
Molecular Identification and Characterization of a Genotype 3 Hepatitis E Virus (HEV) Strain Detected in a Wolf Faecal Sample, Italy

 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Molecular Screening for Orthohepeviruses
2.3. Whole Genome Sequencing
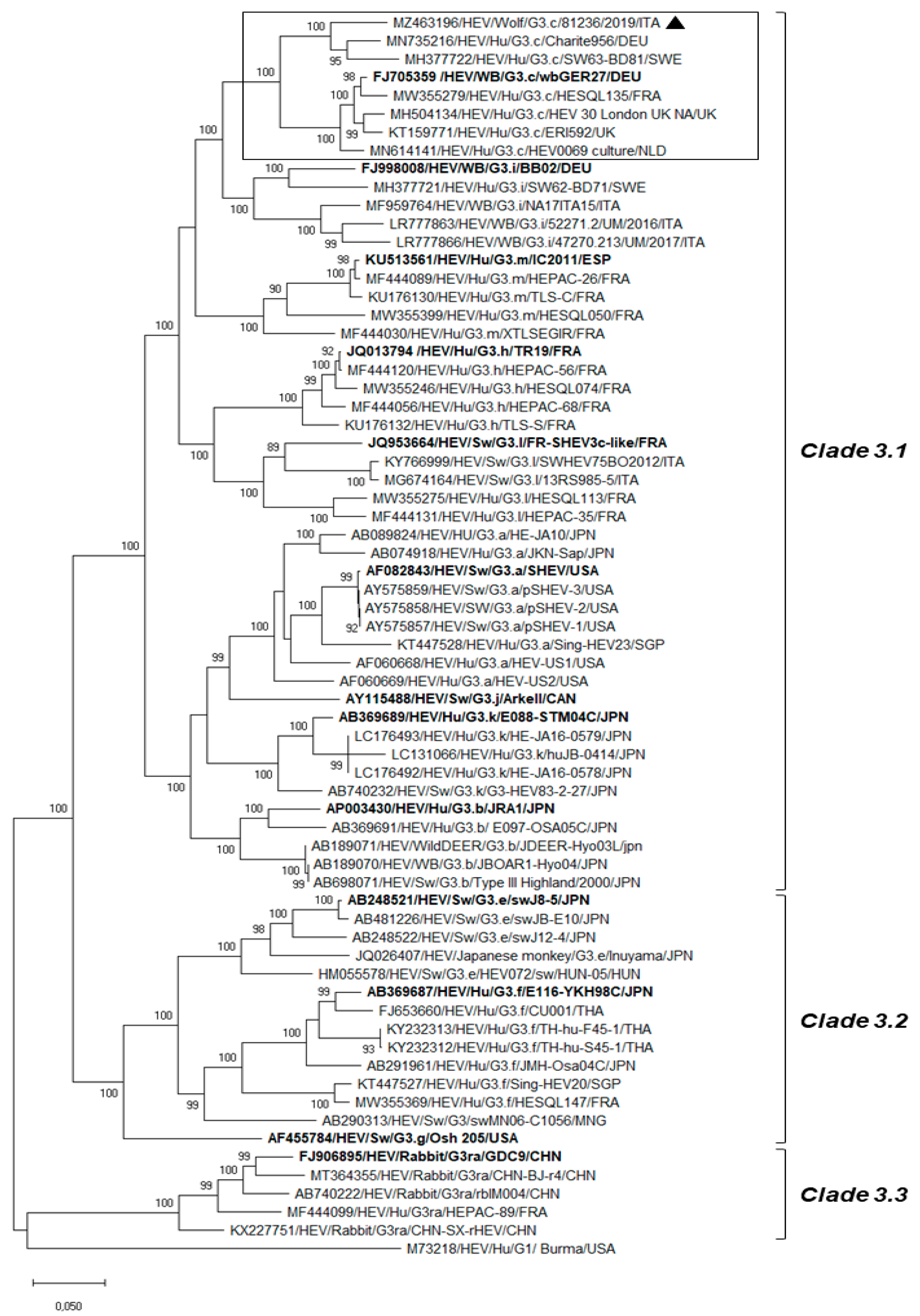
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kamar, N.; Izopet, J.; Pavio, N.; Aggarwal, R.; Labrique, A.; Wedemeyer, H.; Dalton, H.R. Hepatitis E virus infection. Nat. Rev. Dis. Prim. 2017, 3, 17086. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Meng, X.J. Structural and molecular biology of hepatitis E virus. Comput. Struct. Biotechnol. J. 2021, 19, 1907–1916. [Google Scholar] [CrossRef]
- Purdy, M.A.; Harrison, T.J.; Jameel, S.; Meng, X.J.; Okamoto, H.; Van der Poel, W.H.M.; Smith, D.B.; Ictv Report Consortium. ICTV Virus Taxonomy Profile: Hepeviridae. J. Gen. Virol. 2017, 98, 2645–2646. [Google Scholar] [CrossRef]
- Smith, D.B.; Simmonds, P.; Members of the International Committee on the Taxonomy of Viruses Study Group; Jameel, S.; Emerson, S.U.; Harrison, T.J.; Meng, X.J.; Okamoto, H.; Van der Poel, W.H.M.; Purdy, M.A. Consensus proposals for classification of the family Hepeviridae. J. Gen. Virol. 2014, 95, 2223–2232. [Google Scholar] [CrossRef]
- Smith, D.B.; Izopet, J.; Nicot, F.; Simmonds, P.; Jameel, S.; Meng, X.J.; Norder, H.; Okamoto, H.; van der Poel, W.H.M.; Reuter, G.; et al. Update: Proposed reference sequences for subtypes of hepatitis E virus (species Orthohepevirus A). J. Gen. Virol. 2020, 101, 692–698. [Google Scholar] [CrossRef]
- Pavio, N.; Doceul, V.; Bagdassarian, E.; Johne, R. Recent knowledge on hepatitis E virus in Suidae reservoirs and transmission routes to human. Vet. Res. 2017, 48, 78. [Google Scholar] [CrossRef] [Green Version]
- Kenney, S.P.; Meng, X.J. Hepatitis E Virus: Animal Models and Zoonosis. Annu. Rev. Anim. Biosci. 2019, 7, 427–448. [Google Scholar] [CrossRef]
- Nakamura, M.; Takahashi, K.; Taira, K.; Taira, M.; Ohno, A.; Sakugawa, H.; Arai, M.; Mishiro, S. Hepatitis E virus infection in wild mongooses of Okinawa, Japan: Demonstration of anti-HEV antibodies and a full-genome nucleotide sequence. Hepatol. Res. 2006, 34, 137–140. [Google Scholar] [CrossRef]
- Zhang, W.; Shen, Q.; Mou, J.; Yang, Z.B.; Yuan, C.L.; Cui, L.; Zhu, J.G.; Hua, X.G.; Xu, C.M.; Hu, J. Cross-species infection of hepatitis E virus in a zoo-like location, including birds. Epidemiol. Infect. 2008, 136, 1020–1026. [Google Scholar] [CrossRef]
- Nidaira, M.; Takahashi, K.; Ogura, G.; Taira, K.; Okano, S.; Kudaka, J.; Itokazu, K.; Mishiro, S.; Nakamura, M. Detection and phylogenetic analysis of hepatitis E viruses from mongooses in Okinawa, Japan. J. Vet. Med. Sci. 2012, 74, 1665–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raj, V.S.; Smits, S.L.; Pas, S.D.; Provacia, L.B.; Moorman-Roest, H.; Osterhaus, A.D.; Haagmans, B.L. Novel hepatitis E virus in ferrets, the Netherlands. Emerg. Infect. Dis. 2012, 18, 1369–1370. [Google Scholar] [CrossRef] [PubMed]
- Bodewes, R.; van der Giessen, J.; Haagmans, B.L.; Osterhaus, A.D.; Smits, S.L. Identification of multiple novel viruses, including a parvovirus and a hepevirus, in feces of red foxes. J. Virol. 2013, 87, 7758–7764. [Google Scholar] [CrossRef] [Green Version]
- Krog, J.S.; Breum, S.Ø.; Jensen, T.H.; Larsen, L.E. Hepatitis E virus variant in farmed mink, Denmark. Emerg. Infect. Dis. 2013, 19, 2028–2030. [Google Scholar] [CrossRef] [Green Version]
- Johne, R.; Heckel, G.; Plenge-Bönig, A.; Kindler, E.; Maresch, C.; Reetz, J.; Schielke, A.; Ulrich, R.G. Novel hepatitis E virus genotype in Norway rats, Germany. Emerg. Infect. Dis. 2010, 16, 1452–1455. [Google Scholar] [CrossRef] [Green Version]
- Eiden, M.; Dähnert, L.; Spoerel, S.; Vina-Rodriguez, A.; Schröder, R.; Conraths, F.J.; Groschup, M.H. Spatial-Temporal Dynamics of Hepatitis E Virus Infection in Foxes (Vulpes vulpes) in Federal State of Brandenburg, Germany, 1993–2012. Front. Microbiol. 2020, 11, 115. [Google Scholar] [CrossRef]
- Lanszki, Z.; Kurucz, K.; Zeghbib, S.; Kemenesi, G.; Lanszki, J.; Jakab, F. Identification of Hepatitis E Virus in the Feces of Red Foxes (Vulpes vulpes). Animals 2020, 10, 1841. [Google Scholar] [CrossRef]
- Jothikumar, N.; Cromeans, T.L.; Robertson, B.H.; Meng, X.J.; Hill, V.R. A broadly reactive one-step real-time RT-PCR assay for rapid and sensitive detection of hepatitis E virus. J. Virol. Methods. 2006, 131, 65–71. [Google Scholar] [CrossRef]
- Di Profio, F.; Melegari, I.; Sarchese, V.; Robetto, S.; Marruchella, G.; Bona, M.C.; Orusa, R.; Martella, V.; Marsilio, F.; Di Martino, B. Detection and genetic characterization of hepatitis E virus (HEV) genotype 3 subtype c in wild boars in Italy. Arch. Virol. 2016, 161, 2829–2834. [Google Scholar] [CrossRef] [PubMed]
- Drexler, J.F.; Seelen, A.; Corman, V.M.; Fumie Tateno, A.; Cottontail, V.; Melim Zerbinati, R.; Gloza-Rausch, F.; Klose, S.M.; Adu-Sarkodie, Y.; Oppong, S.K.; et al. Bats worldwide carry hepatitis E virus-related viruses that form a putative novel genus within the family Hepeviridae. J. Virol. 2012, 86, 9134–9147. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.J.; Purcell, R.H.; Halbur, P.G.; Lehman, J.R.; Webb, D.M.; Tsareva, T.S.; Haynes, J.S.; Thacker, B.J.; Emerson, S.U. A novel virus in swine is closely related to the human hepatitis E virus. Proc. Natl. Acad. Sci. USA 1997, 94, 9860–9865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes, G.R.; Kim, J.P. Sequence-independent, single-primer amplification (SISPA) of complex DNA populations. Mol. Cell. Probes 1991, 5, 473–481. [Google Scholar] [CrossRef]
- Allander, T.; Tammi, M.T.; Eriksson, M.; Bjerkner, A.; Tiveljung-Lindell, A.; Andersson, B. Cloning of a human parvovirus by molecular screening of respiratory tract samples. Proc. Natl. Acad. Sci. USA 2005, 02, 12891–12896. [Google Scholar] [CrossRef] [Green Version]
- Djikeng, A.; Halpin, R.; Kuzmickas, R.; Depasse, J.; Feldblyum, J.; Sengamalay, N.; Afonso, C.; Zhang, X.; Anderson, N.G.; Ghedin, E.; et al. Viral genome sequencing by random priming methods. BMC Genom. 2008, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Reuter, G.; Nemes, C.; Boros, A.; Kapusinszky, B.; Delwart, E.; Pankovics, P. Astrovirus in wild boars (Sus scrofa) in Hungary. Arch. Virol. 2012, 157, 1143–1147. [Google Scholar] [CrossRef]
- Norder, H.; Galli, C.; Magnil, E.; Sikora, P.; Ekvärn, E.; Nyström, K.; Magnius, L.O. Hepatitis E Virus Genotype 3 Genomes from RNA-Positive but Serologically Negative Plasma Donors Have CUG as the Start Codon for ORF3. Intervirology 2018, 61, 96–103. [Google Scholar] [CrossRef]
- Oliveira-Filho, E.F.; König, M.; Thiel, H.J. Genetic variability of HEV isolates: Inconsistencies of current classification. Vet. Microbiol. 2013, 165, 148–154. [Google Scholar] [CrossRef]
- Nicot, F.; Dimeglio, C.; Migueres, M.; Jeanne, N.; Latour, J.; Abravanel, F.; Ranger, N.; Harter, A.; Dubois, M.; Lameiras, S.; et al. Classification of the Zoonotic Hepatitis E Virus Genotype 3 Into Distinct Subgenotypes. Front. Microbiol. 2021, 11, 634430. [Google Scholar] [CrossRef]
- Meng, X.J.; Halbur, P.G.; Shapiro, M.S.; Govindarajan, S.; Bruna, J.D.; Mushahwar, I.K.; Purcell, R.H.; Emerson, S.U. Genetic and experimental evidence for cross-species infection by swine hepatitis E virus. J. Virol. 1998, 72, 9714–9721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo Presti, A.; Bruni, R.; Villano, U.; Marcantonio, C.; Equestre, M.; Ciuffetelli, M.; Grimaldi, A.; Suffredini, E.; Di Pasquale, S.; De Medici, D.; et al. Phylogenetic analysis and epidemiological history of Hepatitis E virus 3f and 3c in swine and wild boar, Italy. Heliyon 2020, 6, e05110. [Google Scholar] [CrossRef]
- Di Bartolo, I.; Ponterio, E.; Castellini, L.; Ostanello, F.; Ruggeri, F.M. Viral and antibody HEV prevalence in swine at slaughterhouse in Italy. Vet. Microbiol. 2011, 149, 330–338. [Google Scholar] [CrossRef] [Green Version]
- Serracca, L.; Battistini, R.; Rossini, I.; Mignone, W.; Peletto, S.; Boin, C.; Pistone, G.; Ercolini, R.; Ercolini, C. Molecular Investigation on the Presence of Hepatitis E Virus (HEV) in Wild Game in North-Western Italy. Food Environ. Virol. 2015, 7, 206–212. [Google Scholar] [CrossRef]
- Masotti, C.; Battistini, R.; Mignone, W.; Berio, E.; Dellepiane, M.; Andreoli, T.; Razzuoli, E.; Peletto, S.; Beltramo, C.; Acutis, P.; et al. Hepatitis E virus research in wild boars during 2017/2018 and 2018/2019 hunting season. Large Anim. Rev. 2020, 26, 149–151. [Google Scholar]
- Garbuglia, A.R.; Alessandrini, A.I.; Pavio, N.; Tessé, S.; Grignolo, S.; Viscoli, C.; Lapa, D.; Capobianchi, M.R. Male patient with acute hepatitis E in Genoa, Italy: Figatelli (pork liver sausage) as probable source of the infection. Clin. Microbiol. Infect. 2015, 21, e4–e6. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhang, W.; Shen, Q.; Yang, S.; Huang, F.; Li, P.; Guo, X.; Yang, Z.; Cui, L.; Zhu, J.; et al. Prevalence of antibody to hepatitis E virus among pet dogs in the Jiang-Zhe area of China. Scand. J. Infect. Dis. 2009, 41, 291–295. [Google Scholar] [CrossRef]
- Liang, H.; Chen, J.; Xie, J.; Sun, L.; Ji, F.; He, S.; Zheng, Y.; Liang, C.; Zhang, G.; Su, S.; et al. Hepatitis E virus serosurvey among pet dogs and cats in several developed cities in China. PLoS ONE 2014, 9, e98068. [Google Scholar] [CrossRef]
- Dähnert, L.; Conraths, F.J.; Reimer, N.; Groschup, M.H.; Eiden, M. Molecular and serological surveillance of Hepatitis E virus in wild and domestic carnivores in Brandenburg, Germany. Transbound. Emerg. Dis. 2018, 65, 1377–1380. [Google Scholar] [CrossRef]
- Capozza, P.; Martella, V.; Lanave, G.; Beikpour, F.; Di Profio, F.; Palombieri, A.; Sarchese, V.; Marsilio, F.; La Rosa, G.; Suffredini, E.; et al. A surveillance study of hepatitis E virus infection in household cats. Res. Vet. Sci. 2021, 137, 40–43. [Google Scholar] [CrossRef]
- Du, Z.Q.; Vincent-Naulleau, S.; Gilbert, H.; Vignoles, F.; Créchet, F.; Shimogiri, T.; Yasue, H.; Leplat, J.J.; Bouet, S.; Gruand, J.; et al. Detection of novel quantitative trait loci for cutaneous melanoma by genome-wide scan in the MeLiM swine model. Int. J. Cancer 2007, 120, 303–320. [Google Scholar] [PubMed]
- Di Martino, B.; Di Profio, F.; Melegari, I.; Sarchese, V.; Robetto, S.; Marsilio, F.; Martella, V. Detection of hepatitis E virus (HEV) in goats. Virus Res. 2016, 225, 69–72. [Google Scholar] [CrossRef]
- Sarchese, V.; Di Profio, F.; Melegari, I.; Palombieri, A.; Sanchez, S.B.; Arbuatti, A.; Ciuffetelli, M.; Marsilio, F.; Martella, V.; Di Martino, B. Hepatitis E virus in sheep in Italy. Transbound. Emerg. Dis. 2019, 66, 1120–1125. [Google Scholar] [CrossRef]
- Thiry, D.; Mauroy, A.; Pavio, N.; Purdy, M.A.; Rose, N.; Thiry, E.; de Oliveira-Filho, E.F. Hepatitis E Virus and Related Viruses in Animals. Transbound. Emerg. Dis. 2017, 64, 37–52. [Google Scholar] [CrossRef]


{kind=link}
| Subtype | Strain Name | ORF1 | ORF2 | ORF3 | GenBank Accession no. |
|---|---|---|---|---|---|
| 3a | Meng | 83.6% | 85.6% | 92.7% | AF082843 |
| 3b | JRA1 | 83.5% | 85.9% | 94.0% | AP003430 |
| 3c | wbGER27 | 88.8% | 90.6% | 94.0% | FJ705359 |
| 3e | swJ8-5 | 79.4% | 84.1% | 93.5% | AB248521 |
| 3f | E116-YKH98C | 79.7% | 83.8% | 94.0% | AB369687 |
| 3g | Osh 205 | 80.3% | 84.1% | 93.2% | AF455784 |
| 3h | TR19 | 84.6% | 86.7% | 94.0% | JQ013794 |
| 3i | BB02 | 85.0% | 87.8% | 95.7% | FJ998008 |
| 3j | Arkell | 82.4% | 86.0% | 93.0% | AY115488 |
| 3k | E088-STM04C | 83.6% | 86.4% | 95.1% | AB369689 |
| 3l | FR-SHEV3c-like | 84.1% | 86.2% | 93.2% | JQ953664 |
| 3m | IC2011 | 84.0% | 87.3% | 93.5% | KU513561 |
| 3ra | GDC9 | 74.9% | 81.6% | 88.1% | FJ906895 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarchese, V.; Fruci, P.; Palombieri, A.; Di Profio, F.; Robetto, S.; Ercolini, C.; Orusa, R.; Marsilio, F.; Martella, V.; Di Martino, B. Molecular Identification and Characterization of a Genotype 3 Hepatitis E Virus (HEV) Strain Detected in a Wolf Faecal Sample, Italy. Animals 2021, 11, 3465. https://doi.org/10.3390/ani11123465
Sarchese V, Fruci P, Palombieri A, Di Profio F, Robetto S, Ercolini C, Orusa R, Marsilio F, Martella V, Di Martino B. Molecular Identification and Characterization of a Genotype 3 Hepatitis E Virus (HEV) Strain Detected in a Wolf Faecal Sample, Italy. Animals. 2021; 11(12):3465. https://doi.org/10.3390/ani11123465
Chicago/Turabian StyleSarchese, Vittorio, Paola Fruci, Andrea Palombieri, Federica Di Profio, Serena Robetto, Carlo Ercolini, Riccardo Orusa, Fulvio Marsilio, Vito Martella, and Barbara Di Martino. 2021. "Molecular Identification and Characterization of a Genotype 3 Hepatitis E Virus (HEV) Strain Detected in a Wolf Faecal Sample, Italy" Animals 11, no. 12: 3465. https://doi.org/10.3390/ani11123465