
Carcass and Pork Quality and Gut Environment of Pigs Fed a Diet Supplemented with the Bokashi Probiotic


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Feed
2.2. Carcass and Meat Quality
2.3. Shear Force
2.4. Proximate Analysis
2.5. Mineral Composition
2.6. Microbiological Determinations
Identification of LAB and Yeasts
2.7. Statistical Analysis
3. Results
3.1. Microbiological Analysis
3.2. Carcass and Meat Quality
3.3. Mineral Concentration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Gaggía, F.; Mattarelli, P.; Biavati, B. Probiotics and prebiotics in animal feeding for safe food production. Int. J. Food Microbiol. 2010, 141, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Rai, V.; Yadav, B.; Lakhani, G.P. Application of probiotic and prebiotic in animals production: A review. Environ. Ecol. 2013, 31, 873–876. [Google Scholar]
- Shon, S.K.; Hong, J.W.; Min, B.J.; Lee, W.B.; Kim, I.H. Effects of Lactobacillus reuteri-based direct-fed microbial supplementation for growing-finishing pigs. Asian-Australas. J. Anim. Sci. 2005, 18, 370–374. [Google Scholar] [CrossRef]
- Lee, S.J.; Lee, S.S.; Shim, N.H.; Ok, J.U.; Jung, H.S.; Chu, G.M.; Kim, I.H.; Kim, J.D. Effects of dietary synbiotics from anaerobic microflora on growth performance, noxious gas emission and fecal pathogenic bacteria population in weaning pigs. Asian-Australas. J. Anim. Sci. 2009, 22, 1202–1208. [Google Scholar] [CrossRef]
- Dvoroznáková, E.; Bucková, B.; Hurníková, Z.; Revajová, V.; Lauková, A. Effect of probiotic bacteria on phagocytosis and respiratory burst activity of blood polymorphonuclear leukocytes (PMNL) in mice infected with Trichinella spiralis. Vet. Parasitol. 2016, 231, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Dhama, K.; Verma, V.; Sawant, P.M.; Tiwari, R.; Vaid, R.K.; Chauhan, R.S. Applications of probiotics in poultry: Enhancing immunity and beneficial effects on production performances and health. Vet. Immunol. Immunopathol. 2011, 13, 1–19. [Google Scholar]
- Koenen, M.E.; Kramer, J.; Van Der Hulst, R.; Heres, L.; Jeurissen, S.H.; Boersma, W.J. Immunomodulation by probiotic lactobacilli in layer and meat-type chickens. Br. Poult. Sci. 2004, 45, 355–366. [Google Scholar] [CrossRef]
- Nousiainen, J.; Setälä, J. Lactic acid bacteria as animal probiotics. In Lactic Acid Bacteria; Salminen, S., Von Wright, A., Eds.; Marcel Dekker: New York, NY, USA, 1993; pp. 315–356. [Google Scholar]
- Meng, Q.W.; Yan, L.; Ao, X.; Zhou, T.X.; Wang, J.P.; Lee, J.H.; Kim, I.H. Influence of probiotics in different energy and nutrient density diets on growth performance, nutrient digestibility, meat quality, and blood characteristics in growing-finishing pigs. J. Anim. Sci. 2010, 88, 3320–3326. [Google Scholar] [CrossRef] [Green Version]
- Modesto, M.; D’Aimmo, M.R.; Stefanini, I.; Trevisi, P.; De Filippi, S.; Casini, L.; Mazzoni, M.; Bosi, P.; Biavati, B. A novel strategy to select Bifidobacterium strains and prebiotics as natural growth promoters in newly weaned pigs. Livest. Sci. 2009, 122, 248–258. [Google Scholar] [CrossRef]
- Napiórkowska, B.; Dobrowolska, Z.; Więcek, J.; Gajewska, J.; Rekiel, A. Effect of a probiotic preparation on daily weight gain, survival rate and composition of faecal microflora in piglets. Rocz. Naukowe PTZ 2014, 10, 57–68. [Google Scholar]
- Dowarah, R.; Verma, A.K.; Agarwal, N. The use of Lactobacillus as an alternative of antibiotic growth promoters in pigs: A review. Anim. Nutr. 2017, 3, 1–6. [Google Scholar] [CrossRef]
- Alexopoulos, C.; Georgoulakis, I.E.; Tzivara, A.; Kyriakis, S.C.; Govaris, A. Field evaluation of the efficacy of a probiotic-containing Bacillus licheniformis and Bacillus subtilis spores, on the health status, performance and carcass quality of grower and finishing pigs. J. Vet. Med. Ser. A. Physiol. Pathol. Clin. Med. 2004, 51, 306–312. [Google Scholar] [CrossRef]
- Suo, C.; Yin, Y.; Wang, X.; Lou, X.; Song, D.; Wang, X.; Gu, Q. Effects of Lactobacillus plantarum ZJ316 on pig growth and pork quality. BMC Vet. Res. 2012, 8, 89. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Su, B.; Wang, J.; Zhang, C.; Shan, A. Effects of probiotics on growth, pork quality and serum metabolites in growing-finishing pigs. J. Northeast Agric. Univ. 2013, 20, 57–63. [Google Scholar] [CrossRef]
- Laskowska, E.; Jarosz, Ł.; Grądzki, Z. Effect of multi-microbial probiotic formulation Bokashi on pro- and anti-inflammatory cytokines profile in the serum, colostrum and milk of sows, and in a culture of polymorphonuclear cells isolated from colostrum. Probiotics Antimicrob. Proteins 2019, 11, 220–232. [Google Scholar] [CrossRef] [Green Version]
- Reszka, P.; Dunisławska, A.; Sławińska, A.; Siwek, M.; Kapelański, W.; Bogucka, J. Influence of the effective microorganisms (EM) on performance, intestinal morphology and gene expression in the jejunal mucosa of pigs fed different diets. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1444–1453. [Google Scholar] [CrossRef]
- Rybarczyk, A.; Bogusławska-Wąs, E.; Łupkowska, A. Effect of EM® probiotic on gut microbiota, growth performance, carcass and meat quality of pigs. Livest. Sci. 2020, 241, 104206. [Google Scholar] [CrossRef]
- Prange, H.; Juggrt, L.; Scharner, E. Untersuchungen zur Muskel Fleischqualität beim Schwein. Arch. Exp. Vet. Med. 1977, 31, 235–248. [Google Scholar]
- Naveau, J.; Pommeret, P.; Lechaux, P. Proposition d’une méthode de mesuredu rendement technologique: La méthode Napole. Techni-Porc. 1985, 8, 7–13. [Google Scholar]
- Koćwin-Podsiadła, M.; Kurył, J.; Krzęcio, E.; Antosik, K.; Zybert, A.; Sieczkowska, H. An association between genotype at the CAST (calpastatin) locus and carcass quality traits in porkers free of RYR1T allele. Anim. Sci. Pap. Rep. 2004, 22, 497–505. [Google Scholar]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 20th ed.; AOAC: Rockville, MD, USA, 2016. [Google Scholar]
- ISO 4833-1:2013. Microbiology of the Food Chain—Horizontal Method for the Enumeration of Microorganisms. Part 2: Colony Count at 30 °C by the Surface Plating Technique. Available online: https://www.iso.org/obp/ui/#iso:std:iso:4833:-1:ed-1:v1:en (accessed on 27 May 2021).
- ISO 21527-1:2008. Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Yeasts and Moulds. Part 1: Colony Count Technique in Products with Water Activity Greater than 0.95. Available online: https://www.iso.org/obp/ui/#iso:std:iso:21527:-1:ed-1:v1:en (accessed on 27 May 2021).
- ISO 21528-1:2017. Microbiology of the Food Chain—Horizontal Method for the Detection and Enumeration of Enterobacteriaceae—Part 1: Detection of Enterobacteriaceae. Available online: https://www.iso.org/obp/ui/#iso:std:iso:21528:-1:ed-2:v1:en (accessed on 27 May 2021).
- ISO 15214:1998. Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Mesophilic Lactic Acid Bacteria. Available online: https://www.iso.org/obp/ui/#iso:std:iso:15214:ed-1:v1:en (accessed on 27 May 2021).
- ISO 15213:2003. Microbiology of Food and Animal Feeding Stuffs—Horizontal Method for the Enumeration of Sulfite-Reducing Bacteria Growing under Anaerobic Conditions. Available online: https://www.iso.org/obp/ui/#iso:std:iso:15213:ed-1:v1:en (accessed on 27 May 2021).
- ISO 6579-1:2017. Microbiology of the Food Chain—Horizontal Method for the Detection, Enumeration and Serotyping of Salmonella—Part 1: Detection of Salmonella spp. Available online: https://www.iso.org/obp/ui/#iso:std:iso:6579:-1:ed-1:v1:en (accessed on 27 May 2021).
- ISO 11290-1:2017. Microbiology of the Food Chain—Horizontal Method for the Detection and Enumeration of Listeria Monocytogenes and of Listeria spp.—Part 1: Detection Method. Available online: https://www.iso.org/obp/ui/#iso:std:iso:11290:-1:ed-2:v1:en (accessed on 27 May 2021).
- ISO 7218:2007. Microbiology of Food and Animal Feeding Stuffs—General Requirements and Guidance for Microbiological Examinations. Available online: https://www.iso.org/obp/ui/#iso:std:iso:7218:ed-3:v1:amd:1:v2:en (accessed on 27 May 2021).
- Tajabadi, N.; Mardan, M.; Abdul Manap, M.; Shuhaimi, M.; Meimandipour, A.; Nateghi, L. Detection and identification of Lactobacillus bacteria found in the honey stomach of the giant honeybee Apis dorsata. Apidologie 2011, 42, 642–649. [Google Scholar] [CrossRef]
- Dongyou, L. Molecular Detection of Foodborne Pathogens; CRC Press: Boca Raton, FL, USA, 2009; p. 905. ISBN 1420076442/9781420076448. [Google Scholar]
- Rybarczyk, A.; Bogusławska-Wąs, E.; Dłubała, A. Effect of BioPlus YC probiotic supplementation on gut microbiota, production performance, carcass and meat quality of pigs. Animals 2021, 11, 1581. [Google Scholar] [CrossRef]
- Bajagai, Y.S.; Klieve, A.V.; Dart, P.J.; Bryden, W.J. Probiotics in Animal Nutrition: Production, Impact and Regulation; Food and Agriculture Organization of the United Nations, FAO: Rome, Italy, 2016. [Google Scholar]
- Giang, H.H.; Viet, T.Q.; Ogle, B.; Lindberg, J.E. Growth performance, digestiolilty gut enviroment and health status in weaned piglets fed a diet supplemented with potentially probiotic complexes of lactic acid bacteria. Livest. Sci. 2010, 129, 95–103. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, J.Q.; Liu, H.L.; Lai, T.; Ma, J.L.; Wang, J.F.; Zhu, Y.H. Evaluation of Lactobacillus rhamnosus GG using an Escherichia coli K88 model of piglet diarrhoea: Effects on diarrhoea incidence, faecal microflora and immune responses. Vet. Microbiol. 2010, 141, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.H.; Qiao, S.Y.; Li, D.F.; Piao, X.S.; Ren, J.P. Effects of Lactobacilli on the performance, diarrhoea incidence, VFA concentration and gastrointestinal microbial flora of weaning pigs. Asian Australas. J. Anim. Sci. 2004, 17, 401–409. [Google Scholar] [CrossRef]
- Chiang, M.L.; Chen, H.C.; Chen, K.N.; Lin, Y.C.; Lin, Y.T.; Chen, M.J. Optimizing production of two potential probiotic Lactobacilli strains isolated from piglet feces as feed additives for weaned piglets. Asian Australas. J. Anim. Sci. 2015, 28, 1163–1170. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.Y.; Kim, Y.H.; Rhee, M.H.; Song, J.C.; Lee, K.W.; Kim, K.S.; Lee, S.P.; Lee, I.S.; Park, S.C. Selection of Lactobacillus sp. PSC101 that produces active dietary enzymes such as amylase, lipase, phytase and protease in pigs. J. Gen. Appl. Microbiol. 2007, 53, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Barowicz, T.; Migdał, W.; Pieszka, M.; Živkovič, B. The effect of linseed PUFA n-3 and probiotics on fatty acid composition and cholesterol level in longissimus dorsi muscle of fatteners. Biotechnol. Anim. Husb. 2003, 19, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Jukna, C.; Jukna, V.; Šimkus, A. The effect of probiotics and phytobiotics on meat properties and quality in pigs. Vet. Med. Zoot. 2005, 29, 80–84. [Google Scholar]
- He, M.; Fang, S.; Huang, X.; Zhao, Y.; Ke, S.; Yang, H.; Huang, L. Evaluating the contribution of gut microbiota to the variation of porcine fatness with the cecum and fecal samples. Front. Microbiol. 2016, 7, 2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, C.; Yan, W.; Sun, C.; Ji, C.; Zhou, Q.; Zhang, D.; Yang, N. The gut microbiota is largely independent of host genetics in regulating fat deposition in chickens. ISME J. 2019, 13, 1422–1436. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Cui, Y.; Lu, H.; Wang, G.; Ma, X. Effect of long-term dietary probiotic Lactobacillus reuteri or antibiotics on meat quality, muscular amino acids and fatty acids in pigs. Meat Sci. 2021, 171, 108234. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.Y.; Belal, S.A.; Kong, D.R.; Choi, Y.I.; Kim, Y.H.; Choe, H.S.; Heo, J.Y.; Shim, K.S. Influence of probiotics-friendly pig production on meat quality and physicochemical characteristics. Korean J. Food Sci. Anim. Resour. 2018, 38, 403–416. [Google Scholar] [CrossRef] [PubMed]
- Khanal, P.; Maltecca, C.; Schwab, C.; Fix, J.; Tiezzi, F. Microbiability of meat quality and carcass composition traits in swine. J. Anim. Breed. Genet. 2021, 138, 223–236. [Google Scholar] [CrossRef]
- Li, L.; Chen, C.X. Effects of probiotics on meat quality of broilers and mechanism. Heilongjiang Anim. Sci. Vet. Med. 2009, 11, 43–44. [Google Scholar]
- Jiang, J. Effect of ASTA on weight gain and meat quality on finishing pigs. Hunan Feed 2011, 5, 40–43. [Google Scholar]
- Nam, Y.J.; Choi, Y.M.; Lee, S.H.; Choe, J.H.; Jeong, D.W.; Kim, Y.Y.; Kim, B.C. Sensory evaluations of porcine longissimus dorsi muscle: Relationships with postmortem meat quality traits and muscle fiber characteristics. Meat Sci. 2009, 83, 731–736. [Google Scholar] [CrossRef]
- Joo, S.T.; Kim, G.D.; Hwang, Y.H.; Ryu, Y.C. Control of fresh meat quality through manipulation of muscle fiber characteristics. Meat Sci. 2013, 95, 828–836. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, T.L.; Koohmaraie, M.; Shackelford, S.D. Effect of vitamin C concentration and co-injection with calcium chloride on beef retail display color. J. Anim. Sci. 1996, 74, 1846–1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Wu, S.; He, J.; He, C.; Wang, P.; Zeng, Q.; Yang, F. Salt-induced osmotic stress stimulates selenium biotransformation in Lactobacillus rhamnosus ATCC 53103. Food Sci. Technol. 2020, 131, 109763. [Google Scholar] [CrossRef]
- Skrypnik, K.; Suliburska, J. Association between the gut microbiota and mineral metabolism. J. Sci. Food Agric. 2018, 98, 2449–2460. [Google Scholar] [CrossRef] [PubMed]
- De Vries, A.G.; Van Der Wal, P.G.; Long, T.; Eikelenboom, G.; Merks, J.W.M. Genetic parameters of pork quality and production traits in yorkshire populations. Livest. Prod. Sci. 1994, 40, 277–289. [Google Scholar] [CrossRef]
- Jacyno, E.; Pietruszka, A.; Kołodziej, A. Influence of pig meatiness on pork meat quality. Pol. J. Food Nutr. Sci. 2006, 15, 137–140. [Google Scholar]
- Huff-Lonergan, E.; Baas, T.J.; Malek, M.; Dekkers, J.C.M.; Prusa, K.; Rothschild, M.F. Correlations among selected pork quality traits. J. Anim. Sci. 2002, 80, 617–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miar, Y.; Plastow, G.S.; Moore, S.S.; Manafiazar, G.; Charagu, P.; Kemp, R.A.; Van Haandel, B.; Huisman, A.E.; Zhang, C.Y.; McKay, R.M.; et al. Genetic and phenotypic parameters for carcass and meat quality traits in commercial crossbred pigs. J. Anim. Sci. 2014, 92, 2869–2884. [Google Scholar] [CrossRef] [Green Version]


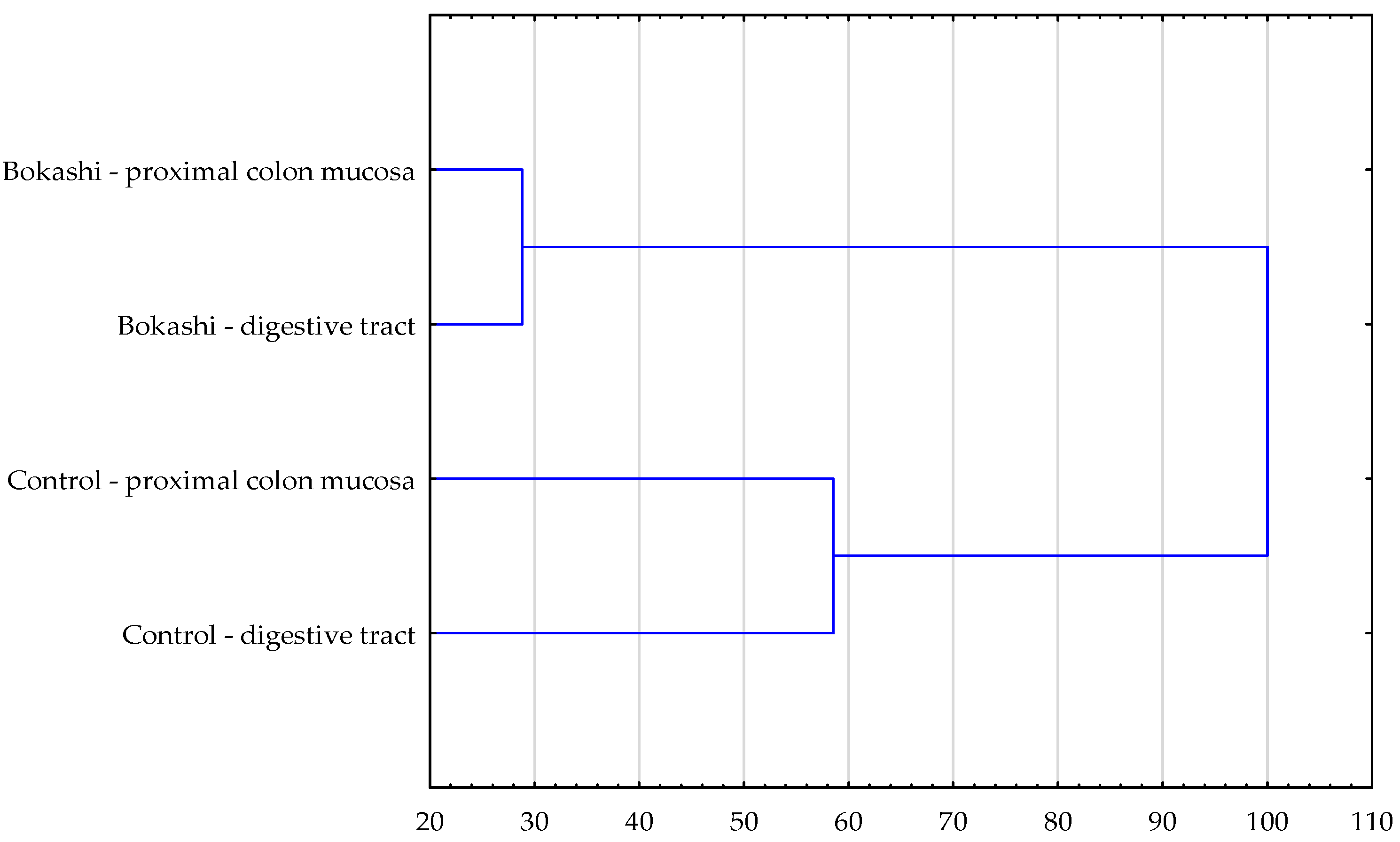{kind=link}
| Microbiota Composition (log10/g) | Control | Feed + Bokashi Mixture | Bokashi (Preparation) | p-Value | |
|---|---|---|---|---|---|
| LAB | L. plantarum | 0.95 c ± 0.18 | 1.92 b ± 0.06 | 2.81 a ± 0.35 | 0.001 |
| L. casei | 1.54 c ± 0.07 | 3.73 b ± 0.29 | 4.68 a ± 0.35 | 0.005 | |
| Total | 1.90 c ± 0.18 | 3.92 b ± 0.06 | 4.96 a ± 0.35 | 0.003 | |
| TYMC | S. cerevisiae | 0.25 c ± 0.33 | 3.98 b ± 0.25 | 6.88 a ± 0.08 | 0.009 |
| Candida spp. | < 1.0 | <1.0 | 1.0 ± 0.05 | - | |
| Total | 0.25 c ± 0.33 | 3.98 b ± 0.25 | 6.88 a ± 3.14 | 0.003 | |
| TBC | - | 3.28 a ± 0.09 | 2.00 ab ± 0.41 | 1.41 b ± 0.29 | 0.002 |
| Microbiological Fractions (log10/g) | Control | Bokashi | p-Value |
|---|---|---|---|
| Proximal colon mucosa | |||
| TBC | 7.90 ± 0.43 | 7.79 ± 0.40 | 0.706 |
| LAB | 6.04 b ± 0.45 | 7.21 a ± 1.06 | 0.036 |
| TYMC | 2.15 b ± 0.06 | 2.85 a ± 0.20 | 0.026 |
| TCE | 7.24 ± 1.21 | 6.35 ± 0.33 | 0.195 |
| CL | 2.77 a ± 0.01 | 2.27 b ± 0.28 | 0.009 |
| Digestive tract of proximal colon | |||
| TBC | 7.08 ± 0.02 | 7.48 ± 0.90 | 0.414 |
| LAB | 8.28 ± 0.44 | 8.18 ± 0.24 | 0.633 |
| TYMC | 1.79 b ± 0.01 | 2.24 a ± 0.28 | 0.016 |
| TCE | 6.36 a ± 0.05 | 4.61 b ± 0.56 | 0.001 |
| CL | 2.44 a ± 0.47 | 1.58 b ± 0.01 | 0.028 |
| Traits | Control | Bokashi | p-Value |
|---|---|---|---|
| HCW (kg) | 85.30 ± 0.95 | 85.07 ± 0.70 | 0.844 |
| Meatiness (%) | 54.75 B ± 0.49 | 57.01 A ± 0.46 | 0.000 |
| Backfat thickness (mm) | 19.64 A ± 0.63 | 16.54 B ± 0.67 | 0.000 |
| Muscle thickness (mm) | 57.88 b ± 1.49 | 60.34 a ± 0.96 | 0.022 |
| Traits | Control | Bokashi | p-Value |
|---|---|---|---|
| pH35min | 6.63 ± 0.03 | 6.63 ± 0.03 | 0.883 |
| pH24 | 5.74 A ± 0.05 | 5.58 B ± 0.02 | 0.003 |
| Drip loss (%) EC24 (mS/cm) | 2.50 B ± 0.18 | 4.14 A ± 0.26 | 0.000 |
| 4.32 ± 0.17 | 3.95 ± 0.20 | 0.153 | |
| TY (%) | 94,67 A ± 1.10 | 87,52 B ± 0.60 | 0.000 |
| L* | 55.45 ± 0.57 | 56.04 ± 0.53 | 0.449 |
| a* | 5.48 ± 0.14 | 5.73 ± 0.14 | 0.209 |
| b* | 13.89 b ± 0.14 | 14.37 a ± 0.14 | 0.014 |
| C* | 14.94 b ± 0.16 | 15.49 a ± 0.14 | 0.011 |
| h0 | 68.53 ± 0.45 | 68.27 ± 0.52 | 0.705 |
| Shear force (kg) | 4.55 B ± 0.15 | 5.35 A ± 0.14 | 0.000 |
| Total protein (%) | 24.30 A ± 0.06 | 24.03 B ± 0.08 | 0.009 |
| Intramuscular fat (%) | 1.42 ± 0.09 | 1.46 ± 0.08 | 0.722 |
| Dry matter (%) | 26.08 ± 0.10 | 25.82 ± 0.10 | 0.069 |
| Traits | Control | Bokashi | p-Value |
|---|---|---|---|
| L* | 83.98 ± 0.44 | 83.23 ± 0.23 | 0.138 |
| a* | 2.76 ± 0.14 | 3.09 ± 0.15 | 0.098 |
| b* | 11.41 ± 0.22 | 11.00 ± 0.21 | 0.182 |
| C* | 11.76 ± 0.23 | 11.45 ± 0.21 | 0.334 |
| h0 | 76.43 a ± 0.61 | 73.98 b ± 0.75 | 0.014 |
| Traits (mg/kg) | Feed | Control | Bokashi | p-Value |
|---|---|---|---|---|
| K | 5216 | 2941.55 ± 16.17 | 2941.77 ± 18.97 | 0.993 |
| Na | 1414 | 346.30 b ± 4.18 | 361.81 a ± 4.73 | 0.017 |
| Mg | 1615 | 266.40 b ± 1.09 | 271.22 a ± 1.57 | 0.014 |
| P | 4909 | 2203.53 ± 10.76 | 2217.70 ± 11.27 | 0.367 |
| Cr | 0.12 | 0.04 ± 0.00 | 0.05 ± 0.00 | 0.087 |
| Mn | 81.7 | 0.09 ± 0.00 | 0.08 ± 0.00 | 0.538 |
| Fe | 203 | 5.21 ± 0.29 | 5.07 ± 0.11 | 0.662 |
| Cu | 20.1 | 0.40 ± 0.01 | 0.41 ± 0.01 | 0.456 |
| Zn | 106.2 | 13.74 ± 0.19 | 13.45 ± 0.20 | 0.318 |
| Ca | 6537 | 36.18 ± 0.46 | 37.44 ± 0.52 | 0.076 |
| Se | 0.18 | 0.08 B ± 0.00 | 0.09 A ± 0.00 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rybarczyk, A.; Bogusławska-Wąs, E.; Pilarczyk, B. Carcass and Pork Quality and Gut Environment of Pigs Fed a Diet Supplemented with the Bokashi Probiotic. Animals 2021, 11, 3590. https://doi.org/10.3390/ani11123590
Rybarczyk A, Bogusławska-Wąs E, Pilarczyk B. Carcass and Pork Quality and Gut Environment of Pigs Fed a Diet Supplemented with the Bokashi Probiotic. Animals. 2021; 11(12):3590. https://doi.org/10.3390/ani11123590
Chicago/Turabian StyleRybarczyk, Artur, Elżbieta Bogusławska-Wąs, and Bogumiła Pilarczyk. 2021. "Carcass and Pork Quality and Gut Environment of Pigs Fed a Diet Supplemented with the Bokashi Probiotic" Animals 11, no. 12: 3590. https://doi.org/10.3390/ani11123590