
Inflammatory Mediation of Heat Stress-Induced Growth Deficits in Livestock and Its Potential Role as a Target for Nutritional Interventions: A Review

Abstract
:Simple Summary
Abstract
1. Introduction
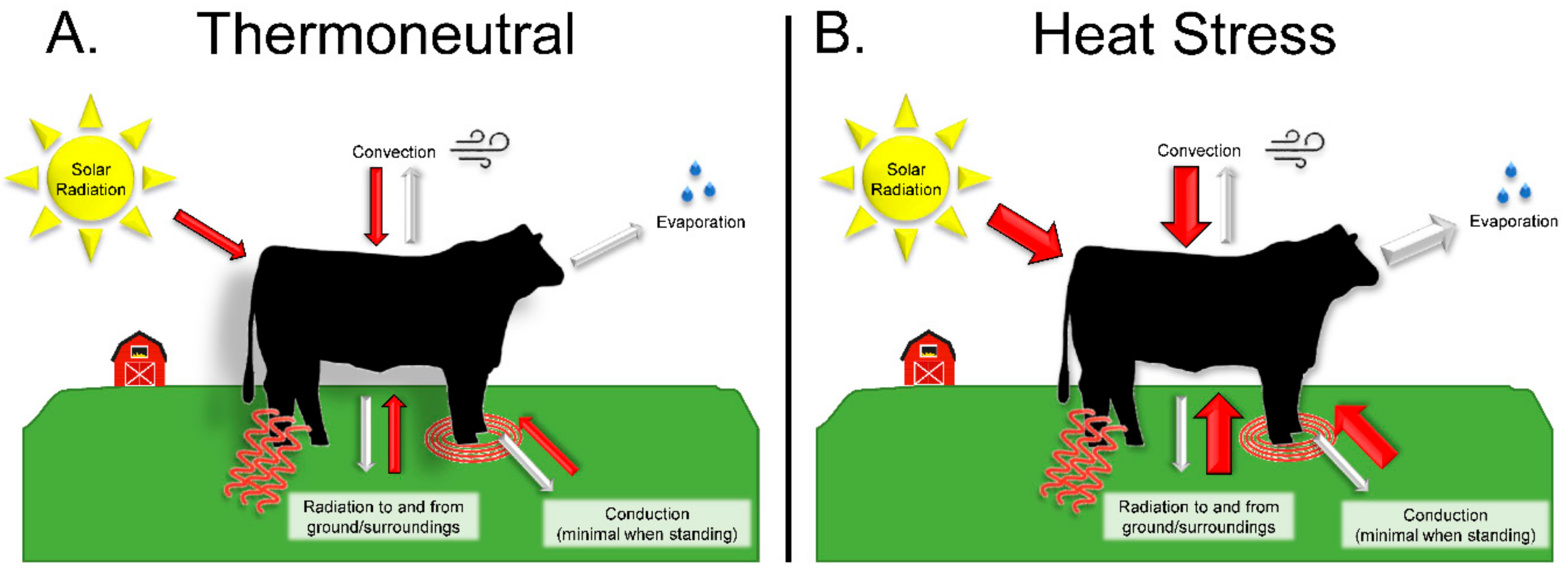
2. The Impact of Heat Stress in Livestock
2.1. Heat Stress Conditions
2.2. Consequences of Heat Stress
2.2.1. Hyperthermia
2.2.2. Hyperventilation
2.2.3. Endocrine Changes
2.2.4. Anorexia and Poor Growth Performance
2.3. Common Abatement Strategies for Heat Stress
2.3.1. Protection via Environmental Modification
2.3.2. Genetic Selection for Heat Tolerance
2.3.3. Nutritional Management of Heat-Stressed Livestock
3. The Role of Inflammation in Heat Stress
3.1. Inflammatory Regulation of Muscle Growth
3.2. Methods for Targeting Inflammation during Heat Stress
3.2.1. Brown Seaweed & Seaweed Extract
3.2.2. Resveratrol
3.2.3. Turmeric Curcumin
3.2.4. Vitamin E + Selenium
3.2.5. ω-3. Polyunsaturated Fatty Acids
3.2.6. Probiotics
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, M.; Dunshea, F.R.; Warner, R.D.; DiGiacomo, K.; Osei-Amponsah, R.; Chauhan, S.S. Impacts of heat stress on meat quality and strategies for amelioration: A review. Int. J. Biometeorol. 2020, 64, 1613–1628. [Google Scholar] [CrossRef]
- Carnovale, F.; Phillips, C.J.C. The Effects of Heat Stress on Sheep Welfare during Live Export Voyages from Australia to the Middle East. Animals 2020, 10, 694. [Google Scholar] [CrossRef] [PubMed]
- St-Pierre, N.R.; Cobanov, B.; Schnitkey, G. Economic Losses from Heat Stress by US Livestock Industries. J. Dairy Sci. 2003, 86, E52–E77. [Google Scholar] [CrossRef] [Green Version]
- Hahn, G.L. Dynamic responses of cattle to thermal heat loads. J. Anim. Sci. 1999, 77, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Gregory, N.G. How climatic changes could affect meat quality. Food Res. Int. 2010, 43, 1866–1873. [Google Scholar] [CrossRef]
- Meehl, G.A.; Tebaldi, C. More intense, more frequent, and longer lasting heat waves in the 21st century. Science 2004, 305, 994–997. [Google Scholar] [CrossRef] [Green Version]
- Hahn, G.; Mader, T. Heat waves in relation to thermoregulation, feeding behavior and mortality of feedlot cattle. In Proceedings of the Fifth International Livestock Environment Symposium, Bloomington, IN, USA, 1 May 1997. [Google Scholar]
- Brown-Brandl, T.M.; Eigenberg, R.A.; Hahn, G.L.; Nienaber, J.A.; Mader, T.L.; Spiers, D.E.; Parkhurst, A.M. Analyses of thermoregulatory responses of feeder cattle exposed to simulated heat waves. Int. J. Biometeorol. 2005, 49, 285–296. [Google Scholar] [CrossRef]
- Grandin, T. Evaluation of the welfare of cattle housed in outdoor feedlot pens. Vet. Anim. Sci. 2016, 1–2, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Mader, T.L. BILL E. KUNKLE INTERDISCIPLINARY BEEF SYMPOSIUM: Animal welfare concerns for cattle exposed to adverse environmental conditions1. J. Anim. Sci. 2014, 92, 5319–5324. [Google Scholar] [CrossRef] [Green Version]
- Lees, A.M.; Sejian, V.; Wallage, A.L.; Steel, C.C.; Mader, T.L.; Lees, J.C.; Gaughan, J.B. The Impact of Heat Load on Cattle. Animals 2019, 9, 322. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, K.F.; Mader, T.L. Managing Heat Stress Episodes in Confined Cattle. Vet. Clin. N. Am. Food Anim. Pract. 2018, 34, 325–339. [Google Scholar] [CrossRef] [PubMed]
- Kreikemeier, K.K.; Unruh, J.A.; Eck, T.P. Factors affecting the occurrence of dark-cutting beef and selected carcass traits in finished beef cattle. J. Anim. Sci. 1998, 76, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, T.L.; Cundiff, L.V.; Shackelford, S.D.; Koohmaraie, M. Characterization of biological types of cattle (Cycle V): Carcass traits and longissimus palatability. J. Anim. Sci. 2001, 79, 1209–1222. [Google Scholar] [CrossRef]
- Renaudeau, D.; Collin, A.; Yahav, S.; de Basilio, V.; Gourdine, J.L.; Collier, R.J. Adaptation to hot climate and strategies to alleviate heat stress in livestock production. Animal 2012, 6, 707–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pezzopane, J.R.M.; Nicodemo, M.L.F.; Bosi, C.; Garcia, A.R.; Lulu, J. Animal thermal comfort indexes in silvopastoral systems with different tree arrangements. J. Therm. Biol. 2019, 79, 103–111. [Google Scholar] [CrossRef]
- Sparke, E.; Young, B.; Gaughan, J.; Holt, M.; Goodwin, P. Heat Load in Feedlot Cattle; Meat and Livestock Australia Ltd.: Sydney, Australia, 2001. [Google Scholar]
- Kibler, H.H. Environmental Physiology and Shelter Engineering with Special Reference to Domestic Animals. In LXVII, Thermal Effects of Various Temperature-Humidity Combinations on Holstein Cattle as Measured by Eight Physiological Responses; Research bulletin, University of Missouri: Columbia, MO, USA, 1964. [Google Scholar]
- Thom, E.C. Cooling Degree-Days. Air Cond. Heating Ventil. 1958, 53, 65–72. [Google Scholar]
- Thom, E.C. The Discomfort Index. Weatherwise 1959, 12, 57–61. [Google Scholar] [CrossRef]
- Eigenberg, R.; Brown-Brandl, T.; Nienaber, J. Development of a livestock weather safety monitor for feedlot cattle. Appl. Eng. Agric. 2007, 23, 657–660. [Google Scholar] [CrossRef]
- Nienaber, J.A.; Hahn, G.L. Livestock production system management responses to thermal challenges. Int. J. Biometeorol. 2007, 52, 149–157. [Google Scholar] [CrossRef] [Green Version]
- Mader, T.L.; Gaughan, J.B.; Johnson, L.J.; Hahn, G.L. Tympanic temperature in confined beef cattle exposed to excessive heat load. Int. J. Biometeorol. 2010, 54, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Marai, I.F.M.; El-Darawany, A.A.; Fadiel, A.; Abdel-Hafez, M.A.M. Physiological traits as affected by heat stress in sheep—A review. Small Rumin. Res. 2007, 71, 1–12. [Google Scholar] [CrossRef]
- Archana, P.R.; Sejian, V.; Ruban, W.; Bagath, M.; Krishnan, G.; Aleena, J.; Manjunathareddy, G.B.; Beena, V.; Bhatta, R. Comparative assessment of heat stress induced changes in carcass traits, plasma leptin profile and skeletal muscle myostatin and HSP70 gene expression patterns between indigenous Osmanabadi and Salem Black goat breeds. Meat Sci. 2018, 141, 66–80. [Google Scholar] [CrossRef] [PubMed]
- Brown-Brandl, T.M.; Eigenberg, R.A.; Nienaber, J.A.; Hahn, G.L. Dynamic Response Indicators of Heat Stress in Shaded and Non-shaded Feedlot Cattle, Part 1: Analyses of Indicators. Biosyst. Eng. 2005, 90, 451–462. [Google Scholar] [CrossRef] [Green Version]
- Gaughan, J.B.; Mader, T.L.; Holt, S.M.; Lisle, A. A new heat load index for feedlot cattle. J. Anim. Sci. 2008, 86, 226–234. [Google Scholar] [CrossRef] [Green Version]
- Buffington, D.E.; Collazo-Arocho, A.; Canton, G.H.; Pitt, D.; Thatcher, W.W.; Collier, R.J. Black Globe-Humidity Index (BGHI) as Comfort Equation for Dairy Cows. Trans. ASAE 1981, 24, 0711–0714. [Google Scholar] [CrossRef]
- Barnes, T.L.; Cadaret, C.N.; Beede, K.A.; Schmidt, T.B.; Petersen, J.L.; Yates, D.T. Hypertrophic muscle growth and metabolic efficiency were impaired by chronic heat stress, improved by zilpaterol supplementation, and not affected by ractopamine supplementation in feedlot lambs1. J. Anim. Sci 2019, 97, 4101–4113. [Google Scholar] [CrossRef]
- Barnes, T.L.; Burrack, R.M.; Schmidt, T.B.; Petersen, J.L.; Yates, D.T. Sustained heat stress elevated corneal and body surface temperatures and altered circulating leukocytes and metabolic indicators in wether lambs supplemented with ractopamine or zilpaterol. J. Anim. Sci. 2021, 99, skab236. [Google Scholar] [CrossRef] [PubMed]
- Swanson, R.M.; Tait, R.G.; Galles, B.M.; Duffy, E.M.; Schmidt, T.B.; Petersen, J.L.; Yates, D.T. Heat stress-induced deficits in growth, metabolic efficiency, and cardiovascular function coincided with chronic systemic inflammation and hypercatecholaminemia in ractopamine-supplemented feedlot lambs. J. Anim. Sci. 2020, 98, skaa168. [Google Scholar] [CrossRef]
- Cadaret, C.N.; Abebe, M.D.; Barnes, T.L.; Posont, R.J.; Yates, D.T. Lipopolysaccharide endotoxin injections elevated salivary TNFalpha and corneal temperatures and induced dynamic changes in circulating leukocytes, inflammatory cytokines, and metabolic indicators in wether lambs. J. Anim. Sci. 2021, 99, skab120. [Google Scholar] [CrossRef]
- Cadaret, C.N.; Merrick, E.M.; Barnes, T.L.; Beede, K.A.; Posont, R.J.; Petersen, J.L.; Yates, D.T. Sustained maternal inflammation during the early third-trimester yields intrauterine growth restriction, impaired skeletal muscle glucose metabolism, and diminished beta-cell function in fetal sheep. J. Anim. Sci. 2019, 97, 4822–4833. [Google Scholar] [CrossRef]
- Sula, M.J.; Winslow, C.M.; Boileau, M.J.; Barker, L.D.; Panciera, R.J. Heat-related injury in lambs. J. Vet. Diagn. Investig. 2012, 24, 772–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busby, D.; Loy, J.D. Heat Stress in Feedlot Cattle: Producer Survey Results; Beef Research Report; Iowa State University: Aimes, IA, USA, 1997. [Google Scholar]
- Gaughan, J.B.; Mader, T.L.; Holt, S.M.; Josey, M.J.; Rowan, K.J. Heat tolerance of Boran and Tuli crossbred steers. J. Anim. Sci. 1999, 77, 2398–2405. [Google Scholar] [CrossRef] [PubMed]
- Hales, J.R.S.; Webster, M.E.D. Respiratory function during thermal tachypnoea in sheep. J. Physiol. 1967, 190, 241–260. [Google Scholar] [CrossRef] [Green Version]
- Sivakumar, A.V.N.; Singh, G.; Varshney, V.P. Antioxidants Supplementation on Acid Base Balance during Heat Stress in Goats. Asian-Australas J. Anim. Sci. 2010, 23, 1462–1468. [Google Scholar] [CrossRef]
- Johnson, H.D.; Vanjonack, W.J. Effects of environmental and other stressors on blood hormone patterns in lactating animals. J. Dairy Sci. 1976, 59, 1603–1617. [Google Scholar] [CrossRef]
- Brown-Brandl, T.M.; Chitko-McKown, C.G.; Eigenberg, R.A.; Mayer, J.J.; Welsh, T.H.; Davis, J.D.; Purswell, J.L. Physiological responses of feedlot heifers provided access to different levels of shade. Animal 2017, 11, 1344–1353. [Google Scholar] [CrossRef]
- Reith, R.R.; Sieck, R.L.; Grijalva, P.C.; Duffy, E.M.; Swanson, R.M.; Fuller, A.M.; Beede, K.A.; Beard, J.K.; Diaz, D.E.; Schmidt, T.B.; et al. Heat stress and β-adrenergic agonists alter the adipose transcriptome and fatty acid mobilization in ruminant livestock. Transl. Anim. Sci. 2020, 4, S141–S144. [Google Scholar] [CrossRef] [PubMed]
- Kubik, R.M.; Tietze, S.M.; Schmidt, T.B.; Yates, D.T.; Petersen, J.L. Investigation of the skeletal muscle transcriptome in lambs fed beta adrenergic agonists and subjected to heat stress for 21 d. Transl. Anim. Sci. 2018, 2, S53–S56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magdub, A.; Johnson, H.D.; Belyea, R.L. Effect of Environmental Heat and Dietary Fiber on Thyroid Physiology of Lactating Cows1. J. Dairy Sci. 1982, 65, 2323–2331. [Google Scholar] [CrossRef]
- Horowitz, M. Heat acclimation: Phenotypic plasticity and cues to the underlying molecular mechanisms. J. Therm. Biol. 2001, 26, 357–363. [Google Scholar] [CrossRef]
- Root, A.W.; Shulman, D.; Root, J.; Diamond, F. The interrelationships of thyroid and growth hormones: Effect of growth hormone releasing hormone in hypo- and hyperthyroid male rats. Acta Endocrinol. Suppl. 1986, 279, 367–375. [Google Scholar] [CrossRef]
- Capuco, A.V.; Kahl, S.; Jack, L.J.; Bishop, J.O.; Wallace, H. Prolactin and growth hormone stimulation of lactation in mice requires thyroid hormones. Proc. Soc. Exp. Biol. Med. 1999, 221, 345–351. [Google Scholar] [CrossRef] [PubMed]
- McGuire, M.A.; Beede, D.K.; Collier, R.J.; Buonomo, F.C.; DeLorenzo, M.A.; Wilcox, C.J.; Huntington, G.B.; Reynolds, C.K. Effects of acute thermal stress and amount of feed intake on concentrations of somatotropin, insulin-like growth factor (IGF)-I and IGF-II, and thyroid hormones in plasma of lactating Holstein cows2. J. Anim. Sci. 1991, 69, 2050–2056. [Google Scholar] [CrossRef]
- Leal-Cerro, A.; Soto, A.; Martínez, M.A.; Dieguez, C.; Casanueva, F.F. Influence of Cortisol Status on Leptin Secretion. Pituitary 2001, 4, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Mitlöhner, F.M.; Morrow, J.L.; Dailey, J.W.; Wilson, S.C.; Galyean, M.L.; Miller, M.F.; McGlone, J.J. Shade and water misting effects on behavior, physiology, performance, and carcass traits of heat-stressed feedlot cattle. J. Anim. Sci. 2001, 79, 2327–2335. [Google Scholar] [CrossRef]
- Gaughan, J.B.; Mader, T.L. Effects of sodium chloride and fat supplementation on finishing steers exposed to hot and cold conditions. J. Anim. Sci. 2009, 87, 612–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang-Fung-Martel, J.; Harrison, M.T.; Brown, J.N.; Rawnsley, R.; Smith, A.P.; Meinke, H. Negative relationship between dry matter intake and the temperature-humidity index with increasing heat stress in cattle: A global meta-analysis. Int. J. Biometeorol. 2021, 65, 2099–2109. [Google Scholar] [CrossRef]
- Nicolás-López, P.; Macías-Cruz, U.; Mellado, M.; Correa-Calderón, A.; Meza-Herrera, C.A.; Avendaño-Reyes, L. Growth performance and changes in physiological, metabolic and hematological parameters due to outdoor heat stress in hair breed male lambs finished in feedlot. Int. J. Biometeorol. 2021, 65, 1451–1459. [Google Scholar] [CrossRef] [PubMed]
- Russi, J.P.; DiLorenzo, N.; Relling, A.E. Effects of rumen-protected carbohydrate supplementation on performance and blood metabolites in feedlot finishing steers during heat stress. Transl. Anim. Sci. 2019, 3, 513–521. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, R.L.; Cadaret, C.N.; Swanson, R.M.; Beede, K.A.; Posont, R.J.; Schmidt, T.B.; Petersen, J.L.; Yates, D.T. Body composition estimated by bioelectrical impedance analyses is diminished by prenatal stress in neonatal lambs and by heat stress in feedlot wethers. Transl. Anim. Sci. 2019, 3, 1691–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mader, T.L.; Davis, M.S. Effect of management strategies on reducing heat stress of feedlot cattle: Feed and water intake1. J. Anim. Sci. 2004, 82, 3077–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernabucci, U.; Lacetera, N.; Baumgard, L.H.; Rhoads, R.P.; Ronchi, B.; Nardone, A. Metabolic and hormonal acclimation to heat stress in domesticated ruminants. Animal 2010, 4, 1167–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheelock, J.B.; Rhoads, R.P.; VanBaale, M.J.; Sanders, S.R.; Baumgard, L.H. Effects of heat stress on energetic metabolism in lactating Holstein cows1. J. Dairy Sci. 2010, 93, 644–655. [Google Scholar] [CrossRef]
- Rhoads, M.L.; Rhoads, R.P.; VanBaale, M.J.; Collier, R.J.; Sanders, S.R.; Weber, W.J.; Crooker, B.A.; Baumgard, L.H. Effects of heat stress and plane of nutrition on lactating Holstein cows: I. Production, metabolism, and aspects of circulating somatotropin. J. Dairy Sci. 2009, 92, 1986–1997. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Y.; Kronenfeld, J.M.; Renquist, B.J. Feed intake–dependent and –independent effects of heat stress on lactation and mammary gland development. J. Dairy Sci. 2020, 103, 12003–12014. [Google Scholar] [CrossRef] [PubMed]
- Belhadj Slimen, I.; Najar, T.; Ghram, A.; Abdrrabba, M. Heat stress effects on livestock: Molecular, cellular and metabolic aspects, a review. J. Anim. Physiol. Anim. Nutr. 2016, 100, 401–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, S.; Summers, C.M.; Pearce, S.C.; Gabler, N.K.; Valentine, R.J.; Baumgard, L.H.; Rhoads, R.P.; Selsby, J.T. Short-term heat stress causes altered intracellular signaling in oxidative skeletal muscle. J. Anim. Sci. 2017, 95, 2438–2451. [Google Scholar] [CrossRef] [PubMed]
- Rhoads, R.P.; Obrien, M.D.; Greer, K.; Cole, L.; Sanders, S.; Wheelock, J.B.; Baumgard, L.H. Consequences of heat stress on the profile of skeletal muscle gene expression in beef cattle. FASEB J. 2008, 22, 1165.1161. [Google Scholar] [CrossRef]
- Fernández-Fernández, M.R.; Valpuesta, J.M. Hsp70 chaperone: A master player in protein homeostasis. F1000 Res. 2018, 7, F1000. [Google Scholar] [CrossRef] [Green Version]
- Mkize, L.S.; Zishiri, O.T. Novel single nucleotide polymorphisms in the heat shock protein 70.1 gene in South African Nguni crossbred cattle. Trop. Anim. Health Prod. 2020, 52, 893–901. [Google Scholar] [CrossRef]
- Blackshaw, J.; Blackshaw, A. Heat stress in cattle and the effect of shade on production and behaviour: A review. Aust. J. Exp. Agric. 1994, 34, 285–295. [Google Scholar] [CrossRef]
- Beede, D.K.; Collier, R.J. Potential Nutritional Strategies for Intensively Managed Cattle during Thermal Stress. J. Anim. Sci. 1986, 62, 543–554. [Google Scholar] [CrossRef]
- Mitlohner, F.M.; Galyean, M.L.; McGlone, J.J. Shade effects on performance, carcass traits, physiology, and behavior of heat-stressed feedlot heifers. J. Anim. Sci. 2002, 80, 2043–2050. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, D.V. Heat Stress Interaction with Shade and Cooling. J. Dairy Sci. 1994, 77, 2044–2050. [Google Scholar] [CrossRef]
- Mader, T.L.; Griffin, D. Management of Cattle Exposed to Adverse Environmental Conditions. Vet. Clin. N. Am. Food Anim. Pract. 2015, 31, 247–258. [Google Scholar] [CrossRef]
- Hahn, G.L. Management and Housing of Farm Animals in Hot Environments; CRC Press, Inc.: Boca Raton, FL, USA, 1985; pp. 151–174. [Google Scholar]
- Mader, T.L.; Davis, M.S.; Gaughan, J.B. Effect of sprinkling on feedlot microclimate and cattle behavior. Int. J. Biometeorol. 2007, 51, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Dahl, G.E.; Tao, S.; Laporta, J. Heat Stress Impacts Immune Status in Cows Across the Life Cycle. Front. Vet. Sci. 2020, 7, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leighton, R.E.; Rupel, I.W.; Williams, J.S.; Shrode, R.R. A study of the influence of solar radiation on physiological responses of dairy cattle. J. Dairy Sci. 1960, 43, 1245–1254. [Google Scholar]
- Mader, T.L.; Holt, S.M.; Hahn, G.L.; Davis, M.S.; Spiers, D.E. Feeding strategies for managing heat load in feedlot cattle. J. Anim. Sci. 2002, 80, 2373–2382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McManus, C.; Louvandini, H.; Gugel, R.; Sasaki, L.C.B.; Bianchini, E.; Bernal, F.E.M.; Paiva, S.R.; Paim, T.P. Skin and coat traits in sheep in Brazil and their relation with heat tolerance. Trop. Anim. Health Prod. 2011, 43, 121–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, P.J. Physiological and cellular adaptations of zebu cattle to thermal stress. Anim. Reprod. Sci. 2004, 82–83, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Frisch, J.E.; Vercoe, J.E. Food intake, eating rate, weight gains, metabolic rate and efficiency of feed utilization in Bos taurus and Bos indicus crossbred cattle. Anim. Sci. 1977, 25, 343–358. [Google Scholar] [CrossRef]
- Taye, M.; Lee, W.; Caetano-Anolles, K.; Dessie, T.; Hanotte, O.; Mwai, O.A.; Kemp, S.; Cho, S.; Oh, S.J.; Lee, H.-K.; et al. Whole genome detection of signature of positive selection in African cattle reveals selection for thermotolerance. Anim. Sci. J. 2017, 88, 1889–1901. [Google Scholar] [CrossRef] [PubMed]
- Finch, V. Comparison of non-evaporative heat transfer in different cattle breeds. Aust. J. Agric. Res. 1985, 36, 497–508. [Google Scholar] [CrossRef]
- Mateescu, R.G.; Sarlo-Davila, K.M.; Dikmen, S.; Rodriguez, E.; Oltenacu, P.A. The effect of Brahman genes on body temperature plasticity of heifers on pasture under heat stress. J. Anim. Sci. 2020, 98, skaa126. [Google Scholar] [CrossRef] [PubMed]
- Hammond, A.C.; Olson, T.A.; Chase, C.C., Jr.; Bowers, E.J.; Randel, R.D.; Murphy, C.N.; Vogt, D.W.; Tewolde, A. Heat tolerance in two tropically adapted Bos taurus breeds, Senepol and Romosinuano, compared with Brahman, Angus, and Hereford cattle in Florida. J. Anim. Sci. 1996, 74, 295–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, A.M.F.; Titto, E.L.; Infante, P.; Titto, C.G.; Geraldo, A.M.; Alves, A.; Leme, T.M.; Baccari, F.; Almeida, J.A. Evaporative heat loss in Bos taurus: Do different cattle breeds cope with heat stress in the same way? J. Therm. Biol. 2014, 45, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Elzo, M.A.; Johnson, D.D.; Wasdin, J.G.; Driver, J.D. Carcass and meat palatability breed differences and heterosis effects in an Angus–Brahman multibreed population. Meat Sci. 2012, 90, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Mader, T.L. Environmental stress in confined beef cattle1. J. Anim. Sci. 2003, 81, E110–E119. [Google Scholar] [CrossRef]
- Broadway, P.R.; Carroll, J.A.; Burdick Sanchez, N.C.; Cravey, M.D.; Corley, J.R. Some Negative Effects of Heat Stress in Feedlot Heifers May Be Mitigated via Yeast Probiotic Supplementation. Front. Vet. Sci. 2019, 6, 515. [Google Scholar] [CrossRef]
- Valadez-García, K.M.; Avendaño-Reyes, L.; Díaz-Molina, R.; Mellado, M.; Meza-Herrera, C.A.; Correa-Calderón, A.; Macías-Cruz, U. Free ferulic acid supplementation of heat-stressed hair ewe lambs: Oxidative status, feedlot performance, carcass traits and meat quality. Meat Sci. 2021, 173, 108395. [Google Scholar] [CrossRef]
- Boyd, B.M.; Shackelford, S.D.; Hales, K.E.; Brown-Brandl, T.M.; Bremer, M.L.; Spangler, M.L.; Wheeler, T.L.; King, D.A.; Erickson, G.E. Effects of shade and feeding zilpaterol hydrochloride to finishing steers on performance, carcass quality, heat stress, mobility, and body temperature. J. Anim. Sci. 2015, 93, 5801–5811. [Google Scholar] [CrossRef] [PubMed]
- Bagath, M.; Krishnan, G.; Devaraj, C.; Rashamol, V.P.; Pragna, P.; Lees, A.M.; Sejian, V. The impact of heat stress on the immune system in dairy cattle: A review. Res. Vet. Sci. 2019, 126, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Hall, L.W.; Villar, F.; Chapman, J.D.; McLean, D.J.; Long, N.M.; Xiao, Y.; Collier, J.L.; Collier, R.J. An evaluation of an immunomodulatory feed ingredient in heat-stressed lactating Holstein cows: Effects on hormonal, physiological, and production responses. J. Dairy Sci. 2018, 101, 7095–7105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelnour, S.A.; Abd El-Hack, M.E.; Khafaga, A.F.; Arif, M.; Taha, A.E.; Noreldin, A.E. Stress biomarkers and proteomics alteration to thermal stress in ruminants: A review. J. Therm. Biol. 2019, 79, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Carswell, E.A.; Old, L.J.; Kassel, R.L.; Green, S.; Fiore, N.; Williamson, B. An endotoxin-induced serum factor that causes necrosis of tumors. Proc. Natl. Acad. Sci. USA 1975, 72, 3666–3670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borish, L.C.; Steinke, J.W. 2. Cytokines and chemokines. J. Allergy Clin. Immunol. 2003, 111, S460–S475. [Google Scholar] [CrossRef] [PubMed]
- Zelová, H.; Hošek, J. TNF-α signalling and inflammation: Interactions between old acquaintances. Inflamm. Res. 2013, 62, 641–651. [Google Scholar] [CrossRef]
- Chen, S.; Wang, J.; Peng, D.; Li, G.; Chen, J.; Gu, X. Exposure to heat-stress environment affects the physiology, circulation levels of cytokines, and microbiome in dairy cows. Sci. Rep. 2018, 8, 14606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, L.; Zheng, N.; Zhao, S.; Cheng, J.; Yang, Y.; Zhang, Y.; Yang, H.; Wang, J. Long-term heat stress induces the inflammatory response in dairy cows revealed by plasma proteome analysis. Biochem. Biophys. Res. Commun. 2016, 471, 296–302. [Google Scholar] [CrossRef]
- Park, D.S.; Gu, B.H.; Park, Y.J.; Joo, S.S.; Lee, S.S.; Kim, S.H.; Kim, E.T.; Kim, D.H.; Lee, S.S.; Lee, S.J.; et al. Dynamic changes in blood immune cell composition and function in Holstein and Jersey steers in response to heat stress. Cell Stress Chaperones. 2021, 26, 705–720. [Google Scholar] [CrossRef]
- Dodd, S.L.; Gagnon, B.J.; Senf, S.M.; Hain, B.A.; Judge, A.R. Ros-mediated activation of NF-kappaB and Foxo during muscle disuse. Muscle Nerve 2010, 41, 110–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauhan, S.S.; Rashamol, V.P.; Bagath, M.; Sejian, V.; Dunshea, F.R. Impacts of heat stress on immune responses and oxidative stress in farm animals and nutritional strategies for amelioration. Int. J. Biometeorol. 2021, 65, 1231–1244. [Google Scholar] [CrossRef]
- Song, J.H.; Kim, K.J.; Choi, S.Y.; Koh, E.J.; Park, J.; Lee, B.Y. Korean ginseng extract ameliorates abnormal immune response through the regulation of inflammatory constituents in Sprague Dawley rat subjected to environmental heat stress. J. Ginseng. Res. 2019, 43, 252–260. [Google Scholar] [CrossRef]
- Mohyuddin, S.G.; Qamar, A.; Hu, C.-y.; Li, Y.; Chen, S.-W.; Wen, J.-y.; Bao, M.-L.; Ju, X.H. Terpinen4-ol inhibits heat stress induced inflammation in colonic tissue by Activating Occludin, Claudin-2 and TLR4/NF-κB signaling pathway. Int. Immunopharmacol. 2021, 99, 107727. [Google Scholar] [CrossRef] [PubMed]
- Mohyuddin, S.G.; Qamar, A.; Hu, C.Y.; Chen, S.W.; Wen, J.Y.; Liu, X.X.; Ma, X.B.; Yu, Z.C.; Yong, Y.H.; Wu, L.Y.; et al. Effect of chitosan on blood profile, inflammatory cytokines by activating TLR4/NF-κB signaling pathway in intestine of heat stressed mice. Sci. Rep. 2021, 11, 20608. [Google Scholar] [CrossRef]
- Ganesan, S.; Reynolds, C.; Hollinger, K.; Pearce, S.C.; Gabler, N.K.; Baumgard, L.H.; Rhoads, R.P.; Selsby, J.T. Twelve hours of heat stress induces inflammatory signaling in porcine skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R1288–R1296. [Google Scholar] [CrossRef] [Green Version]
- Chei, S.; Song, J.H.; Oh, H.J.; Lee, K.; Jin, H.; Choi, S.H.; Nah, S.Y.; Lee, B.Y. Gintonin-Enriched Fraction Suppresses Heat Stress-Induced Inflammation Through LPA Receptor. Molecules 2020, 25, 1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauhan, S.S.; Celi, P.; Fahri, F.T.; Leury, B.J.; Dunshea, F.R. Dietary antioxidants at supranutritional doses modulate skeletal muscle heat shock protein and inflammatory gene expression in sheep exposed to heat stress. J. Anim. Sci. 2014, 92, 4897–4908. [Google Scholar] [CrossRef] [PubMed]
- Maltzahn, J.; Bentzinger, C.; Rudnicki, M. Characteristics of Satellite Cells and Multipotent Adult Stem Cells in the Skeletal Muscle. In Stem Cells and Cancer Stem Cells; Springer Nature: London, UK, 2014; pp. 63–73. [Google Scholar]
- Chal, J.; Pourquié, O. Making muscle: Skeletal myogenesis in vivo and in vitro. Development 2017, 144, 2104–2122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bi, P.; Kuang, S. Meat Science and Muscle Biology Symposium: Stem cell niche and postnatal muscle growth. J. Anim. Sci. 2012, 90, 924–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ten Broek, R.W.; Grefte, S.; Von den Hoff, J.W. Regulatory factors and cell populations involved in skeletal muscle regeneration. J. Cell Physiol. 2010, 224, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.A.; Fiorotto, M.L. Regulation of muscle growth in neonates. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 78–85. [Google Scholar] [CrossRef] [Green Version]
- Dhawan, J.; Rando, T.A. Stem cells in postnatal myogenesis: Molecular mechanisms of satellite cell quiescence, activation and replenishment. Trends Cell Biol. 2005, 15, 666–673. [Google Scholar] [CrossRef]
- Posont, R.J.; Beede, K.A.; Limesand, S.W.; Yates, D.T. Changes in myoblast responsiveness to TNFalpha and IL-6 contribute to decreased skeletal muscle mass in intrauterine growth restricted fetal sheep. Transl. Anim. Sci. 2018, 2, S44–S47. [Google Scholar] [CrossRef] [PubMed]
- Al-Shanti, N.; Saini, A.; Faulkner, S.H.; Stewart, C.E. Beneficial synergistic interactions of TNF-alpha and IL-6 in C2 skeletal myoblasts--potential cross-talk with IGF system. Growth Factors 2008, 26, 61–73. [Google Scholar] [CrossRef]
- Shirakawa, T.; Rojasawasthien, T.; Inoue, A.; Matsubara, T.; Kawamoto, T.; Kokabu, S. Tumor necrosis factor alpha regulates myogenesis to inhibit differentiation and promote proliferation in satellite cells. Biochem. Biophys. Res. Commun. 2021, 580, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, A.M.; DeOcesano-Pereira, C.; Teixeira, C.; Moreira, V. IL-1β and TNF-α Modulation of Proliferated and Committed Myoblasts: IL-6 and COX-2-Derived Prostaglandins as Key Actors in the Mechanisms Involved. Cells 2020, 9, 2005. [Google Scholar] [CrossRef]
- Kumar, A.; Bhatnagar, S.; Paul, P.K. TWEAK and TRAF6 regulate skeletal muscle atrophy. Curr. Opin. Clin. Nutr. Metab. Care 2012, 15, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Mueck, T.; Berger, F.; Buechsler, I.; Valchanova, R.S.; Landuzzi, L.; Lollini, P.L.; Klingel, K.; Munz, B. TRAF6 regulates proliferation and differentiation of skeletal myoblasts. Differentiation 2011, 81, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, D.; Cui, Y.; Lu, S.; Gao, D.; Liu, J. Proinflammatory macrophages impair skeletal muscle differentiation in obesity through secretion of tumor necrosis factor-α via sustained activation of p38 mitogen-activated protein kinase. J. Cell Physiol. 2019, 234, 2566–2580. [Google Scholar] [CrossRef]
- O’Brien, M.E.; Londino, J.; McGinnis, M.; Weathington, N.; Adair, J.; Suber, T.; Kagan, V.; Chen, K.; Zou, C.; Chen, B.; et al. Tumor Necrosis Factor Alpha Regulates Skeletal Myogenesis by Inhibiting SP1 Interaction with cis-Acting Regulatory Elements within the Fbxl2 Gene Promoter. Mol. Cell Biol. 2020, 40, e00040-20. [Google Scholar] [CrossRef] [PubMed]
- Frost, R.A.; Lang, C.H.; Gelato, M.C. Transient Exposure of Human Myoblasts to Tumor Necrosis Factor-α Inhibits Serum and Insulin-Like Growth Factor-I Stimulated Protein Synthesis. Endocrinology 1997, 138, 4153–4159. [Google Scholar] [CrossRef] [PubMed]
- Tracey, K.J.; Wei, H.; Manogue, K.R.; Fong, Y.; Hesse, D.G.; Nguyen, H.T.; Kuo, G.C.; Beutler, B.; Cotran, R.S.; Cerami, A.; et al. Cachectin/tumor necrosis factor induces cachexia, anemia, and inflammation. J. Exp. Med. 1988, 167, 1211–1227. [Google Scholar] [CrossRef]
- Fong, Y.; Moldawer, L.L.; Marano, M.; Wei, H.; Barber, A.; Manogue, K.; Tracey, K.J.; Kuo, G.; Fischman, D.A.; Cerami, A.; et al. Cachectin/TNF or IL-1 alpha induces cachexia with redistribution of body proteins. Am. J. Physiol. 1989, 256, R659–R665. [Google Scholar] [CrossRef] [PubMed]
- Colditz, I.G. Effects of the immune system on metabolism: Implications for production and disease resistance in livestock. Livest. Prod. Sci. 2002, 75, 257–268. [Google Scholar] [CrossRef]
- Gifford, C.A.; Holland, B.P.; Mills, R.L.; Maxwell, C.L.; Farney, J.K.; Terrill, S.J.; Step, D.L.; Richards, C.J.; Burciaga Robles, L.O.; Krehbiel, C.R. Growth and Development Symposium: Impacts of inflammation on cattle growth and carcass merit. J. Anim. Sci. 2012, 90, 1438–1451. [Google Scholar] [CrossRef]
- Schneider, M.J.; Tait, R.G., Jr.; Busby, W.D.; Reecy, J.M. An evaluation of bovine respiratory disease complex in feedlot cattle: Impact on performance and carcass traits using treatment records and lung lesion scores. J. Anim. Sci. 2009, 87, 1821–1827. [Google Scholar] [CrossRef]
- Cadaret, C.N.; Posont, R.J.; Beede, K.A.; Riley, H.E.; Loy, J.D.; Yates, D.T. Maternal inflammation at midgestation impairs subsequent fetal myoblast function and skeletal muscle growth in rats, resulting in intrauterine growth restriction at term. Transl. Anim. Sci. 2019, 3, txz037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadaret, C.N.; Beede, K.A.; Riley, H.E.; Yates, D.T. Acute exposure of primary rat soleus muscle to zilpaterol HCl (beta2 adrenergic agonist), TNFalpha, or IL-6 in culture increases glucose oxidation rates independent of the impact on insulin signaling or glucose uptake. Cytokine 2017, 96, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Yende, S.R.; Harle, U.N.; Chaugule, B.B. Therapeutic potential and health benefits of Sargassum species. Pharmacogn. Rev. 2014, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Braden, K.W.; Blanton, J.R., Jr.; Montgomery, J.L.; van Santen, E.; Allen, V.G.; Miller, M.F. Tasco supplementation: Effects on carcass characteristics, sensory attributes, and retail display shelf-life. J. Anim. Sci. 2007, 85, 754–768. [Google Scholar] [CrossRef] [PubMed]
- Allen, V.G.; Pond, K.R.; Saker, K.E.; Fontenot, J.P.; Bagley, C.P.; Ivy, R.L.; Evans, R.R.; Brown, C.P.; Miller, M.F.; Montgomery, J.L.; et al. Tasco-Forage: III. Influence of a seaweed extract on performance, monocyte immune cell response, and carcass characteristics in feedlot-finished steers. J. Anim. Sci. 2001, 79, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Saker, K.E.; Allen, V.G.; Fontenot, J.P.; Bagley, C.P.; Ivy, R.L.; Evans, R.R.; Wester, D.B. Tasco-Forage: II. Monocyte immune cell response and performance of beef steers grazing tall fescue treated with a seaweed extract. J. Anim. Sci. 2001, 79, 1022–1031. [Google Scholar] [CrossRef] [Green Version]
- Ellamie, A.M.; Fouda, W.A.; Ibrahim, W.M.; Ramadan, G. Dietary supplementation of brown seaweed (Sargassum latifolium) alleviates the environmental heat stress-induced toxicity in male Barki sheep (Ovis aries). J. Therm. Biol. 2020, 89, 102561. [Google Scholar] [CrossRef]
- Yates, D.; Salisbury, M.; Ross, T.; Anderson, H. Effects of Tasco-14 supplementation on growth and fertility traits in young male Boer goats experiencing heat stress. Tex. J. Agric. Nat. Resour. 2010, 23, 12–18. [Google Scholar]
- Malaguarnera, L. Influence of Resveratrol on the Immune Response. Nutrients 2019, 11, 946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, K.; Yan, E.; Song, Z.; Li, S.; Zhang, H.; Zhang, L.; Wang, C.; Wang, T. Protective effect of resveratrol against hepatic damage induced by heat stress in a rat model is associated with the regulation of oxidative stress and inflammation. J. Therm. Biol. 2019, 82, 70–75. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Yu, Q.; He, Y.; Hu, R.; Xia, S.; He, J. Dietary resveratrol supplementation inhibits heat stress-induced high-activated innate immunity and inflammatory response in spleen of yellow-feather broilers. Poult. Sci. 2019, 98, 6378–6387. [Google Scholar] [CrossRef]
- Menon, V.P.; Sudheer, A.R. Antioxidant and anti-inflammatory properties of curcumin. Adv. Exp. Med. Biol. 2007, 595, 105–125. [Google Scholar] [CrossRef]
- Jiang, Z.; Wan, Y.; Li, P.; Xue, Y.; Cui, W.; Chen, Q.; Chen, J.; Wang, F.; Mao, D. Effect of Curcumin Supplement in Summer Diet on Blood Metabolites, Antioxidant Status, Immune Response, and Testicular Gene Expression in Hu Sheep. Animals 2019, 9, 720. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.H.; Shen, C.F.; Wang, G.J.; Kang, Y.; Song, Y.H.; Liu, J.W. Curcumin alleviates acute kidney injury in a dry-heat environment by reducing oxidative stress and inflammation in a rat model. J. Biochem. Mol. Toxicol. 2021, 35, e22630. [Google Scholar] [CrossRef] [PubMed]
- Hoehle, S.I.; Pfeiffer, E.; Sólyom, A.M.; Metzler, M. Metabolism of Curcuminoids in Tissue Slices and Subcellular Fractions from Rat Liver. J. Agric. Food Chem. 2006, 54, 756–764. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhai, Y.; Heng, X.; Che, F.Y.; Chen, W.; Sun, D.; Zhai, G. Oral bioavailability of curcumin: Problems and advancements. J. Drug Target. 2016, 24, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Diplock, A.T.; Lucy, J.A. The biochemical modes of action of vitamin e and selenium: A hypothesis. FEBS Lett. 1973, 29, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Traber, M.G.; Atkinson, J. Vitamin E, antioxidant and nothing more. Free. Radic. Biol. Med. 2007, 43, 4–15. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, S.S.; Celi, P.; Leury, B.J.; Clarke, I.J.; Dunshea, F.R. Dietary antioxidants at supranutritional doses improve oxidative status and reduce the negative effects of heat stress in sheep. J. Anim. Sci. 2014, 92, 3364–3374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellés, M.; del Mar Campo, M.; Roncalés, P.; Beltrán, J.A. Supranutritional doses of vitamin E to improve lamb meat quality. Meat Sci. 2019, 149, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Inoue, T.; Tanaka, M.; Masuda, S.; Ohue-Kitano, R.; Yamakage, H.; Muranaka, K.; Wada, H.; Kusakabe, T.; Shimatsu, A.; Hasegawa, K.; et al. Omega-3 polyunsaturated fatty acids suppress the inflammatory responses of lipopolysaccharide-stimulated mouse microglia by activating SIRT1 pathways. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 552–560. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Liu, W.; Shi, M.; Yang, Z.; Zhang, X.; Gong, P. Docosahexaenoic acid attenuates LPS-stimulated inflammatory response by regulating the PPARgamma/NF-kappaB pathways in primary bovine mammary epithelial cells. Res. Vet. Sci. 2017, 112, 7–12. [Google Scholar] [CrossRef]
- Velten, M.; Britt, R.D., Jr.; Heyob, K.M.; Tipple, T.E.; Rogers, L.K. Maternal dietary docosahexaenoic acid supplementation attenuates fetal growth restriction and enhances pulmonary function in a newborn mouse model of perinatal inflammation. J. Nutr. 2014, 144, 258–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caughey, G.E.; Mantzioris, E.; Gibson, R.A.; Cleland, L.G.; James, M.J. The effect on human tumor necrosis factor alpha and interleukin 1 beta production of diets enriched in n-3 fatty acids from vegetable oil or fish oil. Am. J. Clin. Nutr. 1996, 63, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wu, S.; Chen, C.; Xie, B.; Fang, Z.; Hu, W.; Chen, J.; Fu, H.; He, H. Omega-3 polyunsaturated fatty acid supplementation attenuates microglial-induced inflammation by inhibiting the HMGB1/TLR4/NF-κB pathway following experimental traumatic brain injury. J. Neuroinflamm. 2017, 14, 143. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Wu, Y.; Zhao, H.; Sun, H.; Gao, S. Maresin 1 mitigates renal ischemia/reperfusion injury in mice via inhibition of the TLR4/MAPK/NF-kappaB pathways and activation of the Nrf2 pathway. Drug Des. Devel Ther. 2019, 13, 739–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, H.; Wang, H.; Wang, S.; Tu, Z.; Zhang, L.; Wang, X.; Hou, Y.; Wang, C.; Chen, J.; Liu, Y. Flaxseed Oil Attenuates Intestinal Damage and Inflammation by Regulating Necroptosis and TLR4/NOD Signaling Pathways Following Lipopolysaccharide Challenge in a Piglet Model. Mol. Nutr. Food. Res. 2018, 62, e1700814. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, F.; Odle, J.; Lin, X.; Zhu, H.; Shi, H.; Hou, Y.; Yin, J. Fish oil increases muscle protein mass and modulates Akt/FOXO, TLR4, and NOD signaling in weanling piglets after lipopolysaccharide challenge. J. Nutr. 2013, 143, 1331–1339. [Google Scholar] [CrossRef] [Green Version]
- Soto, G.; Rodriguez, M.J.; Fuentealba, R.; Treuer, A.V.; Castillo, I.; Gonzalez, D.R.; Zuniga-Hernandez, J. Maresin 1, a Proresolving Lipid Mediator, Ameliorates Liver Ischemia-Reperfusion Injury and Stimulates Hepatocyte Proliferation in Sprague-Dawley Rats. Int. J. Mol. Sci. 2020, 21, 540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Z.; Lamont, G.J.; Lamont, R.J.; Uriarte, S.M.; Wang, H.; Scott, D.A. Resolvin D1, resolvin D2 and maresin 1 activate the GSK3β anti-inflammatory axis in TLR4-engaged human monocytes. Innate Immun. 2016, 22, 186–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roque-Jiménez, J.A.; Rosa-Velázquez, M.; Pinos-Rodríguez, J.M.; Vicente-Martínez, J.G.; Mendoza-Cervantes, G.; Flores-Primo, A.; Lee-Rangel, H.A.; Relling, A.E. Role of Long Chain Fatty Acids in Developmental Programming in Ruminants. Animals 2021, 11, 762. [Google Scholar] [CrossRef]
- Nickles, K.R.; Hamer, L.; Coleman, D.N.; Relling, A.E. Supplementation with eicosapentaenoic and docosahexaenoic acids in late gestation in ewes changes adipose tissue gene expression in the ewe and growth and plasma concentration of ghrelin in the offspring1. J. Anim. Sci. 2019, 97, 2631–2643. [Google Scholar] [CrossRef]
- Carranza Martin, A.C.; Coleman, D.N.; Garcia, L.G.; Furnus, C.C.; Relling, A.E. Prepartum fatty acid supplementation in sheep. III. Effect of eicosapentaenoic acid and docosahexaenoic acid during finishing on performance, hypothalamus gene expression, and muscle fatty acids composition in lambs. J. Anim. Sci. 2018, 96, 5300–5310. [Google Scholar] [CrossRef]
- Mohtashami, B.; Khalilvandi-Behroozyar, H.; Pirmohammadi, R.; Dehghan-Banadaky, M.; Kazemi-Bonchenari, M.; Dirandeh, E.; Ghaffari, M.H. The effect of supplemental bioactive fatty acids on growth performance and immune function of milk-fed Holstein dairy calves during heat stress. Br. J. Nutr. 2021, 126, 1–14. [Google Scholar] [CrossRef]
- Most, M.S.; Grijalva, P.C.; Beer, H.N.; Gibbs, R.L.; Hicks, Z.H.; Lacey, T.A.; Schmidt, T.B.; Petersen, J.L.; Yates, D.T. Dexamethasone and fish oil improve average daily gain but not muscle mass or protein content in feedlot wethers after chronic heat stress. Transl. Anim. Sci. 2021, 5, S46–S50. [Google Scholar] [CrossRef]
- Grijalva, P.C.; Most, M.S.; Gibbs, R.L.; Hicks, Z.H.; Lacey, T.A.; Beer, H.N.; Schmidt, T.B.; Petersen, J.L.; Yates, D.T. Fish oil and dexamethasone administration partially mitigate heat stress-induced changes in circulating leukocytes and metabolic indicators in feedlot wethers. Transl. Anim. Sci. 2021, 5, S30–S33. [Google Scholar] [CrossRef]
- Wistuba, T.J.; Kegley, E.B.; Apple, J.K. Influence of fish oil in finishing diets on growth performance, carcass characteristics, and sensory evaluation of cattle. J. Anim. Sci. 2006, 84, 902–909. [Google Scholar] [CrossRef]
- Malyar, R.M.; Naseri, E.; Li, H.; Ali, I.; Farid, R.A.; Liu, D.; Maroof, K.; Nasim, M.; Banuree, S.A.H.; Huang, K.; et al. Hepatoprotective Effects of Selenium-Enriched Probiotics Supplementation on Heat-Stressed Wistar Rat Through Anti-Inflammatory and Antioxidant Effects. Biol. Trace Elem. Res. 2021, 199, 3445–3456. [Google Scholar] [CrossRef] [PubMed]
- Mayorga, E.J.; Kvidera, S.K.; Horst, E.A.; Al-Qaisi, M.; McCarthy, C.S.; Abeyta, M.A.; Lei, S.; Elsasser, T.H.; Kahl, S.; Kiros, T.G.; et al. Effects of dietary live yeast supplementation on growth performance and biomarkers of metabolism and inflammation in heat-stressed and nutrient-restricted pigs. Transl. Anim. Sci. 2021, 5, txab072. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| THI Category | THI Ranges by Species | Respiration | Body Temperature Divergence (°C) | Behavioral Responses | Stress Level | |
|---|---|---|---|---|---|---|
| Cattle 1 | Sheep 2 & Goats 3 | |||||
| Normal | <74.0 | <82.0 | Normal | 0.00 | Normal | None |
| Alert | 74.0 to 79.0 | 82.0 to 84.0 | Steady but increased breathing | +0.11 | Increased standing | Mild |
| Danger | 79.1 to 84.0 | 84.1 to 86.0 | Rapid, shallow breaths | +0.28 | Bunching, ↓ feed intake | Moderate |
| Emergency | >84.0 | >86.0 | Deep, abdominal breaths with tongue extended | +0.56 | Lingering at water source | Severe |
| Supplement | Source | Species Tested | References |
|---|---|---|---|
| Brown seaweed/extract | Sargassum & Ascophyllum spp. | Cattle, sheep, goats | [127,128,129,130,131,132] |
| Resveratrol | Vitis spp. | Rats, chickens | [133,134,135] |
| Turmeric curcumin | Curcuma longa | Sheep, rats | [136,137,138,139,140] |
| Vitamin E + Selenium | Commercial supplements | Sheep, goats, | [104,141,142,143,144] |
| ω-3 PUFA 1 | EPA 2, DHA 3, fish oil | Cattle, sheep, mice, pigs, rats | [145,146,147,148,149,150,151,152,153,154,155,156,157,158,159,160,161] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Most, M.S.; Yates, D.T. Inflammatory Mediation of Heat Stress-Induced Growth Deficits in Livestock and Its Potential Role as a Target for Nutritional Interventions: A Review. Animals 2021, 11, 3539. https://doi.org/10.3390/ani11123539
Most MS, Yates DT. Inflammatory Mediation of Heat Stress-Induced Growth Deficits in Livestock and Its Potential Role as a Target for Nutritional Interventions: A Review. Animals. 2021; 11(12):3539. https://doi.org/10.3390/ani11123539
Chicago/Turabian StyleMost, Micah S., and Dustin T. Yates. 2021. "Inflammatory Mediation of Heat Stress-Induced Growth Deficits in Livestock and Its Potential Role as a Target for Nutritional Interventions: A Review" Animals 11, no. 12: 3539. https://doi.org/10.3390/ani11123539