
The Click Production of Captive Yangtze Finless Porpoises (Neophocaena asiaeorientalis asiaorientalis) Is Influenced by Social and Environmental Factors

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Animals and Captive Management
2.2. Data Collection
2.3. Analysis
2.3.1. Echolocation Signal Analysis
2.3.2. Statistical Analysis
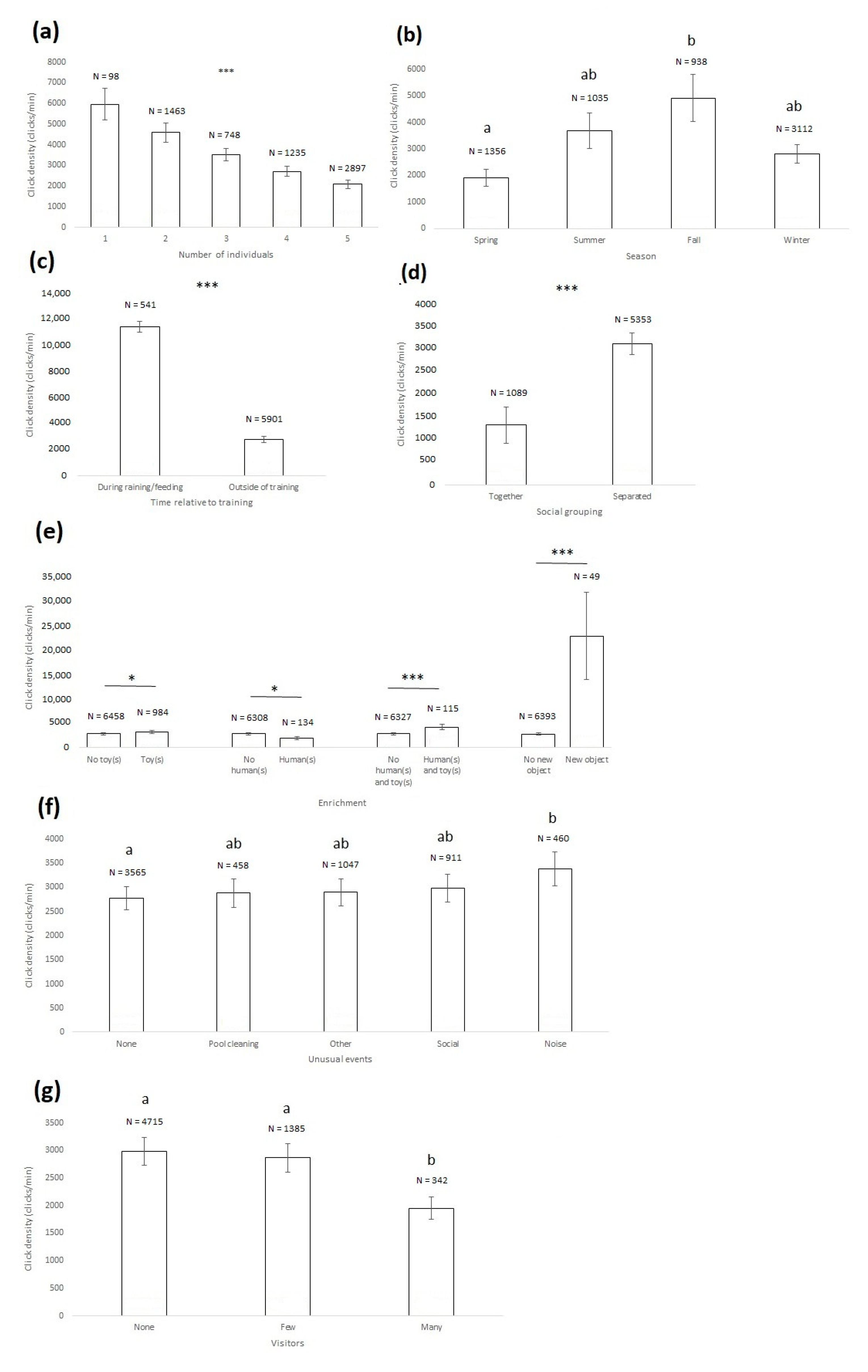
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tyack, P.L.; Clark, C.W. Communication and acoustic behavior of dolphins and whales. In Hearing by Whales and Dolphins; Springer: New York, NY, USA, 2000; pp. 156–224. [Google Scholar]
- Richardson, W.J.; Greene, C.R., Jr.; Malme, C.I.; Thomson, D.H. Marine Mammals and Noise; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Au, W.W. Characteristics of dolphin sonar signals. In The Sonar of Dolphins; Springer: Berlin/Heidelberg, Germany, 2003; pp. 115–139. [Google Scholar]
- Lammers, M.O.; Au, W.W.; Herzing, D.L. The broadband social acoustic signaling behavior of spinner and spotted dolphins. J. Acoust. Soc. Am. 2003, 114, 1629–1639. [Google Scholar] [CrossRef]
- Jensen, F.H.; Beedholm, K.; Wahlberg, M.; Bejder, L.; Madsen, P.T. Estimated communication range and energetic cost of bottlenose dolphin whistles in a tropical habitat. J. Acoust. Soc. Am. 2012, 131, 582–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akamatsu, T.; Wang, D.; Wang, K.; Naito, Y. Biosonar behaviour of free-ranging porpoises. Proc. R. Soc. B: Boil. Sci. 2005, 272, 797–801. [Google Scholar] [CrossRef] [Green Version]
- Dawson, S.M. Clicks and communication: The behavioural and social contexts of Hector’s dolphin vocalizations. Ethology 1991, 88, 265–276. [Google Scholar] [CrossRef]
- Madsen, P.T.; Carder, D.A.; Bedholm, K.; Ridgway, S.H. Porpoise clicks from a sperm whale nose—Convergent evolution of 130 kHz pulses in toothed whale sonars? Bioacoustics 2005, 15, 195–206. [Google Scholar] [CrossRef]
- Clausen, K.T.; Wahlberg, M.; Beedholm, K.; Deruiter, S.; Madsen, P.T. Click communication in harbour porpoises Phocoena phocoena. Bioacoustics 2011, 20, 1–28. [Google Scholar] [CrossRef]
- Esch, H.C.; Sayigh, L.S.; Blum, J.E.; Wells, R.S. Whistles as potential indicators of stress in bottlenose dolphins (Tursiops truncatus). J. Mammal. 2009, 90, 638–650. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, E.R.; Gartside, D.F. Whistle emissions of Indo-Pacific bottlenose dolphins (Tursiops aduncus) differ with group composition and surface behaviors. J. Acoust. Soc. Am. 2010, 127, 2652–2663. [Google Scholar] [CrossRef] [PubMed]
- Platto, S.; Wang, D.; Wang, K. Variation in the emission rate of sounds in a captive group of false killer whales Pseudorca crassidens during feedings: Possible food anticipatory vocal activity? Chin. J. Oceanol. Limnol. 2015, 34, 1218–1237. [Google Scholar] [CrossRef]
- Hatakeyama, Y.; Soeda, H. Studies on echolocation of porpoises taken in salmon gillnet fisheries. In Sensory Abilities of Cetaceans; Springer: Boston, MA, USA, 1990; pp. 269–281. [Google Scholar]
- Silber, G. Acoustic signal of the vaquita (Phocoena sinus). Aquat. Mamm. 1991, 17, 130–133. [Google Scholar]
- Au, W.W.L.; Kastelein, R.A.; Rippe, T.; Schooneman, N.M. Transmission beam pattern and echolocation signals of a harbor porpoise (Phocoena phocoena). J. Acoust. Soc. Am. 1999, 106, 3699–3705. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Wang, D.; Wang, K.; Akamatsu, T.; Ma, Z.; Han, J. Echolocation click sounds from wild inshore finless porpoise (Neophocaena phocaenoides sunameri) with comparisons to the sonar of riverine N. p. asiaeorientalis. J. Acoust. Soc. Am. 2007, 121, 3938–3946. [Google Scholar] [CrossRef] [PubMed]
- Villadsgaard, A.; Wahlberg, M.; Tougaard, J. Echolocation signals of wild harbour porpoises, Phocoena phocoena. J. Exp. Biol. 2007, 210, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Jefferson, T.A.; Webber, M.A.; Pitman, R.L. Marine Mammals of the World: A Comprehensive Guide to Their Identification; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Bassett, H.R.; Baumann, S.; Campbell, G.S.; Wiggins, S.M.; Hildebrand, J.A. Dall’s porpoise (Phocoenoides dalli) echolocation click spectral structure. J. Acoust. Soc. Am. 2009, 125, 2677. [Google Scholar] [CrossRef]
- Dawson, S.M. The high frequency sounds of free-ranging Hector s dolphins, Cephalorhynchus hectori. Rep. Int. Whal. Comm. Spec. Issue 1988, 9, 339–344. [Google Scholar]
- Kyhn, L.A.; Tougaard, J.; Jensen, F.; Wahlberg, M.; Stone, G.; Yoshinaga, A.; Beedholm, K.; Madsen, P.T. Feeding at a high pitch: Source parameters of narrow band, high-frequency clicks from echolocating off-shore hourglass dolphins and coastal Hector’s dolphins. J. Acoust. Soc. Am. 2009, 125, 1783–1791. [Google Scholar] [CrossRef] [Green Version]
- Kyhn, L.A.; Jensen, F.H.; Beedholm, K.; Tougaard, J.; Hansen, M.; Madsen, P.T. Echolocation in sympatric Peale’s dolphins (Lagenorhynchus australis) and Commerson’s dolphins (Cephalorhynchus commersonii) producing narrow-band high-frequency clicks. J. Exp. Biol. 2010, 213, 1940–1949. [Google Scholar] [CrossRef] [Green Version]
- Götz, T.; Antunes, R.; Heinrich, S. Echolocation clicks of free-ranging Chilean dolphins (Cephalorhynchus eutropia). J. Acoust. Soc. Am. 2010, 128, 563–566. [Google Scholar] [CrossRef] [Green Version]
- Morisaka, T.; Karczmarski, L.; Akamatsu, T.; Sakai, M.; Dawson, S.; Thornton, M. Echolocation signals of Heaviside’s dolphins (Cephalorhynchus heavisidii). J. Acoust. Soc. Am. 2011, 129, 449–457. [Google Scholar] [CrossRef] [Green Version]
- Reyes Reyes, M.V.; Iñíguez, M.A.; Hevia, M.; Hildebrand, J.A.; Melcón, M.L. Description and clustering of echolocation signals of Commerson’s dolphins (Cephalorhynchus commersonii) in Bahía San Julián, Argentina. J. Acoust. Soc. Am. 2015, 138, 2046–2053. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, Y.M.; Morisaka, T.; Sakai, M.; Iwasaki, M.; Wakabayashi, I.; Seko, A.; Kasamatsu, M.; Akamatsu, T.; Kohshima, S. Sound variation and function in captive Commerson’s dolphins (Cephalorhynchus commersonii). Behav. Process. 2014, 108, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Norris, K.S.; Wursig, B.; Wells, R.S.; Wursig, M. The Hawaiian spinner dolphin. University of California Press: Berkeley, CA, USA, 1994; 436p. [Google Scholar]
- Barrett-Lennard, L.G.; Ford, J.K.; Heise, K.A. The mixed blessing of echolocation: Differences in sonar use by fish-eating and mammal-eating killer whales. Anim. Behav. 1996, 51, 553–565. [Google Scholar] [CrossRef] [Green Version]
- Van Parijs, S.M.; Corkeron, P.J. Vocalizations and behaviour of Pacific humpback dolphins Sousa chinensis. Ethology 2001, 107, 701–716. [Google Scholar] [CrossRef]
- Nowacek, D.P. Acoustic ecology of foraging bottlenose dolphins (Tursiops truncatus), habitat-specific use of three sound types. Mar. Mammal Sci. 2005, 21, 587–602. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Royle, J.A.; Pollock, K.H.; Bailey, L.; Hines, J.E. Occupancy Estimation and Modeling: Inferring Patterns and Dynamics of Species Occurrence; Elsevier: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Lima, A.; Lemasson, A.; Boye, M.; Hausberger, M. Vocal activities reflect the temporal distribution of bottlenose dolphin social and non-social activity in a zoological park. Zoo Biol. 2017, 36, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Nowacek, D.P.; Thorne, L.H.; Johnston, D.W.; Tyack, P.L. Responses of cetaceans to anthropogenic noise. Mammal Rev. 2007, 37, 81–115. [Google Scholar] [CrossRef]
- Thompson, P.M.; Brookes, K.L.; Graham, I.M.; Barton, T.R.; Needham, K.; Bradbury, G.; Merchant, N.D. Short-term disturbance by a commercial two-dimensional seismic survey does not lead to long-term displacement of harbour porpoises. Proc. R. Soc. B Boil. Sci. 2013, 280, 20132001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lusseau, D. Effects of tour boats on the behavior of bottlenose dolphins: Using Markov chains to model anthropogenic impacts. Conserv. Biol. 2003, 17, 1785–1793. [Google Scholar] [CrossRef]
- Pirotta, E.; Brookes, K.L.; Graham, I.M.; Thompson, P.M. Variation in harbour porpoise activity in response to seismic survey noise. Biol. Lett. 2014, 10, 20131090. [Google Scholar] [CrossRef] [Green Version]
- Van Parijs, S.M.; Clark, C.W.; Sousa-Lima, R.S.; Parks, S.E.; Rankin, S.; Risch, D.; Van Opzeeland, I.C. Management and research applications of real-time and archival passive acoustic sensors over varying temporal and spatial scales. Mar. Ecol. Prog. Ser. 2009, 395, 21–36. [Google Scholar] [CrossRef] [Green Version]
- Marques, T.A.; Thomas, L.; Martin, S.W.; Mellinger, D.K.; Ward, J.A.; Moretti, D.J.; Harris, D.V.; Tyack, P.L. Estimating animal population density using passive acoustics. Biol. Rev. 2012, 88, 287–309. [Google Scholar] [CrossRef] [Green Version]
- Kimura, S.; Akamatsu, T.; Wang, D.; Li, S.; Wang, K.; Yoda, K. Variation in the production rate of biosonar signals in freshwater porpoises. J. Acoust. Soc. Am. 2013, 133, 3128–3134. [Google Scholar] [CrossRef] [PubMed]
- Ralston, J.V.; Herman, L.M. Dolphin auditory perception. In The Comparative Psychology of Audition: Perceiving Complex Sounds; Lawrence Erlbaum Associates: Hillsdale, NJ, USA, 1989; pp. 295–328. [Google Scholar]
- Herzing, D.L. Acoustics and social behavior of wild dolphins: Implications for a sound society. In Hearing by Whales and Dolphins; Springer: New York, NY, USA, 2000; pp. 225–272. [Google Scholar]
- Akiyama, J.; Ohta, M. Increased number of whistles of bottlenose dolphins, Tursiops truncatus, arising from interaction with people. J. Vet. Med. Sci. 2007, 69, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Therrien, S.C.; Thomas, J.A.; Therrien, R.E.; Stacey, R. Time of Day and Social Change Affect Underwater Sound Production by Bottlenose Dolphins (Tursiops truncatus) at the Brookfield Zoo. Aquat. Mamm. 2012, 38, 65–75. [Google Scholar] [CrossRef]
- Boinski, S.; Gross, T.S.; Davis, J.K. Terrestrial predator alarm vocalizations are a valid monitor of stress in captive brown capuchins (Cebus apella). Zoo Biol. 1999, 18, 295–312. [Google Scholar] [CrossRef]
- Mansour, A.A.; Zakaria, A.H.; Fraser, A.F. Effect of enclosure quality on reactivity and welfare of captive Soemmerring’s gazelle (Gazella soemmerringii). J. Appl. Anim. Welf. Sci. 2000, 3, 335–343. [Google Scholar] [CrossRef]
- Taylor, A.A.; Weary, D.M. Vocal responses of piglets to castration: Identifying procedural sources of pain. Appl. Anim. Behav. Sci. 2000, 69, 255–264. [Google Scholar] [CrossRef]
- Zimmerman, P.H.; Koene, P.; Van Hooff, J.A. Thwarting of behaviour in different contexts and the gakel-call in the laying hen. Appl. Anim. Behav. Sci. 2000, 69, 255–264. [Google Scholar] [CrossRef]
- Zimmerman, P.H.; Lundberg, A.; Keeling, L.J.; Koene, P. The effect of an audience on the gakel-call and other frustration behaviours in the laying hen (Gallus gallus domesticus). Anim. Welf. Potters Bar Wheathampstead 2003, 12, 315–326. [Google Scholar]
- Schon, P.C.; Puppe, B.; Manteuffel, G. Automated recording of stress vocalisations as a tool to document impaired welfare in pigs. Anim. Welf. Potters Bar Wheathampstead 2004, 13, 105–110. [Google Scholar]
- McCowan, B.; Rommeck, I.; Anderson, K.; Heagerty, A.; Cameron, A. Bioacoustic monitoring of aggression in group-housed rhesus macaques. J. Appl. Anim. Welf. Sci. 2006, 9, 261–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golosova, O.S.; Volodin, I.A.; Isaeva, I.L.; Volodina, E.V. Effects of free-ranging, semi-captive and captive management on the acoustics of male rutting calls in Siberian wapiti Cervus elaphus sibiricus. Mammal Res. 2017, 62, 387–396. [Google Scholar] [CrossRef]
- Castellote, M.; Fossa, F. Measuring acoustic activity as a method to evaluate welfare in captive beluga whales (Delphinapterus leucas). Aquat. Mamm. 2006, 32, 325–333. [Google Scholar] [CrossRef]
- Clegg, I.; Borger-Turner, J.; Eskelinen, H. C-Well: The development of a welfare assessment index for captive bottlenose dolphins (Tursiops truncatus). Anim. Welf. 2015, 24, 267–282. [Google Scholar] [CrossRef]
- Li, S.; Wang, K.; Wang, D.; Akamatsu, T. Echolocation signals of the free-ranging Yangtze finless porpoise (Neophocaena phocaenoides asiaeorientialis). J. Acoust. Soc.Am. 2005, 117, 3288–3296. [Google Scholar] [CrossRef]
- Wang, K.; Wang, D.; Akamatsu, T.; Li, S.; Xiao, J. A passive acoustic monitoring method applied to observation and group size estimation of finless porpoises. J. Acoust. Soc. Am. 2005, 118, 1180–1185. [Google Scholar] [CrossRef] [Green Version]
- Kimura, S.; Akamatsu, T.; Li, S.; Dong, S.; Dong, L.; Wang, K.; Wang, D.; Arai, N. Density estimation of Yangtze finless porpoises using passive acoustic sensors and automated click train detection. J. Acoust. Soc. Am. 2010, 128, 1435–1545. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.-T.; Akamatsu, T.; Mei, Z.; Dong, L.; Imaizumi, T.; Wang, K.; Wang, D. Frequent and prolonged nocturnal occupation of port areas by Yangtze finless porpoises (Neophocaena asiaeorientalis): Forced choice for feeding? Integr. Zool. 2015, 10, 122–132. [Google Scholar] [CrossRef]
- Mei, Z.; Huang, S.-L.; Hao, Y.; Turvey, S.T.; Gong, W.; Wang, D. Accelerating population decline of Yangtze finless porpoise (Neophocaena asiaeorientalis asiaeorientalis). Biol. Conserv. 2012, 153, 192–200. [Google Scholar] [CrossRef]
- Wang, D. Population status, threats and conservation of the Yangtze finless porpoise. Chin. Sci. Bull. 2009, 54, 3473–3484. [Google Scholar] [CrossRef] [Green Version]
- Fang, L.; Wang, D.; Li, Y.; Cheng, Z.; Pine, M.K.; Wang, K.; Li, S. The source parameters of echolocation clicks from captive and free-ranging yangtze finless porpoises (neophocaena asiaeorientalis asiaeorientalis). PLoS ONE 2015, 10, e0129143. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Wang, D.; Wang, K.; Li, S.; Dong, S.; Zhao, X.; Akamatsu, T.; Kimura, S. Passive acoustic survey of Yangtze finless porpoises using a cargo ship as a moving platform. J. Acoust. Soc. Am. 2011, 130, 2285–2292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akamatsu, T.; Wang, D.; Nakamura, K.; Wang, K. Echolocation range of captive and free- ranging baiji (Lipotes vexillifer), finless porpoise (Neophocaena phocaenoides), and bottlenose dolphin (Tursiops truncatus). J. Acoust. Soc. Am. 1998, 104, 2511–2516. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2014, 67, 7–48. [Google Scholar]
- Akaike, H. Maximum likelihood identification of Gaussian autoregressive moving average models. Biometrika 1973, 60, 255–265. [Google Scholar] [CrossRef]
- Verboom, W.C.; Kastelein, R.A. Structure of Harbor Porpoise (Phocoena phocoena) Acoustic Signals with High Repetition Rates. In Echolocation in Bats and Dolphins; Thomas, J.A., Moss, C., Vater, M., Eds.; University of Chicago Press: Chicago, IL, USA, 2003; pp. 40–42. [Google Scholar]
- Koschinski, S.; Diederichs, A.; Amundin, M. Click train patterns of free-ranging harbour porpoises acquired using T-PODs may be useful as indicators of their behaviour. J. Cetacean Res. Manag. 2008, 10, 147–155. [Google Scholar]
- Verfuss, U.K.; Miller, L.A.; Pilz, P.K.D.; Schnitzler, H.-U.; Verfuß, U.K. Echolocation by two foraging harbour porpoises (Phocoena phocoena). J. Exp. Biol. 2009, 212, 823–834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soldevilla, M.S.; Wiggins, S.M.; Hildebrand, J.A. Spatial and temporal patterns of Risso’s dolphin echolocation in the Southern California Bight. J. Acoust. Soc. Am. 2010, 127, 124–132. [Google Scholar] [CrossRef]
- Wei, Z.; Wang, D.; Zhang, X.; Wang, K.; Chen, D.; Zhao, Q.; Kuang, X.; Gong, W.; Wang, X. Observation on some sexual behavior of the Yangtze finless porpoise (Neophocaena phocaenoides asiaerientals) in captivity. Acta Theriol. Sin. 2004, 24, 98–102. [Google Scholar]
- Wang, Z.; Akamatsu, T.; Wang, K.; Wang, D. The diel rhythms of biosonar behavior in the Yangtze finless porpoise (Neophocaena asiaeorientalis asiaeorientalis) in the port of the Yangtze River: The correlation between prey availability and boat traffic. PLoS ONE 2014, 9, e97907. [Google Scholar] [CrossRef]
- Rachinas-Lopes, P.; Luís, A.R.; Borges, A.S.; Neto, M.; Dos Santos, M.E. Whistle Stability and Variation in Captive Bottlenose Dolphins (Tursiops truncatus) Recorded in Isolation and Social Contexts. Aquat. Mamm. 2017, 43, 1–13. [Google Scholar] [CrossRef]
- Tyack, P. Whistle repertoires of two bottlenosed dolphins, Tursiops truncatus: Mimicry of signature whistles? Behav. Ecol. Sociobiol. 1986, 18, 251–257. [Google Scholar] [CrossRef]
- Janik, V.M.; Todt, D.; Dehnhardt, G. Signature whistle variations in a bottlenosed dolphin, Tursiops truncatus. Behav. Ecol. Sociobiol. 1994, 35, 243–248. [Google Scholar] [CrossRef]
- Sekiguchi, Y.; Kohshima, S. Resting behaviors of captive bottlenose dolphins (Tursiops truncatus). Physiol. Behav. 2003, 79, 643–653. [Google Scholar] [CrossRef]
- Lopez Marulanda, J.; Adam, O.; Delfour, F. Modulation of whistle production related to training sessions in bottlenose dolphins (Tursiops truncatus) under human care. Zoo Biol. 2016, 35, 495–504. [Google Scholar] [CrossRef]
- Fish, M.P.; Mowbray, W.H. Production of Underwater Sound by the White Whale or Beluga, Delphinapterus Leucas (Pallus) (No. TR17R62 5); Rhode Island University Kingston Narragansett Marine Lab: Narragansett, RI, USA, 1962. [Google Scholar]
- Brickman, J.J. Factors Influencing Whistle Usage of the Pacific White-Sided Dolphin, Lagenorhynchus Obliquidens, at the John G. Shedd Aquarium. Ph.D. Thesis, Western Illinois University, Macomb, IL, USA, 2003. [Google Scholar]
- Miller, P.J.O.; Johnson, M.P.; Tyack, P.L. Sperm whale behaviour indicates the use of echolocation click buzzes ‘creaks’ in prey capture. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2004, 271, 2239–2247. [Google Scholar] [CrossRef]
- Wahlberg, M.; Jensen, F.H.; Aguilar Soto, N.; Beedholm, K.; Bejder, L.; Oliveira, C.; Rasmussen, M.; Simon, M.; Villadsgaard, A.; Madsen, P.T. Source parameters of echolocation clicks from wild bottlenose dolphins (Tursiops aduncus and Tursiops truncatus). J. Acoust. Soc. Am. 2011, 130, 2263–2274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirotta, E.; Thompson, P.M.; Miller, P.I.; Brookes, K.L.; Cheney, B.; Barton, T.R.; Graham, I.M.; Lusseau, D. Scale-dependent foraging ecology of a marine top predator modelled using passive acoustic data. Funct. Ecol. 2014, 28, 206–217. [Google Scholar] [CrossRef] [Green Version]
- Ellison, W.; Southall, B.; Clark, C.; Frankel, A. A new context-based approach to assess marine mammal behavioral responses to anthropogenic sounds. Conserv. Biol. 2011, 26, 21–28. [Google Scholar] [CrossRef]
- Pirotta, E.; Merchant, N.D.; Thompson, P.M.; Barton, T.R.; Lusseau, D. Quantifying the effect of boat disturbance on bottlenose dolphin foraging activity. Biol. Conserv. 2015, 181, 82–89. [Google Scholar] [CrossRef]
- Buckstaff, K.C. Effects of watercraft noise on the acoustic behavior of bottlenose dolphins, Tursiops truncatus, in Sarasota Bay, Florida. Mar. Mammal Sci. 2004, 20, 709–725. [Google Scholar] [CrossRef]
- David, J. Likely sensitivity of bottlenose dolphins to pile-driving noise. Water Environ. J. 2006, 20, 48–54. [Google Scholar] [CrossRef]
- Serres, A. The frequency of solitary behaviours in captive odontocetes is modulated by environmental and social factors. Int. J. Comp. Psychol. 2019, 32. [Google Scholar] [CrossRef]
- Serres, A.; Hao, Y.; Wang, D. Swimming features in captive odontocetes: Indicative of animals’ emotional state? Behav. Process. 2020, 170, 103998. [Google Scholar] [CrossRef]
- Serres, A.; Hao, Y.; Wang, D. Body Contacts and Social Interactions in Captive Odontocetes Are Influenced by the Context: An Implication for Welfare Assessment. Animals 2020, 10, 924. [Google Scholar] [CrossRef] [PubMed]
- Tavolga, W.N. Theoretical principles for the study of communication in cetaceans. Mammalia 1983, 47, 3–26. [Google Scholar] [CrossRef]
- Johnson, C. Animal communication via coordinated cognitive systems. Perspect. Ethol. 1993, 10, 187–207. [Google Scholar]
- Herzing, D.L. Vocalizations and associated underwater behavior of free-ranging Atlantic spotted dolphins, Stenella frontalis and bottlenose dolphins, Tursiops truncatus. Aquat. Mamm. 1996, 22, 61–80. [Google Scholar]
- Busnel, R.G.; Dziedzic, A. Acoustic signals of the pilot whale Globicephala melaena and of the porpoises Delphinus delphis and Phocoena phocoena. In Whales, Dolphins and Porpoises; University of California Press: Oakland, CA, USA, 1966; pp. 607–646. [Google Scholar]
- Dreher, J.J.; Evans, W.E. Cetacean communication. Mar. Bioacoust. 1964, 1, 373–393. [Google Scholar]
- Acevedo-Gutiérrez, A.; Stienessen, S.C. Bottlenose dolphins (Tursiops truncatus) increase number of whistles when feeding. Aquat. Mamm. 2004, 30, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, M.E.D.; Louro, S.; Couchinho, M.N.; Brito, C.M. Whistles of bottlenose dolphins (Tursiops truncatus) in the Sado Estuary, Portugal: Characteristics, production rates, and long-term contour stability. Aquat. Mamm. 2005, 31, 453–462. [Google Scholar] [CrossRef]
- Akamatsu, T.; Hatakeyama, Y.; Kojima, T.; Soeda, H. Echolocation rates of two harbor porpoises (Phocoena phocoena). Mar. Mammal Sci. 1994, 10, 401–411. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Name | Sex | Age (year) | Length (m/h) |
|---|---|---|---|
| Duoduo | M | 8 | 157 |
| F7 * | F | 8 | 145 |
| F9 * | F | 8 | 145 |
| Taotao | M | 14 | 156 |
| Yangyang * | F | 11 | 147 |
| Season | Time of Day | Time Relative to Training * | Social Grouping | Enrichment | Unusual Events | Visitors | |
|---|---|---|---|---|---|---|---|
| Winter (December–February) | Morning (8 am–11 pm) | Outside of training | Together | None | None | None | |
| Spring (March–May) | Noon (11 pm–2 pm) | During training | Separated | Toys a | Pool cleaning d | Few h | |
| Summer (June–August) | Afternoon (2 pm–5 pm) | Humans b | Noise e | Many i | |||
| Fall (September–November) | Humans and toys | Social event f | |||||
| Live fish | Other event g | ||||||
| New object c | |||||||
| (a) | Model Components | Model Characteristics | Scaled Residuals | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Response Variable | Offset | Random | Predictors | AIC | BIC | logLik | Deviance | df.resid | Min | 1Q | Median | 3Q | Max | |
| factors | ||||||||||||||
| Selected model | Click density | Number of individuals | Date, | Number of individuals, | 107,583.40 | 107,717.10 | −53,771.7 | 107,543.40 | 5878 | −4.1621 | −0.0065 | 0.0019 | 0.0093 | 1.0357 |
| Observation session ID | Season, | |||||||||||||
| Time relative to training, | ||||||||||||||
| Social grouping, | ||||||||||||||
| Presence of toys, | ||||||||||||||
| Presence of humans, | ||||||||||||||
| Presence of humans and toys, | ||||||||||||||
| Presence of new objects, | ||||||||||||||
| Unusual events, | ||||||||||||||
| Presence of visitors | ||||||||||||||
| (b) | Number of individuals | Season | Social grouping | Presence of toys | Presence of humans | Presence of humans and toys | Presence of new objects | Time relative to training | Unusual events | Presence of public | ||||
| Selected model | χ2 = 48.687, df = 1, p < 0.0001 | χ2 = 17.076, df = 3, p = 0.0007 | χ2 = 18.709, df = 1, p < 0.0001 | χ2 = 5.256, df = 1, p = 0.0218 | χ2 = 6.580, df = 1, p = 0.0103 | χ2 = 13.205, df = 1, p = 0.0003 | χ2 = 30.067, df = 1, p < 0.0001 | χ2 = 617.822, df = 1, p < 0.0001 | χ2 = 8.915, df = 4, p = 0.0634 | χ2 = 40.891, df = 2, p < 0.0001 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serres, A.; Xu, C.; Hao, Y.; Wang, D. The Click Production of Captive Yangtze Finless Porpoises (Neophocaena asiaeorientalis asiaorientalis) Is Influenced by Social and Environmental Factors. Animals 2021, 11, 511. https://doi.org/10.3390/ani11020511
Serres A, Xu C, Hao Y, Wang D. The Click Production of Captive Yangtze Finless Porpoises (Neophocaena asiaeorientalis asiaorientalis) Is Influenced by Social and Environmental Factors. Animals. 2021; 11(2):511. https://doi.org/10.3390/ani11020511
Chicago/Turabian StyleSerres, Agathe, Chen Xu, Yujiang Hao, and Ding Wang. 2021. "The Click Production of Captive Yangtze Finless Porpoises (Neophocaena asiaeorientalis asiaorientalis) Is Influenced by Social and Environmental Factors" Animals 11, no. 2: 511. https://doi.org/10.3390/ani11020511