. Vet. Pathol. 2011, 88, 117–131. [Google Scholar] [CrossRef]
- Rasotto, R.; Berlato, D.; Goldschmidt, M.H.; Zappulli, V. Prognostic Significance of Canine Mammary Tumor Histologic Subtypes: An Observational Cohort Study of 229 Cases. Vet. Pathol. 2015, 54, 571–578. [Google Scholar] [CrossRef]
- Canadas, A.; França, M.; Pereira, C.; Vilaça, R.; Vilhena, H.; Tinoco, F.; Silva, M.J.; Ribeiro, J.; Medeiros, R.; Oliveira, P.; et al. Canine Mammary Tumors: Comparison of Classification and Grading Methods in a Survival Study. Vet. Pathol. 2019, 56, 208–219. [Google Scholar] [CrossRef]
- Cassali, G.D.; Jark, P.C.; Gamba, C.; Damasceno, K.A.; Estrela-Lima, A.; De Nardi, A.B.; Ferreira, E.; Horta, R.S.; Firmo, B.F.; Sueiro, F.A.R.; et al. Consensus Regarding the Diagnosis, Prognosis and Treatment of Canine and Feline Mammary Tumors–2019. Braz. J. Vet. Pathol. 2020, 13, 555–574. [Google Scholar] [CrossRef]
- Nguyen, F.; Peña, L.; Ibisch, C.; Loussouarn, D.; Gama, A.; Rieder, N.; Belousov, A.; Campone, M.; Abadie, J. Canine invasive mammary carcinomas as models of human breast cancer. Part 1: Natural history and prognostic factors. Breast Cancer Res. Treat. 2018, 167, 635–648. [Google Scholar] [CrossRef] [Green Version]
- Abdelmegeed, S.M.; Mohammed, S. Canine mammary tumors as a model for human disease. Oncol. Lett. 2018, 15, 8195–8205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaszak, I.; Ruszczak, A.; Kanafa, S.; Kacprzak, K.; Król, M.; Jurka, P. Current biomarkers of canine mammary tumors. Acta Vet. Scand. 2018, 60, 66. [Google Scholar] [CrossRef] [Green Version]
- Brunetti, B.; Bacci, B.; Angeli, C.; Benazzi, C.; Muscatello, L.V. p53, ER and ki67 expression in canine mammary carcinomas and correlation with pathological variables and prognosis. Vet. Pathol. 2020, 19. [Google Scholar] [CrossRef]
- Rossi Varallo, G.; Gelaleti, G.B.; Maschio-Signorini, L.B.; Moschetta, M.G.; Lopes, J.R.; De Nardi, A.B.; Tinucci-Costa, M.; Rocha, R.M.; De Campos Zuccari, D.A.P. Prognostic phenotypic classification for canine mammary tumors. Oncol. Lett. 2019, 18, 6545–6553. [Google Scholar]
- Gama, A.; Alves, A.; Schmitt, F. Identification of molecular phenotypes in canine mammary carcinomas with clinical implications: Application of the human classification. Virchows Arch. 2008, 453, 123–132. [Google Scholar] [CrossRef]
- Torres, C.; Antileo, E.; Epuñan, M.J.; Pino, A.M.; Valladares, L.E.; Sierralta, W.D. A cyclic peptide derived from alpha fetoprotein inhibits the proliferative effects of EGF and estradiol in MCF7 cells. Oncol. Rep. 2008, 19, 1597–1604. [Google Scholar] [PubMed]
- Torres, C.G. Antiproliferative Effect of a Cyclopeptide Derived from α-Fetoprotein on Canine Mammary Tumor Cells. Ph.D. Thesis, Universidad de Chile, Santiago, Chile, 2009. [Google Scholar]
- Sorenmo, K.U.; Durham, A.C.; Radaelli, E.; Kristiansen, V.; Peña, L.; Goldschmidt, M.H.; Stefanovski, D. The estrogen effect; clinical and histopathological evidence of dichotomous influences in dogs with spontaneous mammary carcinomas. PLoS ONE 2019, 14, e0224504. [Google Scholar] [CrossRef] [Green Version]
- Spoerri, M.; Guscetti, F.; Hartnack, S.; Boos, A.; Oei, C.; Balogh, O.; Nowaczyk, R.M.; Michel, E.; Reichlerand, I.M.; Kowalewski, M.P. Endocrine control of canine mammary neoplasms: Serum reproductive hormone levels and tissue expression of steroid hormone, prolactin and growth hormone receptors. BMC Vet. Res. 2015, 11, 235. [Google Scholar] [CrossRef] [Green Version]
- Brandão, Y.O.; Toledo, M.B.; Chequin, A.; Cristo, T.G.; Sousa, R.S.; Ramos, E.A.S.; Klassen, G. DNA Methylation Status of the Estrogen Receptor α Gene in Canine Mammary Tumors. Vet. Pathol. 2018, 55, 510–516. [Google Scholar] [CrossRef] [Green Version]
- Prossnitz, E.R.; Arterburn, J.B.; Smith, H.O.; Oprea, T.I.; Sklar, L.A.; Hathaway, H.J. Estrogen signaling through the transmembrane G protein-coupled receptor GPR30. Annu. Rev. Physiol. Mar. 2008, 17, 165–190. [Google Scholar] [CrossRef] [PubMed]
- Levin, E.R.; Hammes, S.R. Nuclear receptors outside the nucleus: Extranuclear signalling by steroid receptors. Nat. Rev. Mol. Cell Biol. 2016, 17, 783–797. [Google Scholar] [CrossRef] [Green Version]
- Caceres, S.; Monsalve, B.; Peña, L.; de Andres, P.J.; Alonso-Diez, A.; Illera, M.J.; Woodward, W.A.; Reuben, J.M.; Silvan, G.; Illera, J.C. In vitro and in vivo effect of flutamide on steroid hormone secretion in canine and human inflammatory breast cancer cell lines. Vet. Comp. Oncol. 2018, 16, 148–158. [Google Scholar] [CrossRef]
- Martin de las Mulas, J.; Ordás, J.; Millán, M.Y.; Chacón, M.; De Lara, F.; Espinoza de los Monteros, A.; Reymundo, C.; Jover, A. Inmunohistochemical expression of estradiol receptor β in normal and tumoral mammary glands. Vet. Pathol. 2004, 41, 269–272. [Google Scholar] [CrossRef] [PubMed]
- De Andrés, P.J.; Cáceres, S.; Clemente, M.; Pérez-Alenza, M.D.; Illera, J.C.; Peña, L. Profile of steroid receptors and increased aromatasa immunoexpression in canine inflammatory mammary cancer as a potential therapeutic target. Reprod. Domest. Anim. 2016, 51, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Calaf, G.M.; Echiburú-Chau, C. Synergistic effect of malathion and estrogen on mammary gland carcinogenesis. Oncol. Rep. 2012, 28, 640–646. [Google Scholar] [CrossRef] [Green Version]
- Torres, C.G.; Pino, A.M.; Sierralta, W.D. A cyclized peptide derived from alpha fetoprotein inhibits the proliferation of ER-positive canine mammary cancer cells. Oncol. Rep. 2009, 21, 1397–1404. [Google Scholar]
- De Andres, P.J. Comparative Aspects Between Human and Canine Mammary Cancer with Special Reference to Inflammatory Mammary Cancer: Histological Grade of Malignancy, Endocrine Mechanisms and Interleukins. Ph.D. Thesis, Universidad Complutense, Madrid, Spain, 2016. [Google Scholar]
- Yager, J.D.; Davidson, N.E. Estrogen Carcinogenesis in Breast Cancer. N. Engl. J. Med. 2006, 354, 270–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobiela, J.; Stefaniak, T.; Krajewsky, J.; Kalinska-Blach, B.; Zuawa-Janicka, D.; Lachinsky, A.; Gackosky, D.; Olinsky, R.; Nowap, J.; Knap, N.; et al. Dynamics of oxidative damage al early stages of estrogen-dependent carcinogenesis. Adv. Exp. Med. Biol. 2008, 617, 609–615. [Google Scholar]
- Marinelli, L.; Gabai, G.; Wolfswinkel, J.; Mol, J.A. Mammary steroid metabolizing enzymes in relation to hyperplasia and tumorigenesis in the dog. J. Steroid Biochem. Mol. Biol. 2004, 92, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Queiroga, F.L.; Pérez-Alenza, M.D.; Silvan, G.; Peña, L.; Lopes, C.; Illera, J.C. Role of steroid hormones and prolactin in canine mammary cancer. J. Steroid Biochem. Mol. Biol. 2005, 94, 181–187. [Google Scholar] [CrossRef]
- Queiroga, F.L.; Pérez-Alenza, D.; González-Gil, A.; Silvan, G.; Peña, L.; Illera, J.C. Serum and Tissue Steroid Hormone Levels in Canine Mammary Tumours: Clinical and Prognostic Implications. Reprod. Dom. Anim. 2015, 50, 858–865. [Google Scholar] [CrossRef] [PubMed]
- Illera, J.C.; Pérez-Alenza, M.D.; Nieto, A.; Jiménez, M.A.; Silvana, G.; Dunnerc, S.; Peña, L. Steroids and receptors in canine mammary cancer. Steroids 2006, 71, 541–548. [Google Scholar] [CrossRef]
- Samavat, H.; Kurzer, M.S. Estrogen Metabolism and Breast Cancer. Cancer Lett. 2015, 356, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Chamkasem, A.; Toniti, W. Sequence to Structure Approach of Estrogen Receptor Alpha and Ligand Interactions. Asian Pac. J. Cancer Prev. 2015, 16, 2161–2166. [Google Scholar] [CrossRef] [Green Version]
- Tavarez, W.; Lavalle, G.; Figueiredo, M.; Souza, A.; Bertagnolli, A.; Paes, P.; Carneiro, R.; Cavalcanti, G.; Melo, M.; Cassali, G. Evaluation of adverse effects in tamoxifen exposed healthy female dogs. Acta Veterinaria Scandinavica 2010, 52, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, K.; Horie-Inoue, K.; Inoue, S. Identification of estrogen-responsive genes based on the DNA binding properties of estrogen receptors using high-throughput sequencing technology. Acta Pharmacologica Sinica 2015, 36, 24–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, E.R. Membrane estrogen receptors signal to determine transcription factor function. Steroids 2018, 132, 1–4. [Google Scholar] [CrossRef]
- Razandi, M.; Pedram, A.; Park, S.T.; Levin, E.R. Proximal events in signaling by plasma membrane estrogen receptors. J. Biol. Chem. 2003, 278, 2701–2712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, E.R. Bidirectional signaling between the estrogen receptor and the epidermal growth factor receptor. Mol. Endocrinol. 2003, 17, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Jorissen, R.N.; Walker, F.; Pouliot, N.; Garrett, T.; Ward, C.W.; Burgess, A.W. Epidermal growth factor Receptor: Mechanisms of Activation and Signaling. Exp. Cell Res. 2003, 284, 31–53. [Google Scholar] [CrossRef]
- Yu, Z.; Gao, W.; Jiang, E.; Lu, F.; Zhang, L.; Shi, Z.; Wang, X.; Chen, L.; Lv, T. Interaction between IGF-IR and ER Induced by E2 and IGF-I. PLoS ONE 2013, 8, e62642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queiroga, F.L.; Pérez-Alenza, M.D.; Silvan, G.; Peña, L.; Lopes, C.S.; Illera, J.C. Crosstalk between GH/IGF-I axis and steroid hormones (progesterone, 17β-estradiol) in canine mammary tumours. J. Steroid Biochem. Mol. Biol. 2008, 110, 76–82. [Google Scholar] [CrossRef]
- Jaillardon, L.; Abadie, J.; Godard, T.; Campone, M.; Loussouarn, D.; Siliart, B.; Nguyen, F. The dog as a naturally-occurring model for insulin-like growth factor type 1 receptor-overexpressing breast cancer: An observational cohort study. BMC Cancer 2015, 15, 664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedram, A.; Razandi, M.; Wallace, D.C.; Levin, E.R. Functional estrogen receptors in the mitochondria of breast cancer cells. Mol. Cell Biol. 2006, 17, 2125–2137. [Google Scholar] [CrossRef] [Green Version]
- Filardo, E.J.; Quinn, J.A.; Sabo, E. Association of the membrane estrogen receptor, GPR30, with breast tumor metastasis and transactivation of the epidermal growth factor receptor. Steroids 2008, 73, 870–873. [Google Scholar] [CrossRef] [PubMed]
- Filardo, E.J.; Graeber, C.T.; Quinn, J.A.; Resnick, M.B.; Giri, D.; DeLlelis, R.A.; Steinhoff, M.M.; Sabo, E. Distribution of GPR30, a seven-membrane spanning estrogen receptor, in primary breast cancer and its association with clinicopathologic determinants of tumor progression. Clin. Cancer Res. 2006, 12, 6359–6366. [Google Scholar] [CrossRef] [Green Version]
- Maggiolini, M.; Picard, D. The unfolding stories of GPR30, a new membrane-bound estrogen receptor. J. Endocrinol. 2010, 204, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Molina, L.; Figueroa, C.D.; Bhoola, K.D.; Ehrenfeld, P. GPER-1/GPR30 a novel estrogen receptor sited in the cell membrane: Therapeutic coupling to breast cancer. Expert. Opin. Ther. Targets 2017, 21, 755–766. [Google Scholar] [CrossRef]
- Sasano, H.; Suzuki, T.; Miki, Y.; Moriya, T. Intracrinology of estrogens and androgens in breast carcinoma. J. Steroid Biochem. Mol. Biol. 2008, 108, 181–185. [Google Scholar] [CrossRef]
- Queiroga, F.L.; Pérez-Alenza, D.; Silvan, G.; Peña, L.; Illera, J.C. Positive correlation of steroid hormones and EGF in canine mammary cancer. J. Steroid Biochem. Mol. Biol. 2009, 115, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Bulun, S.E.; Chen, D.; Moy, I.; Brooks, D.C.; Zhao, H. Aromatase, breast cancer and obesity: A complex interaction. Trends Endocrinol. Metab. 2012, 23, 83–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, H.-Y.; Im, K.-S.; Kim, N.H.; Kim, H.-W.; Shin, J.-I.; Yhee, J.-Y.; Sur, J.-H. Effects of Obesity and Obesity-Related Molecules on Canine Mammary Gland Tumors. Vet. Pathol. 2015, 52, 1045–1051. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Archidona, A.R.; Jiménez, M.A.; Pérez-Alenza, D.; Silván, G.; Illera, J.C.; Peña, L.; Dunner, S. Steroid pathway and oestrone sulphate production in canine inflammatory mammary carcinoma. J. Steroid Biochem. Mol. Biol. 2007, 104, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Nakata, T.; Miki, Y. Estrogen sulfotransferase and steroid sulfatase in human breast carcinoma. Cancer Res. 2003, 63, 2762–2770. [Google Scholar] [PubMed]
- McNamara, K.M.; Guestini, F.; Sauer, T.; Touma, J.; Bukholm, I.R.; Lindstrøm, J.C.; Sasano, H.; Geisler, J. In breast cancer subtypes steroid sulfatase (STS) is associated with less aggressive tumour characteristics. Br. J. Cancer 2018, 118, 1208–1216. [Google Scholar] [CrossRef]
- Hilborn, E.; Stal, O.; Jansson, A. Estrogen and androgen-converting enzymes 17β-hydroxysteroid dehydrogenase and their involvement in cancer: With a special focus on 17β-hydroxysteroid dehydrogenase type 1, 2, and breast cancer. Oncotarget 2017, 8, 30552–30562. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres, C.G.; Iturriaga, M.P.; Cruz, P. Hormonal Carcinogenesis in Canine Mammary Cancer: Molecular Mechanisms of Estradiol Involved in Malignant Progression. Animals 2021, 11, 608. https://doi.org/10.3390/ani11030608
Torres CG, Iturriaga MP, Cruz P. Hormonal Carcinogenesis in Canine Mammary Cancer: Molecular Mechanisms of Estradiol Involved in Malignant Progression. Animals. 2021; 11(3):608. https://doi.org/10.3390/ani11030608
Chicago/Turabian StyleTorres, Cristian G., María P. Iturriaga, and Pamela Cruz. 2021. "Hormonal Carcinogenesis in Canine Mammary Cancer: Molecular Mechanisms of Estradiol Involved in Malignant Progression" Animals 11, no. 3: 608. https://doi.org/10.3390/ani11030608