
A Seasonal Study of Koi Herpesvirus and Koi Sleepy Disease Outbreaks in the United Kingdom in 2018 Using a Pond-Side Test

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. DNA Control and Recombinant Plasmids
2.2. LAMP Primers Design
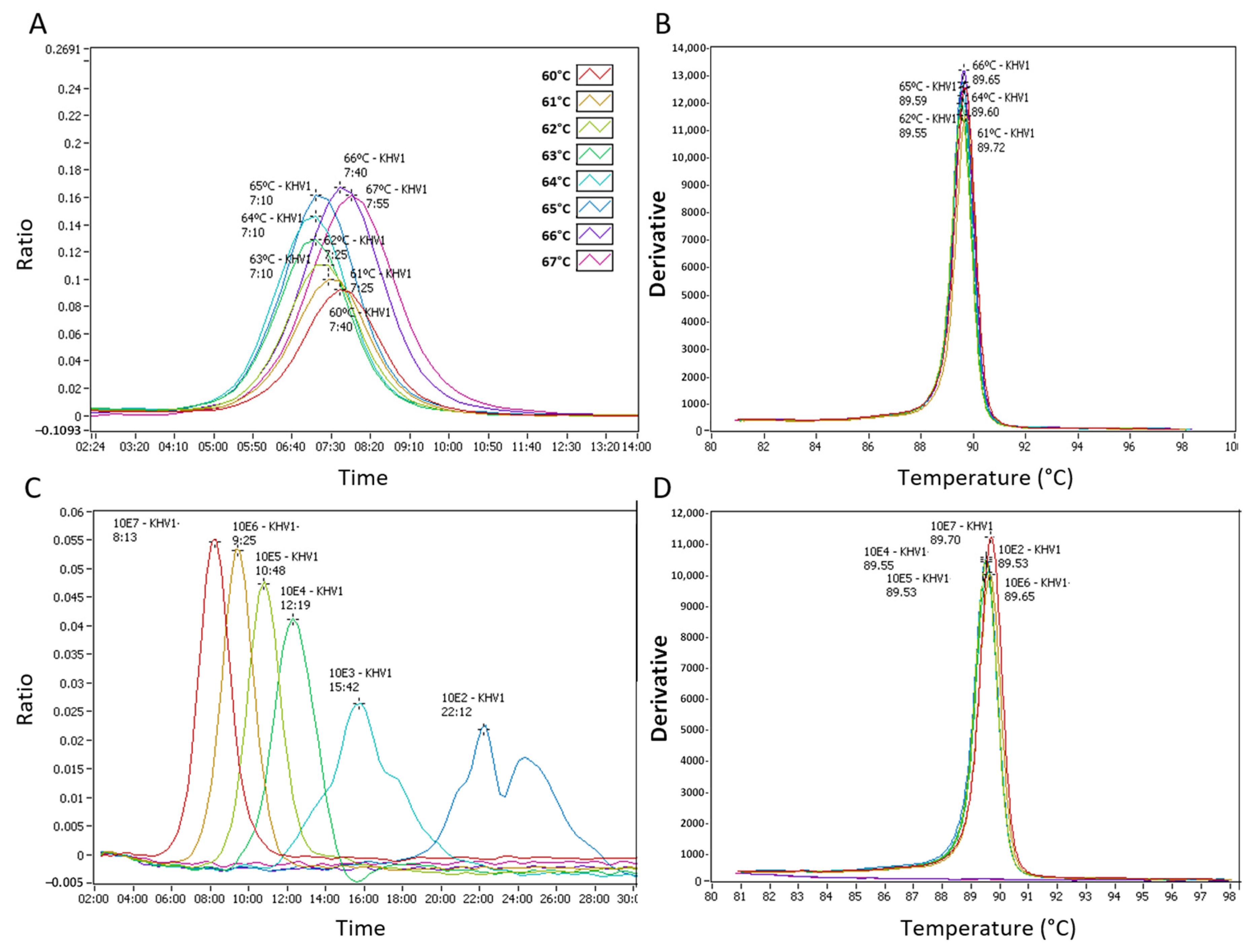
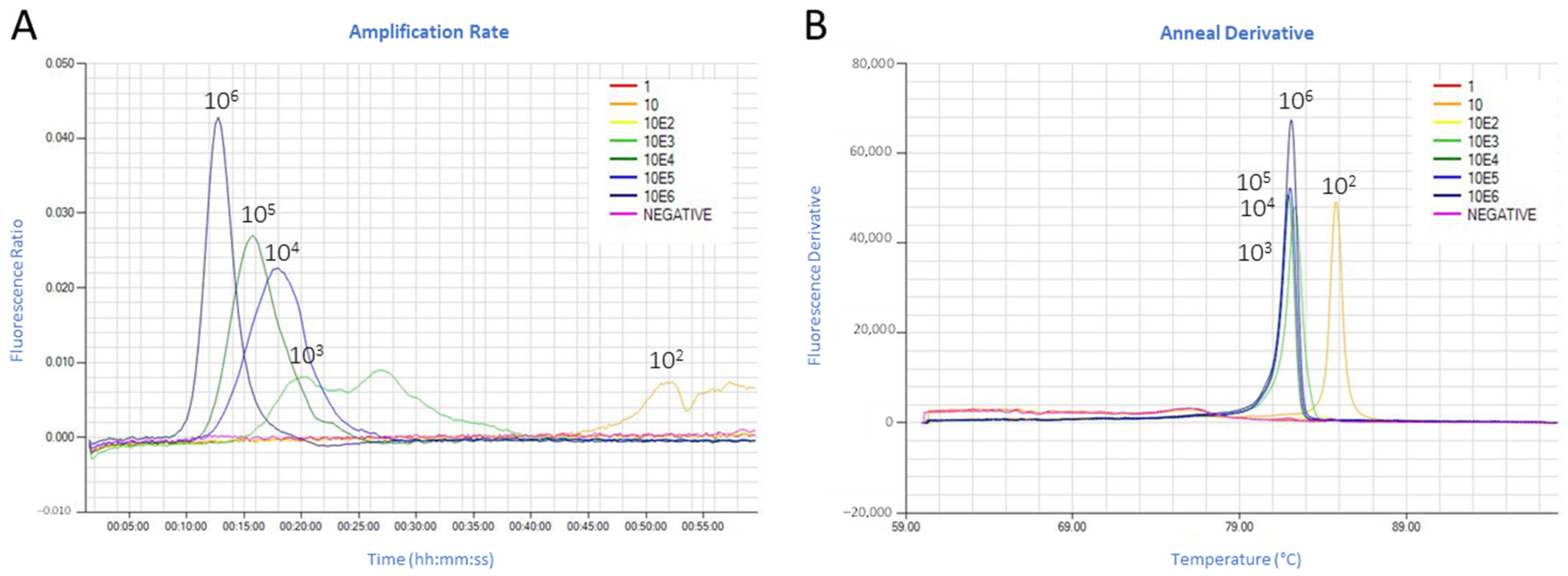
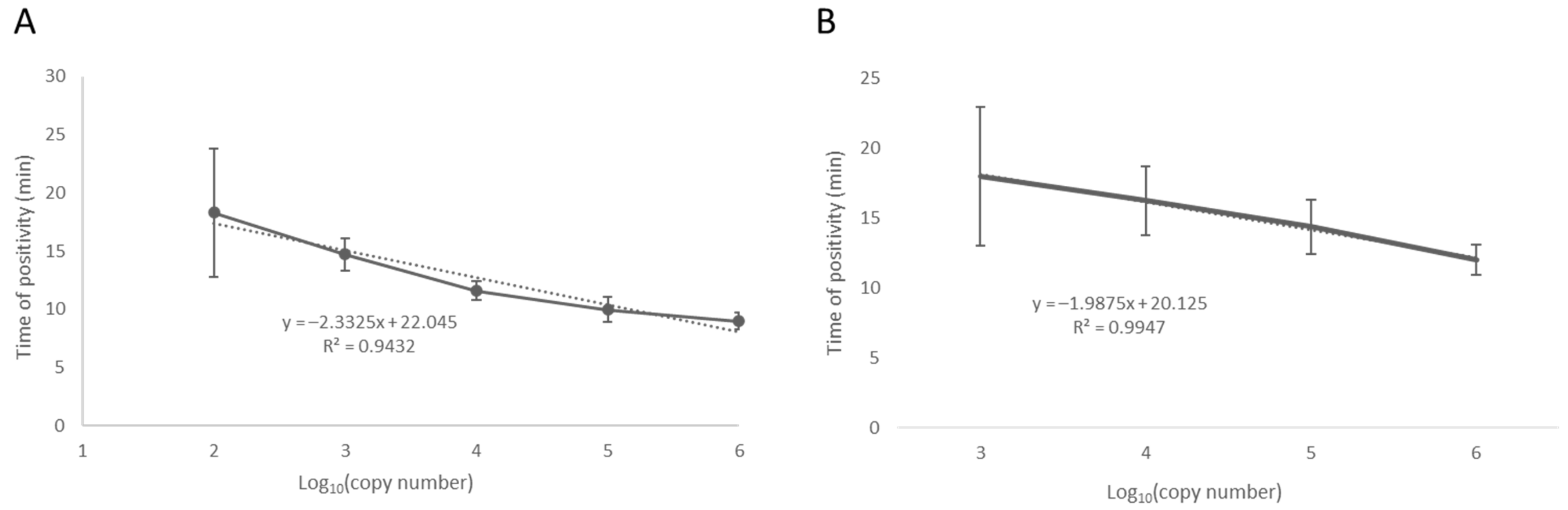
2.3. LAMP Assay Temperature Optimization and Analytical Sensitivity
2.4. Test Specificity
2.5. Pathogen Identification by Standard PCR and Sequencing, and Taqman qPCR
2.6. Testing CyHV-3 LAMP Assay with Archived DNA Samples
2.7. Application of CyHV-3 and CEV LAMP Tests in Non-Lethal Mucus Swabs Samples Taken during Disease Investigations
3. Results
3.1. Fluorescence Real-Time LAMP Test Optimization and Limit of Detection
3.2. Specificity
3.3. CyHV-3 and CEV LAMP Test of Non-Lethal Swabs of Clinically Infected Common Carp
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. FAO Yearbook. Fishery and Aquaculture Statistics 2017/FAO Annuaire; Food and Agriculture Organization of the United Nations: Rome, Italy, 2019. [Google Scholar]
- Su, H.; Su, J. Cyprinid viral diseases and vaccine development. Fish. Shellfish. Immunol. 2018, 83, 84–95. [Google Scholar] [CrossRef]
- Michel, B.; Fournier, G.; Lieffrig, F.; Costes, B.; Vanderplasschen, A. Cyprinid herpesvirus 3. Emerg. Infect. Dis. 2010, 16, 1835–1843. [Google Scholar] [CrossRef] [PubMed]
- Sano, N.; Moriwake, M.; Sano, T. Herpesvirus cyprini: Thermal Effects on Pathogenicity and Oncogenicity. Fish. Pathol. 1993, 28, 171–175. [Google Scholar] [CrossRef]
- Daněk, T.; Kalous, L.; Veselý, T.; Krásová, E.; Reschová, S.; Rylková, K.; Kulich, P.; Petrtýl, M.; Pokorová, D.; Knytl, M. Massive mortality of Prussian carp Carassius gibelio in the upper Elbe basin associated with herpesviral hematopoietic necrosis (CyHV-2). Dis. Aquat. Organ. 2012, 102, 87–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haenen, O.; Way, K.; Gorgoglione, B.; Ito, T.; Paley, R.; Bigarré, L.; Waltzek, T. Novel viral infections threatening Cyprinid fish. In Proceedings of the Bulletin of the European Association of Fish Pathologists, 17th EAFP International Conference, Las Palmas de Gran Canaris, Spain, 7–10 September 2016; Volume 36, pp. 11–23. [Google Scholar]
- Pokorova, D.; Vesely, T.; Piackova, V.; Reschova, S.; Hulova, J. Current knowledge on koi herpesvirus (KHV): A review. Vet. Med. (Praha) 2005, 50, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Gilad, O.; Yun, S.; Adkison, M.A.; Way, K.; Willits, N.H.; Bercovier, H.; Hedrick, R.P. Molecular comparison of isolates of an emerging fish pathogen, koi herpesvirus, and the effect of water temperature on mortality of experimentally infected koi. J. Gen. Virol. 2003, 84, 2661–2668. [Google Scholar] [CrossRef]
- Miwa, S.; Kiryu, I.; Yuasa, K.; Ito, T.; Kaneko, T. Pathogenesis of acute and chronic diseases caused by cyprinid herpesvirus-3. J. Fish. Dis. 2015, 38, 695–712. [Google Scholar] [CrossRef]
- Gotesman, M.; Kattlun, J.; Bergmann, S.M.; El-Matbouli, M. CyHV-3: The third cyprinid herpesvirus. Dis. Aquat. Organ. 2013, 105, 163–174. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.R.; Bently, J.; Beck, L.; Reed, A.; Miller-Morgan, T.; Heidel, J.R.; Kent, M.L.; Rockey, D.D.; Jin, L. Analysis of koi herpesvirus latency in wild common carp and ornamental koi in Oregon, USA. J. Virol. Methods 2013, 187, 372–379. [Google Scholar] [CrossRef]
- Eide, K.E.; Miller-Morgan, T.; Heidel, J.R.; Kent, M.L.; Bildfell, R.J.; LaPatra, S.; Watson, G.; Jin, L. Investigation of Koi Herpesvirus Latency in Koi. J. Virol. 2011, 85, 4954–4962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cano, I.; Mulhearn, B.; Akter, S.; Paley, R. Seroconversion and skin mucosal parameters during koi herpesvirus shedding in common carp, Cyprinus carpio. Int. J. Mol. Sci. 2020, 21, 8482. [Google Scholar] [CrossRef]
- Aoki, T.; Hirono, I.; Kurokawa, K.; Fukuda, H.; Nahary, R.; Eldar, A.; Davison, A.J.; Waltzek, T.B.; Bercovier, H.; Hedrick, R.P. Genome Sequences of Three Koi Herpesvirus Isolates Representing the Expanding Distribution of an Emerging Disease Threatening Koi and Common Carp Worldwide. J. Virol. 2007, 81, 5058–5065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Suárez, N.M.; Wilkie, G.S.; Dong, C.; Bergmann, S.; Lee, P.Y.A.; Davison, A.J.; Vanderplasschen, A.F.C.; Boutier, M. Genomic and biologic comparisons of cyprinid herpesvirus 3 strains. Vet. Res. 2018, 49, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelsma, M.Y.; Way, K.; Dodge, M.J.; Voorbergen-Laarman, M.; Panzarin, V.; Abbadi, M.; El-Matbouli, M.; Skall, H.F.; Kahns, S.; Stone, D.M. Detection of novel strains of cyprinid herpesvirus closely related to koi herpesvirus. Dis. Aquat. Organ. 2013, 107, 113–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swaminathan, T.R.; Kumar, R.; Dharmaratnam, A.; Basheer, V.S.; Sood, N.; Pradhan, P.K.; Sanil, N.K.; Vijayagopal, P.; Jena, J.K. Emergence of carp edema virus in cultured ornamental koi carp, Cyprinus carpio koi, in India. J. Gen. Virol. 2016, 97, 3392–3399. [Google Scholar] [CrossRef] [PubMed]
- Way, K.; Haenen, O.; Stone, D.; Adamek, M.; Bergmann, S.M.; Bigarré, L.; Diserens, N.; El-Matbouli, M.; Gjessing, M.C.; Jung-Schroers, V.; et al. Emergence of carp edema virus (CEV) and its significance to European common carp and koi Cyprinus carpio. Dis. Aquat. Organ. 2017, 126, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Divya, P.; Vertika, B.; Kirty, S.; Jyotirmaya, M.; Pramoda Kumar, S. A review of current understanding on carp edema virus (CEV): A threatful entity in disguise. Int. J. Fish. Aquat. Stud. 2019, 7, 87–93. [Google Scholar]
- Matras, M.; Borzym, E.; Stone, D.; Way, K.; Stachnik, M.; Maj-Paluch, J.; Palusińska, M.; Reichert, M. Carp edema virus in Polish aquaculture—evidence of significant sequence divergence and a new lineage in common carp Cyprinus carpio (L.). J. Fish. Dis. 2017, 40, 319–325. [Google Scholar] [CrossRef]
- Ouyang, P.; Yang, R.; Chen, J.; Wang, K.; Geng, Y.; Lai, W.; Huang, X.; Chen, D.; Fang, J.; Chen, Z.; et al. First detection of carp edema virus in association with cyprinid herpesvirus 3 in cultured ornamental koi, Cyprinus carpio L., in China. Aquaculture 2018, 490, 162–168. [Google Scholar] [CrossRef]
- Toffan, A.; Marsella, A.; Abbadi, M.; Abass, S.; Al-Adhadh, B.; Wood, G.; Stone, D.M. First detection of koi herpesvirus and carp oedema virus in Iraq associated with a mass mortality in common carp (Cyprinus carpio). Transbound. Emerg. Dis. 2020, 67, 523–528. [Google Scholar] [CrossRef]
- Granzow, H.; Fichtner, D.; Schütze, H.; Lenk, M.; Dresenkamp, B.; Nieper, H.; Mettenleiter, T.C. Isolation and partial characterization of a novel virus from different carp species suffering gill necrosis—Ultrastructure and morphogenesis. J. Fish. Dis. 2014, 37, 559–569. [Google Scholar] [CrossRef]
- Ashraf, U.; Lu, Y.; Lin, L.; Yuan, J.; Wang, M.; Liu, X. Spring viraemia of carp virus: Recent advances. J. Gen. Virol. 2016, 97, 1037–1051. [Google Scholar] [CrossRef]
- Taylor, N.G.H.; Peeler, E.J.; Denham, K.L.; Crane, C.N.; Thrush, M.A.; Dixon, P.F.; Stone, D.M.; Way, K.; Oidtmann, B.C. Spring viraemia of carp (SVC) in the UK: The road to freedom. Prev. Vet. Med. 2013, 111, 156–164. [Google Scholar] [CrossRef]
- Taylor, N.G.H.; Dixon, P.F.; Jeffery, K.R.; Peeler, E.J.; Denham, K.L.; Way, K. Koi herpesvirus: Distribution and prospects for control in England and Wales. J. Fish. Dis. 2010, 33, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Bercovier, H.; Fishman, Y.; Nahary, R.; Sinai, S.; Zlotkin, A.; Eyngor, M.; Gilad, O.; Eldar, A.; Hedrick, R.P. Cloning of the koi herpesvirus (KHV) gene encoding thymidine kinase and its use for a highly sensitive PCR based diagnosis. BMC Microbiol. 2005, 5, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilad, O.; Yun, S.; Zagmutt-Vergara, F.J.; Leutenegger, C.M.; Bercovier, H.; Hedrick, R.P. Concentrations of a Koi herpesvirus (KHV) in tissues of experimentally infected Cyprinus carpio koi as assessed by real-time TaqMan PCR. Dis. Aquat. Organ. 2004, 60, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Cano, I.; Stone, D.; Savage, J.; Wood, G.; Mulhearn, B.; Gray, J.; Stinton, N.; Ross, S.; Bonar, M.; Taylor, N.G.H.; et al. Isolation of a Chinook Salmon Bafinivirus (CSBV) in Imported Goldfish Carassius auratus L. In the United Kingdom and evaluation of its virulence in resident fish species. Viruses 2020, 12, 578. [Google Scholar] [CrossRef] [PubMed]
- Barreda-García, S.; Miranda-Castro, R.; de-los-Santos-Álvarez, N.; Miranda-Ordieres, A.J.; Lobo-Castañón, M.J. Helicase-dependent isothermal amplification: A novel tool in the development of molecular-based analytical systems for rapid pathogen detection. Anal. Bioanal. Chem. 2018, 410, 679–693. [Google Scholar] [CrossRef] [PubMed]
- Soliman, H.; El-Matbouli, M. Rapid detection and differentiation of carp oedema virus and cyprinid herpes virus-3 in koi and common carp. J. Fish. Dis. 2018, 41, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Biswas, G.; Sakai, M. Loop-mediated isothermal amplification (LAMP) assays for detection and identification of aquaculture pathogens: Current state and perspectives. Appl. Microbiol. Biotechnol. 2014, 98, 2881–2895. [Google Scholar] [CrossRef]
- Valverde, E.J.; Cano, I.; Castro, D.; Paley, R.K.; Borrego, J.J. Rapid and Sensitive Detection of Lymphocystis Disease Virus Genotype VII by Loop-Mediated Isothermal Amplification. Food Environ. Virol. 2017, 9, 114–122. [Google Scholar] [CrossRef]
- Cano, I.; McCullough, R.; Mulhearn, B.; Gunning, S.; Waine, A.; Joiner, C.; Paley, R. Non-lethal loop-mediated isothermal amplification assay as a point-of-care diagnostics tool for Neoparamoeba perurans, the causative agent of amoebic gill disease. J. Fish. Dis. 2020, 43, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, J.A.; Dickinson, M.; Hobden, E.; Robinson, S.; Giltrap, P.M.; Boonham, N. A five-minute DNA extraction method for expedited detection of Phytophthora ramorum following prescreening using Phytophthora spp. lateral flow devices. J. Microbiol. Methods 2010, 81, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Lau, H.Y.; Botella, J.R. Advanced DNA-Based Point-of-Care Diagnostic Methods for Plant Diseases Detection. Front. Plant. Sci. 2017, 8, 2016. [Google Scholar] [CrossRef]
- Ali, N.; Rampazzo, R.d.C.P.; Costa, A.D.T.; Krieger, M.A. Current Nucleic Acid Extraction Methods and Their Implications to Point-of-Care Diagnostics. Biomed. Res. Int. 2017, 2017, 1–13. [Google Scholar]
- Gunimaladevi, I.; Kono, T.; Venugopal, M.N.; Sakai, M. Detection of koi herpesvirus in common carp, Cyprinus carpio L., by loop-mediated isothermal amplification. J. Fish. Dis. 2004, 27, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Saleh, M.; El-Matbouli, M. Rapid detection of Cyprinid herpesvirus-3 (CyHV-3) using a gold nanoparticle-based hybridization assay. J. Virol. Methods 2015, 217, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Prescott, M.A.; Reed, A.N.; Jin, L.; Pastey, M.K. Rapid Detection of Cyprinid Herpesvirus 3 in Latently Infected Koi by Recombinase Polymerase Amplification. J. Aquat. Anim. Health 2016, 28, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, M.; Watari, H.; Kojima, T.; Ikedo, M. Sensitive and rapid detection of koi herpesvirus by LAMP method. Fish. Pathol. 2006, 41, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Yoshino, M.; Watari, H.; Kojima, T.; Ikedo, M.; Kurita, J. Rapid, sensitive and simple detection method for koi herpesvirus using loop-mediated isothermal amplification. Microbiol. Immunol. 2009, 53, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Neukirch, M.; Böttcher, K.; Bunnajirakul, S. Isolation of a virus from koi with altered gills. Bull. Eur. Assoc. Fish. Pathol. 1999, 19, 221–224. [Google Scholar]
- Gilad, O.; Yun, S.; Andree, K.B.; Adkison, M.A.; Zlotkin, A.; Bercovier, H.; Eldar, A.; Hedrick, R.P. Initial characteristics of koi herpesvirus and development of a polymerase chain reaction assay to detect the virus in koi, Cyprinus carpio koi. Dis. Aquat. Organ. 2002, 48, 101–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vancsok, C.; Peñaranda, M.M.D.; Raj, V.S.; Leroy, B.; Jazowiecka-Rakus, J.; Boutier, M.; Gao, Y.; Wilkie, G.S.; Suárez, N.M.; Wattiez, R.; et al. Proteomic and Functional Analyses of the Virion Transmembrane Proteome of Cyprinid Herpesvirus 3. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [Green Version]
- Costes, B.; Fournier, G.; Michel, B.; Delforge, C.; Raj, V.S.; Dewals, B.; Gillet, L.; Drion, P.; Body, A.; Schynts, F.; et al. Cloning of the Koi Herpesvirus Genome as an Infectious Bacterial Artificial Chromosome Demonstrates That Disruption of the Thymidine Kinase Locus Induces Partial Attenuation in Cyprinus carpio koi. J. Virol. 2008, 82, 4955–4964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klafack, S.; Wang, Q.; Zeng, W.; Wang, Y.; Li, Y.; Zheng, S.; Kempter, J.; Lee, P.Y.; Matras, M.; Bergmann, S.M. Genetic variability of koi herpesvirus in vitro-a natural event? Front. Microbiol. 2017, 8, 982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Lee, X.; Weng, S.; He, J.; Dong, C. Whole-genome sequence of a novel Chinese cyprinid herpesvirus 3 isolate reveals the existence of a distinct European genotype in East Asia. Vet. Microbiol. 2015, 175, 185–194. [Google Scholar] [CrossRef]
- Van Beurden, S.J.; Bossers, A.; Voorbergen-Laarman, M.H.A.; Haenen, O.L.M.; Peters, S.; Abma-Henkens, M.H.C.; Peeters, B.P.H.; Rottier, P.J.M.; Engelsma, M.Y. Complete genome sequence and taxonomic position of anguillid herpesvirus 1. J. Gen. Virol. 2010, 91, 880–887. [Google Scholar] [CrossRef] [PubMed]
- Davison, A.J.; Kurobe, T.; Gatherer, D.; Cunningham, C.; Korf, I.; Fukuda, H.; Hedrick, R.P.; Waltzek, T.B. Comparative Genomics of Carp Herpesviruses. J. Virol. 2013, 87, 2908–2922. [Google Scholar] [CrossRef] [Green Version]
- Gjessing, M.C.; Yutin, N.; Tengs, T.; Senkevich, T.; Koonin, E.; Rønning, H.P.; Alarcon, M.; Ylving, S.; Lie, K.-I.; Saure, B.; et al. Salmon Gill Poxvirus, the Deepest Representative of the Chordopoxvirinae. J. Virol. 2015, 89, 9348–9367. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Howson, E.L.A.; Soldan, A.; Webster, K.; Beer, M.; Zientara, S.; Belák, S.; Sánchez-Vizcafno, J.M.; Van Borm, S.; King, D.P.; Fowler, V.L. Technological advances in veterinary diagnostics: Opportunities to deploy rapid decentralised tests to detect pathogens affecting livestock. OIE Rev. Sci. Tech. 2017, 36, 479–498. [Google Scholar] [CrossRef] [PubMed]
- OIE. Manual of Diagnostic Tests for Aquatic Animals; Office international des épizooties: New York, NY, USA, 2003; ISBN 9290445637. [Google Scholar]
- Ryan, E.; Gloster, J.; Reid, S.M.; Li, Y.; Ferris, N.P.; Waters, R.; Juleff, N.; Charleston, B.; Bankowski, B.; Gubbins, S.; et al. Clinical and laboratory investigations of the outbreaks of foot-and-mouth disease in Southern England in 2007. Vet. Rec. 2008, 163, 139–147. [Google Scholar] [CrossRef] [PubMed]
- OIE. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; Office international des épizooties: New York, NY, USA, 2013. [Google Scholar]
- Adams, A.; Thompson, K.D. Development of diagnostics for aquaculture: Challenges and opportunities. Aquac. Res. 2011, 42, 93–102. [Google Scholar] [CrossRef]
- Lovy, J.; Friend, S.E.; Al-Hussinee, L.; Waltzek, T.B. First report of carp edema virus in the mortality of wild common carp Cyprinus carpio in North America. Dis. Aquat. Organ. 2018, 131, 177–186. [Google Scholar] [CrossRef]
- Reed, A.N.; Izume, S.; Dolan, B.P.; LaPatra, S.; Kent, M.; Dong, J.; Jin, L. Identification of B Cells as a Major Site for Cyprinid Herpesvirus 3 Latency. J. Virol. 2014, 88, 9297–9309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, B.K.; Jacobsen, M.W.; Middelboe, A.L.; Preston, C.M.; Marin, R.; Bekkevold, D.; Knudsen, S.W.; Møller, P.R.; Nielsen, E.E. Remote, autonomous real-time monitoring of environmental DNA from commercial fish. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Sunarto, A.; McColl, K.A.; Crane, M.S.J.; Schat, K.A.; Slobedman, B.; Barnes, A.C.; Walker, P.J. Characteristics of cyprinid herpesvirus 3 in different phases of infection: Implications for disease transmission and control. Virus Res. 2014, 188, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Shephard, K.L. Functions for fish mucus. Rev. Fish. Biol. Fish. 1994, 4, 401–429. [Google Scholar] [CrossRef]
- Kim, S.W.; Giri, S.S.; Kim, S.G.; Kwon, J.; Oh, W.T.; Park, S.C. Carp edema virus and cyprinid herpesvirus-3 coinfection is associated with mass mortality of koi (Cyprinus carpio haematopterus) in the Republic of Korea. Pathogens 2020, 9, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Primer | Sequence (5′-3′) | °C | Position |
|---|---|---|---|---|
| CyH-3 orf43 | F3 | CTGATGGTCCTGTCCGAG | 64 | 72,147 |
| B3 | ACTGGTTAATGCAGTCGTC | 72,357 | ||
| FIP (F1c + F2) | CCGTGTTGAGGTAGAAGAGGCCCAGACCTACGTGAAAGC | - | ||
| BIP (B1c + B2) | CTGGACCTTCCGTCTGCTGGTCTTGTTGCGGAAGGTG | - | ||
| Loop-F | ACATGAGGCTGTTGTAGGTG | 72,240 | ||
| Loop-B | GTACGAGTGCATCGCCTA | 72,296 | ||
| CEV p4a | F3 | CCAGATTTGGGTTGAAACATG | 64 | 139 |
| B3 | GCAAAGTACTACCTCATCCAA | 494 | ||
| FIP (F1c + F2) | CTTGCTCTAGTTCTAGGATTGTACTGTAACTCCTTGAGGAAT | - | ||
| BIP (B1c + B2) | GTCTTGTCAAAGACAGACATCTGTTGACACAATTCCAGAACAAG | - | ||
| Loop-F | ATTGCGGAATTCTAGATCAG | 260 | ||
| Loop-B | GGATTCCTTTCCAGAACATAAC | 387 | ||
| Carp ef1a | F3 | TGCCACTTCAGGATGTCTA | 64 | 800 |
| B3 | GGATGTCCTTGACAGACAC | 1200 | ||
| FIP (F1c + F2) | GTCACAACCATACCAGGCTTGAATTGGAGGTATTGGAACTGTG | - | ||
| BIP (B1c + B2) | GCCAACTTGACCACTGAGGTGTAGCCTCAGCAAGAGATTC | - | ||
| Loop-F | GACACCAGTCTCCACACG | 870 | ||
| Loop-B | AAGTCTGTTGAGATGCACCAT | 965 |
| Pathogen | № | LAMP Amplification | MT | LAMP Interpretation | |
|---|---|---|---|---|---|
| False-Positive | False-Negative | ||||
| CyHV-3 | 85 | 76 | Correct | NA | 9 (˂LOD) |
| CyHV-3 variant | 30 | 0 | NA | 0 | NA |
| CyHV-1 | 28 | 3 | Different in 2 out of 3 | 1 | NA |
| CyHV-2 | 10 | 3 | Same as CyHV3, coinfection confirmed in one sample | 2 | NA |
| AngHV | 2 | 1 | Different | 0 | NA |
| HV | 2 | 1 | Same as CyHV-3 | 1 | NA |
| Negative samples | 47 | 0 | NA | 0 | NA |
| Site | № | LAMP POCT (%) | Site Designation | |||||
|---|---|---|---|---|---|---|---|---|
| ef1a | CyHV-3 | CEV | LAMP POCT | Statutory Diagnostics | ||||
| CyHV-3 | CEV | CyHV-3 | CEV | |||||
| A | 5 | 80 | 100 | 0 | + | - | + | - |
| B | 1 | 100 | 100 | 0 | + | - | + | - |
| C | 5 | 100 | 100 | 40 | + | + | + | + |
| D | 1 | 100 | 100 | 0 | + | - | + | - |
| E | 5 | 100 | 80 | 0 | + | - | + | - |
| F | 1 | 100 | 100 | 0 | + | - | + | - |
| G | 5 | 0 | 20 | 0 | + | - | + | - |
| H | 1 | 100 | 0 | 0 | - | - | - | - |
| I | 4 | 100 | 100 | 0 | + | - | + | - |
| J | 2 | 100 | 100 | 0 | + | - | + | - |
| K | 10 | 40 | 70 | 0 | + | - | + | - |
| L | 10 | 20 | 100 | 0 | + | - | + | - |
| M | 2 | 50 | 0 | 0 | - | - | - | - |
| N | 3 | 100 | 66 | 0 | + | - | + | - |
| O | 3 | 100 | 0 | 0 | - | - | - | - |
| P | 5 | 40 | 100 | 0 | + | - | + | - |
| Assay | Test Visualization | Time | Target | LOD | Specificity | Sample Type | Internal Control | Reference |
|---|---|---|---|---|---|---|---|---|
| LAMP | Gel electrophoresis | ˃60 | CyHV-3 tk | 10−6 stock dilution | Only in silico tested | Homogenates | None | [38] |
| LAMP | Turbidity | 60 | CyHV-3 | 10−7 stock dilution; 6 copies | No cross-reaction with fish HVs (n = 3) | Homogenates | None | [41,42] |
| RPA | LFD | ˂30 | CyHV-3 mcp | 10 copies | No cross-reaction with CyHV-2 (n = 1) | White blood cells | None | [40] |
| Multiplex RPA | LFD | 25 | CyHV-3 tk CEV p4a | CyHV-3: 21 copies CEV: 1.8 copies | No cross-reaction with CyHV-1 or CyHV-2 (n = 2) | Homogenates | None | [31] |
| AuNPs hybridization | Colorimetric | 21 | CyHV-3 tk | 150 copies | No cross-reaction with CyHV-1 or CyHV-2 (n = 2) | Homogenates | None | [39] |
| LAMP | Fluorescence real-time | ˂20 | CyHV-3 orf43 | 100 copies | 4.4% cross-reaction with other fish HVs (n = 72) | Homogenates Gill/skin swabs | Host ef1a | This study |
| LAMP | Fluorescence real-time | ˂20 | CEV p4a | 1000 copies | No cross-reaction with fish HVs (n = 3) | Homogenates Gill/skin swabs | Host ef1a | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cano, I.; Worswick, J.; Mulhearn, B.; Stone, D.; Wood, G.; Savage, J.; Paley, R. A Seasonal Study of Koi Herpesvirus and Koi Sleepy Disease Outbreaks in the United Kingdom in 2018 Using a Pond-Side Test. Animals 2021, 11, 459. https://doi.org/10.3390/ani11020459
Cano I, Worswick J, Mulhearn B, Stone D, Wood G, Savage J, Paley R. A Seasonal Study of Koi Herpesvirus and Koi Sleepy Disease Outbreaks in the United Kingdom in 2018 Using a Pond-Side Test. Animals. 2021; 11(2):459. https://doi.org/10.3390/ani11020459
Chicago/Turabian StyleCano, Irene, John Worswick, Brian Mulhearn, David Stone, Gareth Wood, Jacqueline Savage, and Richard Paley. 2021. "A Seasonal Study of Koi Herpesvirus and Koi Sleepy Disease Outbreaks in the United Kingdom in 2018 Using a Pond-Side Test" Animals 11, no. 2: 459. https://doi.org/10.3390/ani11020459