
Association of Melatonin Administration in Pregnant Ewes with Growth, Redox Status and Immunity of Their Offspring

 , ,
, ,  , ,
, ,  ,
,  and
and 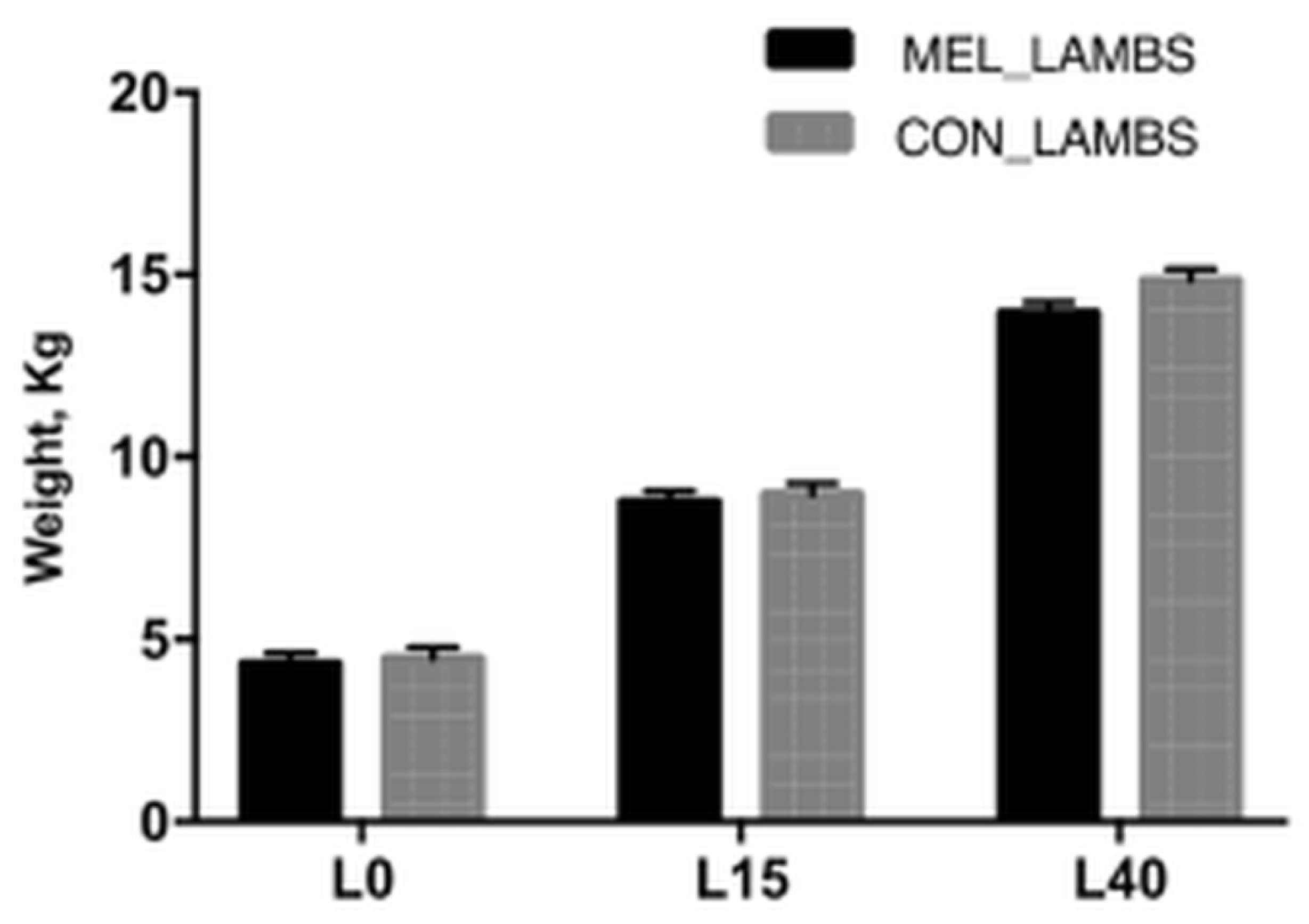{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Overview
2.2. Sampling
2.3. Blood Antioxidant Methods
2.4. Colostrum and Milk Antioxidant Methods
ABTS•+ Radical Scavenging Assay
2.5. Chemical Analysis of Colostrum and Milk
2.6. Cytokines Assays in Blood and Colostrum Samples
2.7. IgG Assay in Blood and Colostrum Samples
2.8. Statistical Analysis
3. Results
3.1. Lambs’ Body Weight Gain
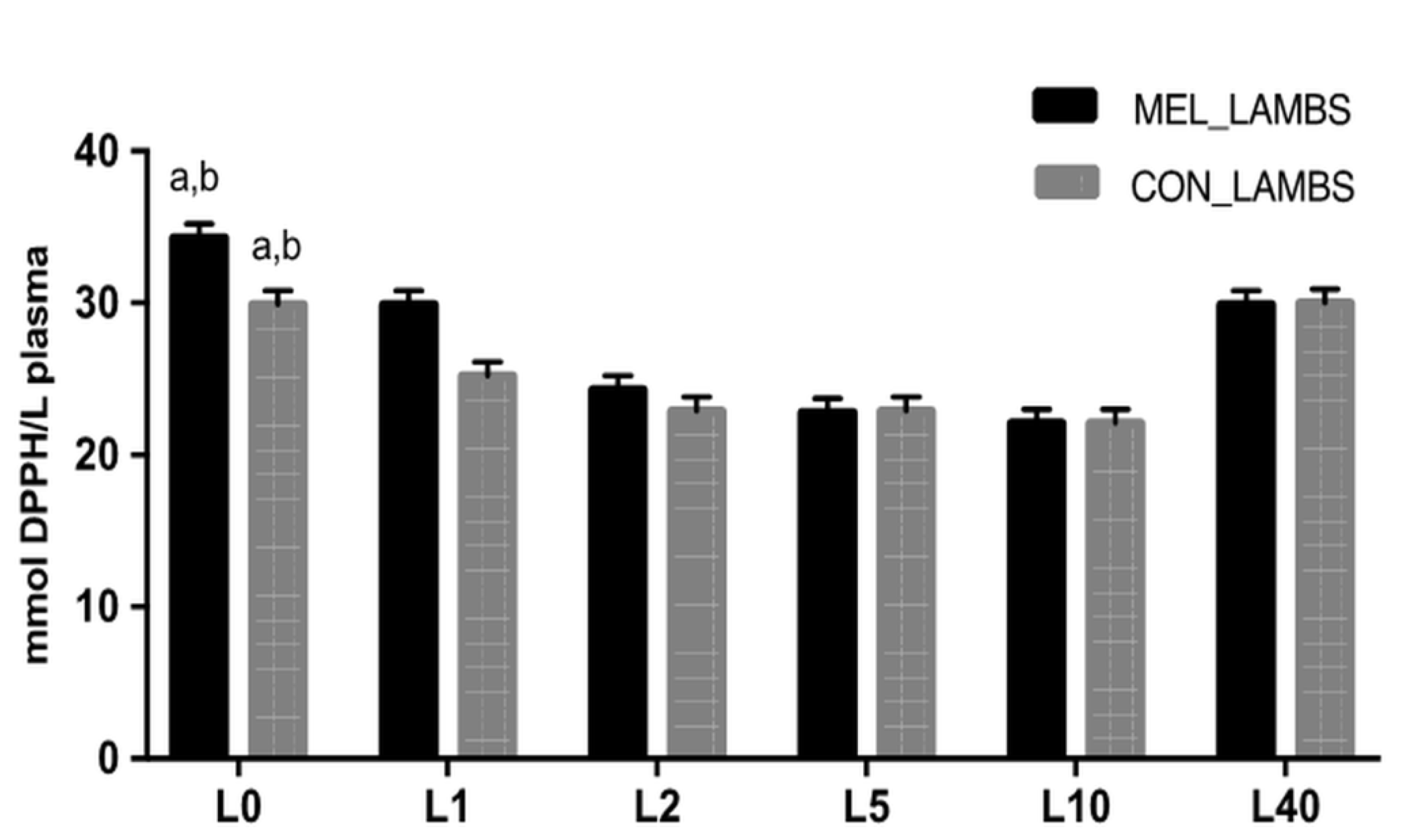
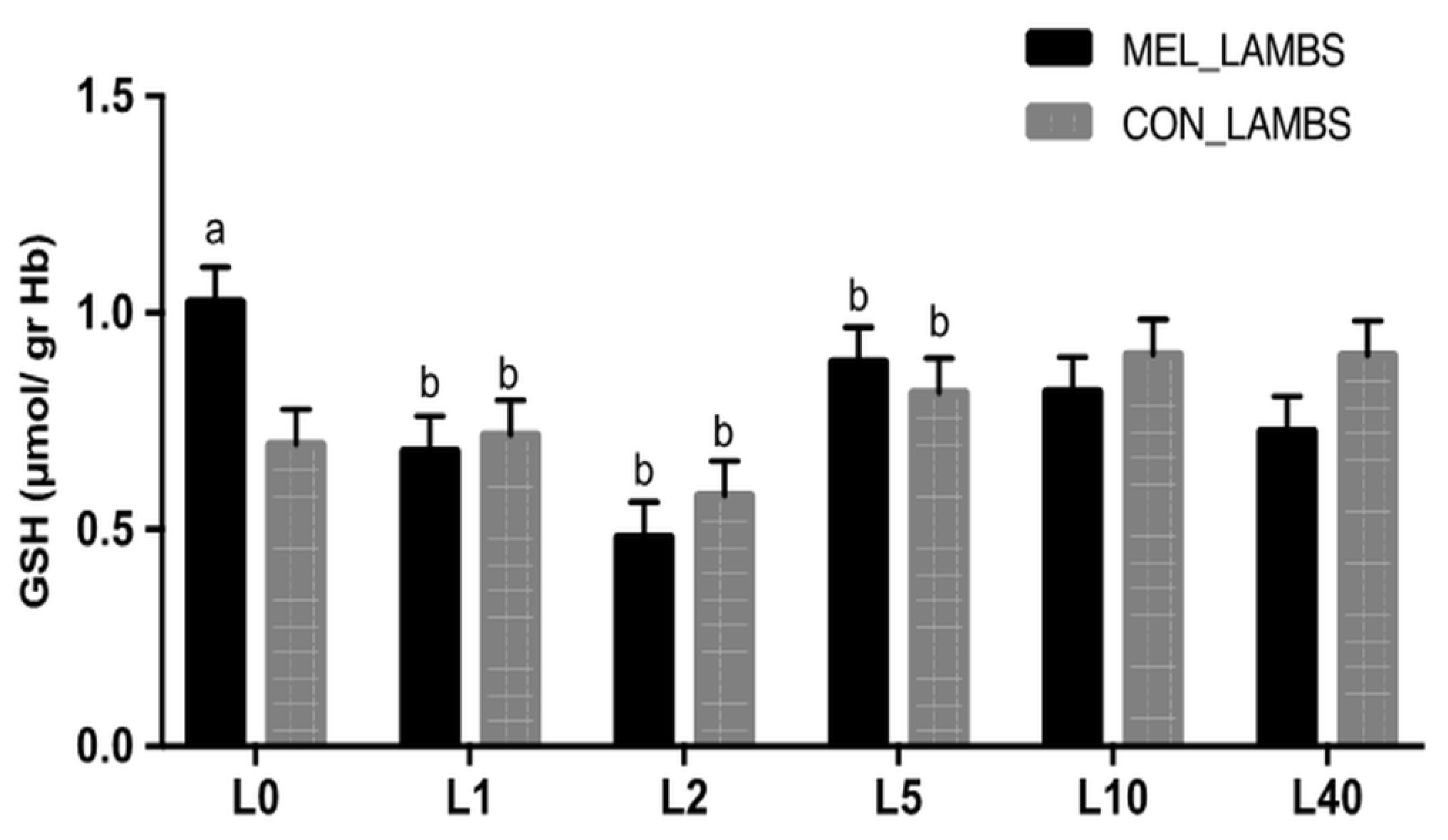
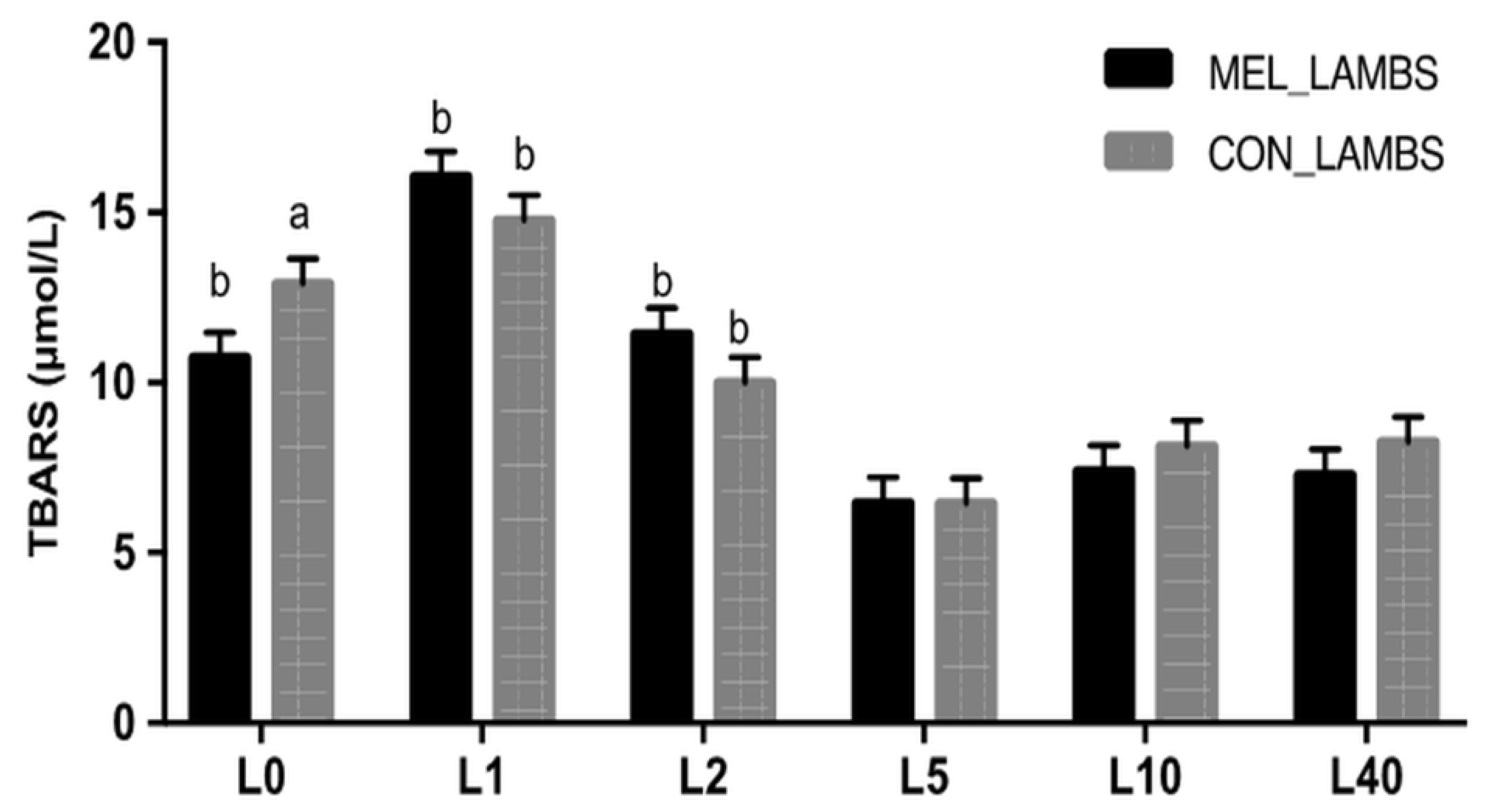
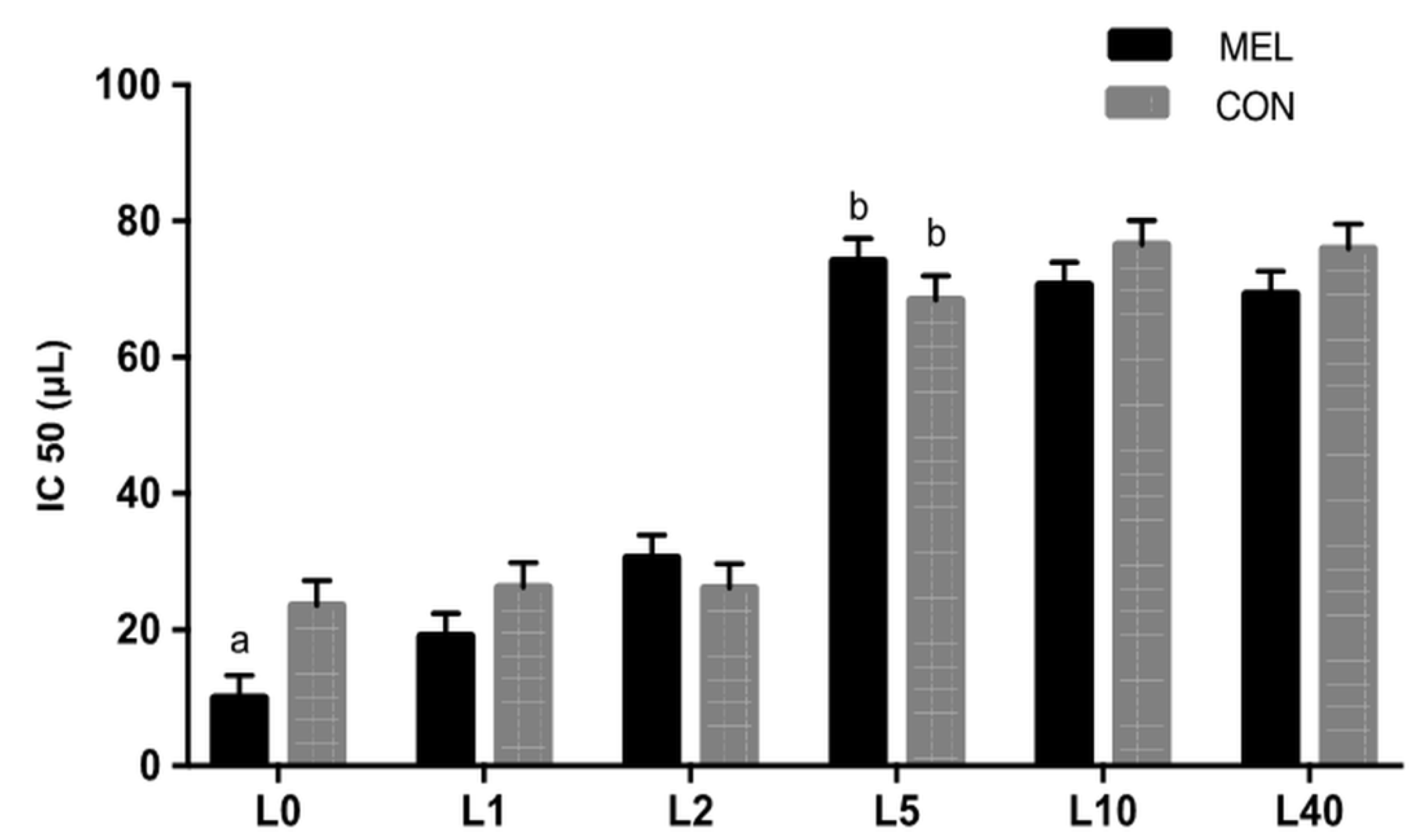
3.2. Antioxidant Biomarkers in Blood
3.3. Antioxidant Biomarkers in Colostrum/Milk Samples
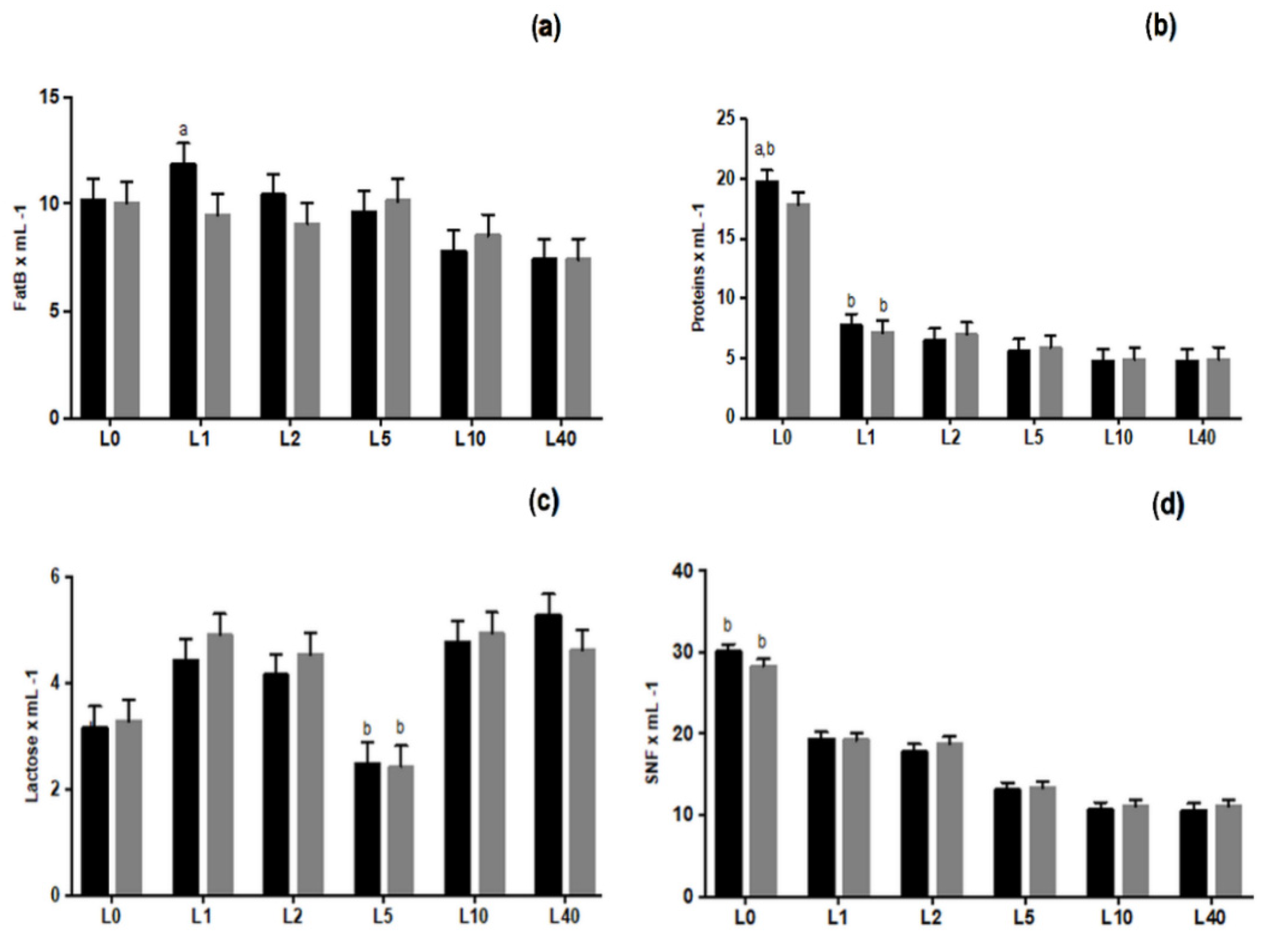
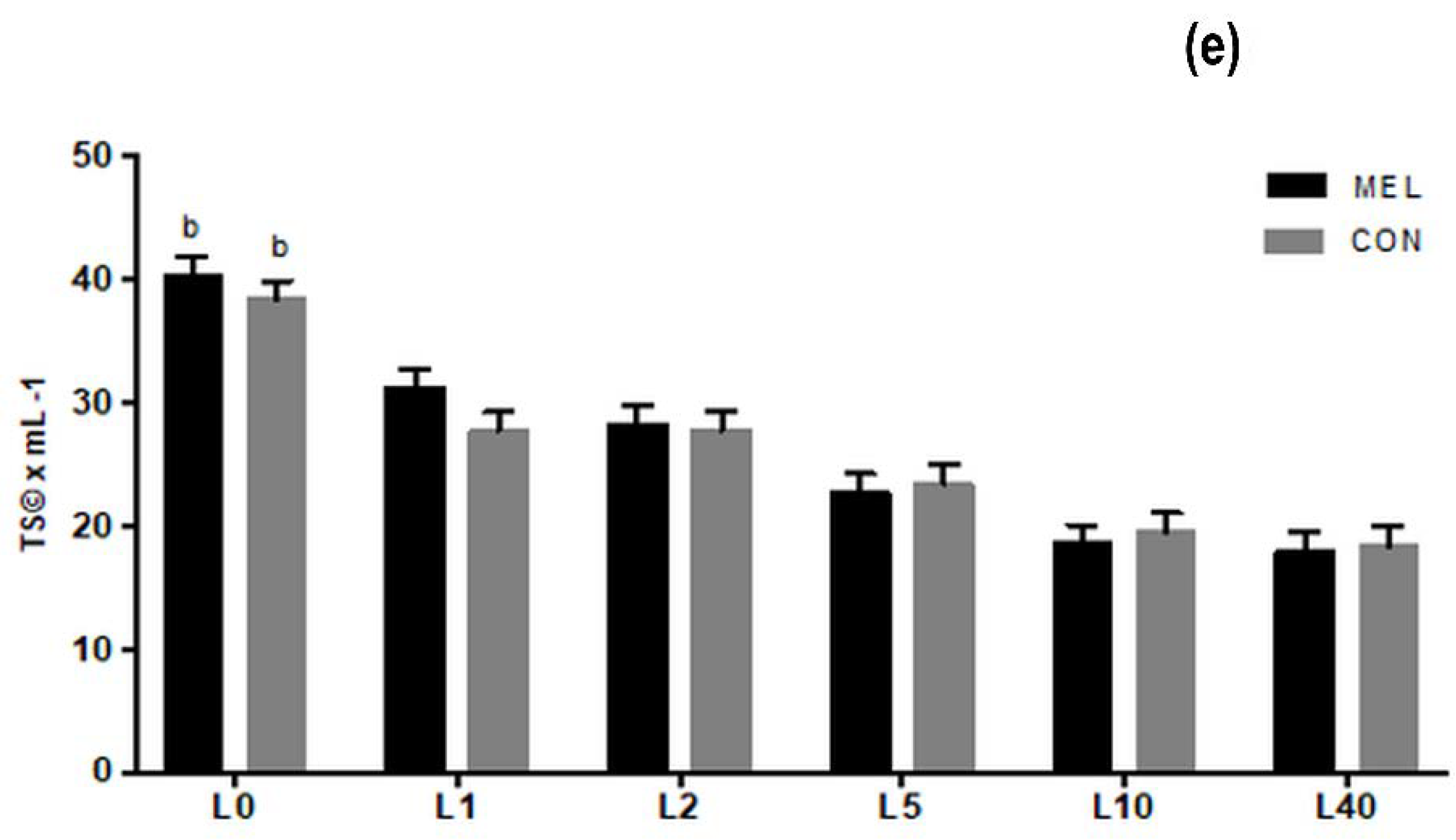
3.4. Chemical Analysis of Colostrum and Milk Samples
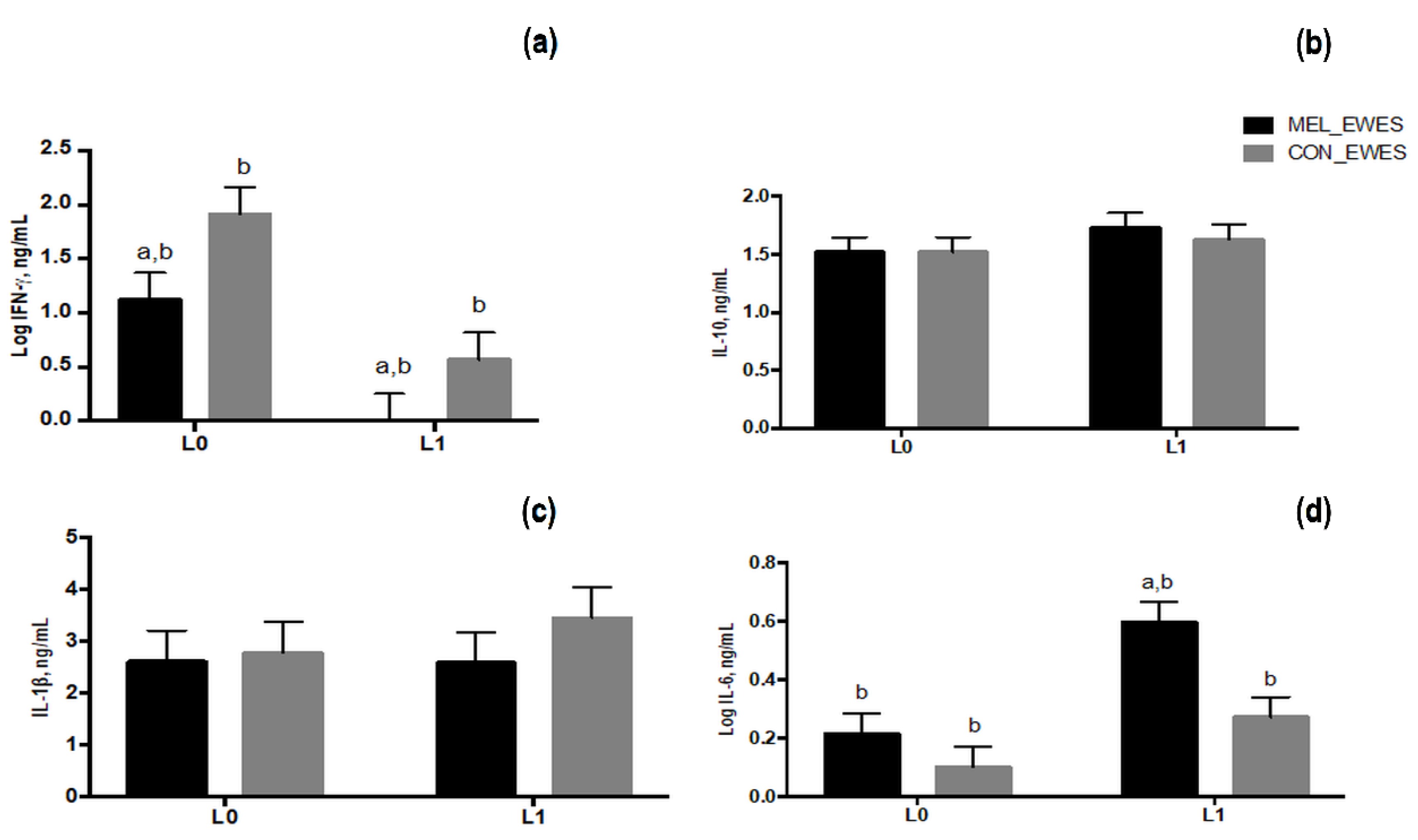
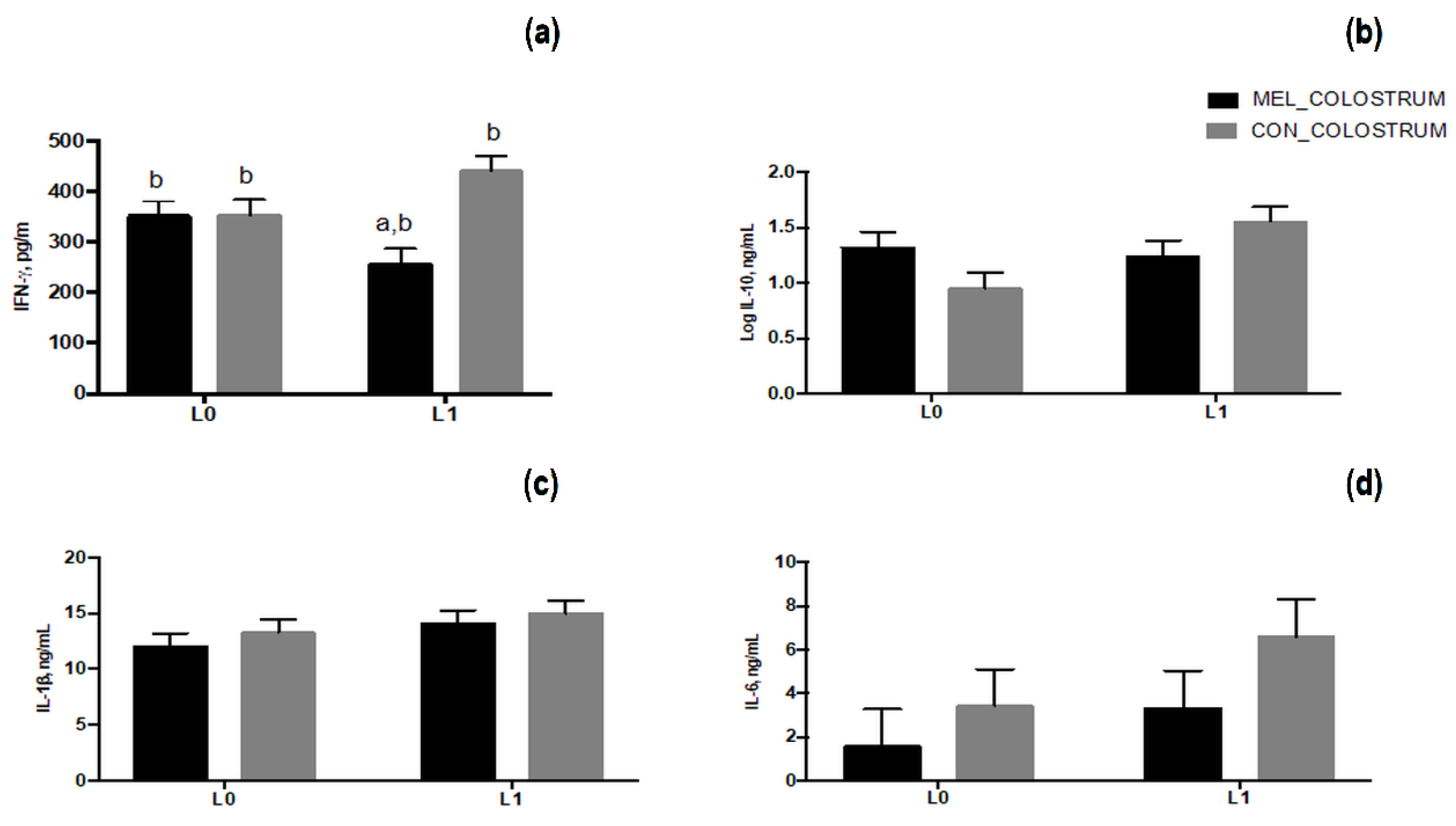
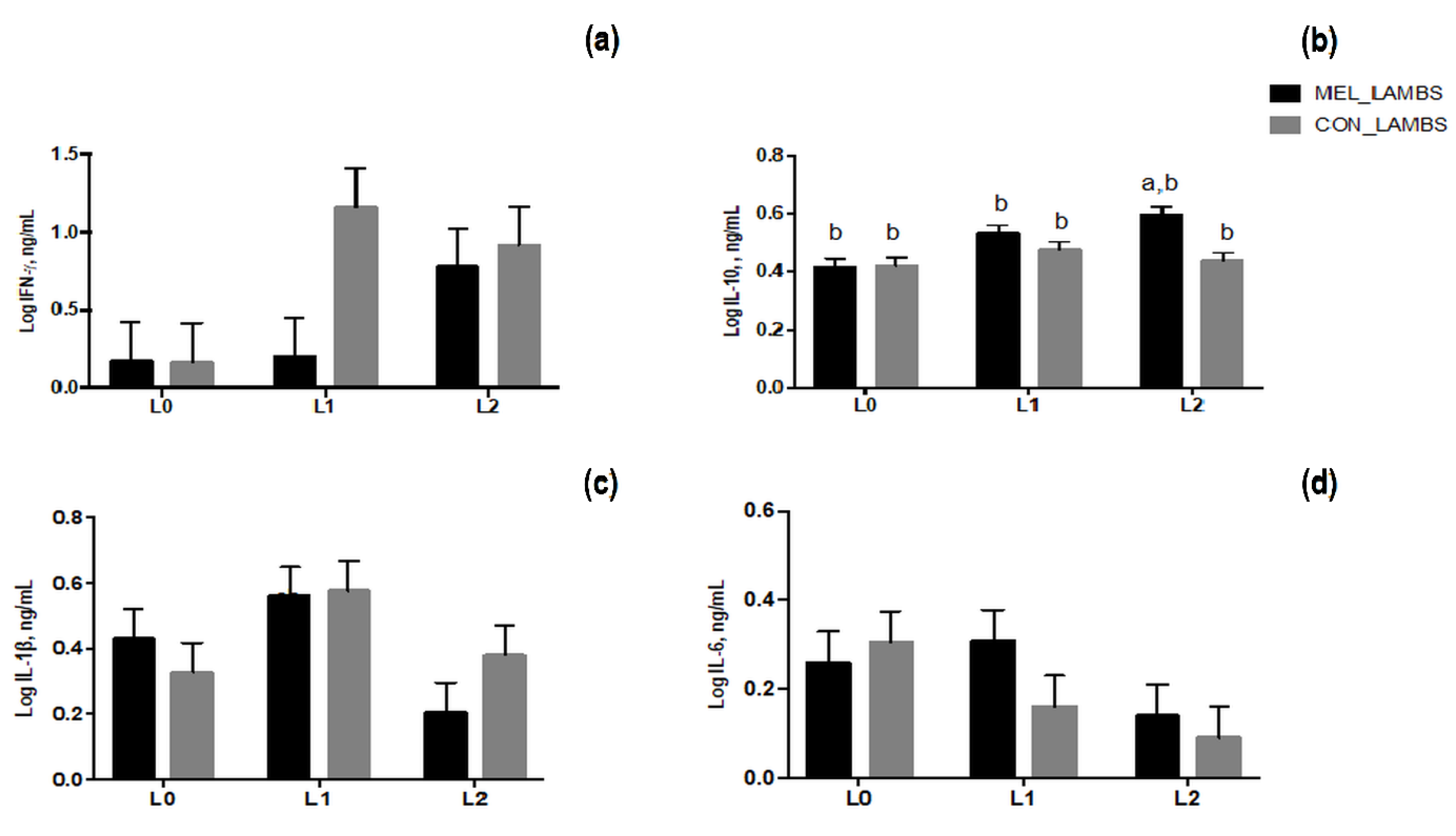
3.5. Cytokines Secretion in Blood of Ewes and Lambs and in Colostrum Samples
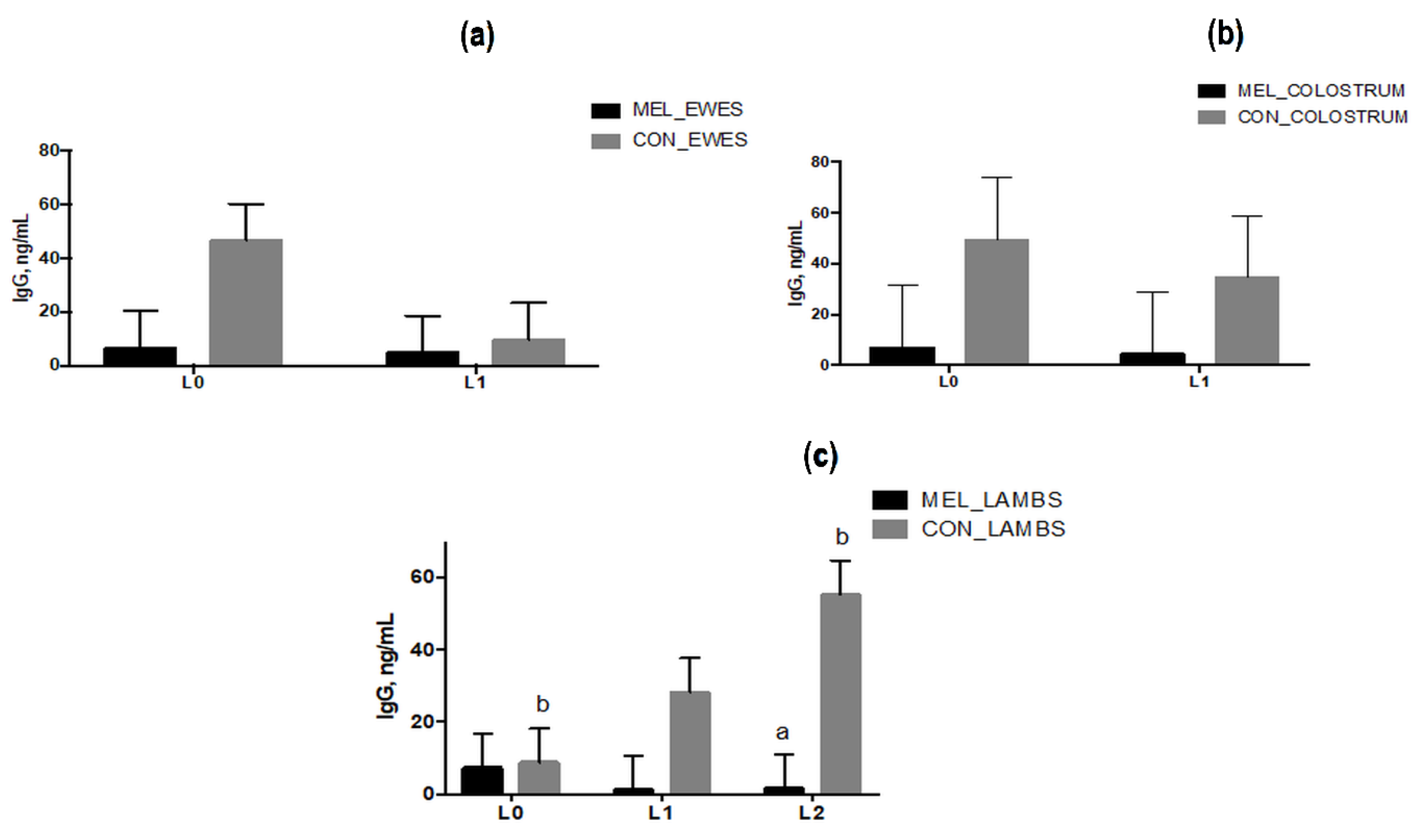
3.6. IgG Levels in Blood and Colostrum Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sinclair, K.D.; Rutherford, K.; Wallace, J.; Brameld, J.; Stöger, R.; Alberio, R.; Sweetman, D.; Gardner, D.S.; Perry, V.; Adam, C.L.; et al. Epigenetics and developmental programming of welfare and production traits in farm animals. Reprod. Fertil. Dev. 2016, 28, 1443–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartol, F.F.; Wiley, A.A.; Miller, D.J.; Silva, A.J.; Roberts, K.E.; Davolt, M.L.P.; Chen, J.C.; Frankshun, A.-L.; Camp, M.E.; Rahman, K.M.; et al. Lactation Biology Symposium: Lactocrine signaling and developmental programming1,2. J. Anim. Sci. 2013, 91, 696–705. [Google Scholar] [CrossRef]
- Chadio, S.; Kotsampasi, B. The role of early life nutrition in programming of reproductive function. J. Dev. Orig. Health Dis. 2014, 5, 2–15. [Google Scholar] [CrossRef]
- Spencer, S.J.; Galic, M.A.; Pittman, Q.J. Neonatal programming of innate immune function. Am. J. Physiol. Endocrinol. Metab. 2011, 300, E11–E18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merlot, E.; Couret, D.; Otten, W. Prenatal stress, fetal imprinting and immunity. Brain Behav. Immun. 2008, 22, 42–51. [Google Scholar] [CrossRef]
- Tao, J.; Lv, J.; Li, W.; Zhang, P.; Mao, C.; Xu, Z. Exogenous melatonin reduced blood pressure in late-term ovine fetus via MT1/MT2 receptor pathways. Reprod. Biol. 2016, 16, 212–217. [Google Scholar] [CrossRef]
- Mutinati, M.; Piccinno, M.; Roncetti, M.; Campanile, D.; Rizzo, A.; Sciorsci, R. Oxidative Stress during Pregnancy In The Sheep. Reprod. Domest. Anim. Zuchthyg. 2013, 48, 353–357. [Google Scholar] [CrossRef]
- Bouroutzika, E.; Kouretas, D.; Papadopoulos, S.; Veskoukis, A.S.; Theodosiadou, E.; Makri, S.; Paliouras, C.; Michailidis, M.-L.; Caroprese, M.; Valasi, I. Effects of Melatonin Administration to Pregnant Ewes under Heat-Stress Conditions, in Redox Status and Reproductive Outcome. Antioxidants 2020, 9, 266. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-C.; Sheen, J.-M.; Tiao, M.-M.; Tain, Y.-L.; Huang, L.-T. Roles of Melatonin in Fetal Programming in Compromised Pregnancies. Int. J. Mol. Sci. 2013, 14, 5380–5401. [Google Scholar] [CrossRef] [Green Version]
- Pevet, P.; Challet, E. Melatonin: Both master clock output and internal time-giver in the circadian clocks network. J. Physiol. 2011, 105, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Gomes, P.R.L.; Motta-Teixeira, L.C.; Gallo, C.C.; Carmo Buonfiglio, D.D.; Camargo, L.S.; Quintela, T.; Reiter, R.J.; Amaral, F.G.D.; Cipolla-Neto, J. Maternal pineal melatonin in gestation and lactation physiology, and in fetal development and programming. Gen. Comp. Endocrinol. 2021, 300, 113633. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.-X.; Fuentes-Broto, L. Melatonin: A Multitasking Molecule. Prog. Brain Res. 2010, 181, 127–151. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Calvo, J.R.; Karbownik, M.; Qi, W.; Tan, D.X. Melatonin and Its Relation to the Immune System and Inflammation. Ann. N. Y. Acad. Sci. 2000, 917, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Radogna, F.; Diederich, M.; Ghibelli, L. Melatonin: A pleiotropic molecule regulating inflammation. Biochem. Pharmacol. 2010, 80, 1844–1852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardeland, R.; Cardinali, D.P.; Srinivasan, V.; Spence, D.W.; Brown, G.M.; Pandi-Perumal, S.R. Melatonin—A pleiotropic, orchestrating regulator molecule. Prog. Neurobiol. 2011, 93, 350–384. [Google Scholar] [CrossRef] [Green Version]
- Mauriz, J.L.; Collado, P.S.; Veneroso, C.; Reiter, R.J.; González-Gallego, J. A review of the molecular aspects of melatonin’s anti-inflammatory actions: Recent insights and new perspectives. J. Pineal Res. 2013, 54, 1–14. [Google Scholar] [CrossRef]
- Carpentieri, A.; Diaz de Barboza, G.; Areco, V.; Peralta Lopez, M.; Tolosa de Talamoni, N. New perspectives in melatonin uses. Pharmacol. Res. 2012, 65, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Okatani, Y.; Okamoto, K.; Hayashi, K.; Wakatsuki, A.; Tamura, S.; Sagara, Y. Maternal-fetal transfer of melatonin in pregnant women near term. J. Pineal Res. 1998, 25, 129–134. [Google Scholar] [CrossRef]
- McMillen, I.C.; Houghton, D.C.; Young, I.R. Melatonin and the development of circadian and seasonal rhythmicity. J. Reprod. Fertil. Suppl. 1995, 49, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Deguchi, T. Ontogenesis of a biological clock for serotonin:acetyl coenzyme A N-acetyltransferase in pineal gland of rat. Proc. Natl. Acad. Sci. USA 1975, 72, 2814–2818. [Google Scholar] [CrossRef] [Green Version]
- Kivelä, A. Serum melatonin during human pregnancy. Acta Endocrinol. 1991, 124, 233–237. [Google Scholar] [CrossRef]
- Pang, S.F.; Tang, P.L.; Tang, G.W.K.; Yam, A.W.C.; Ng, K.W. Plasma levels of immunoreactive melatonin, estradiol, progesterone, follicle stimulating hormone, and beta-human chorionic gonadotropin during pregnancy and shortly after parturition in humans. J. Pineal Res. 1987, 4, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Wierrani, F.; Grin, W.; Hlawka, B.; Kroiss, A. Elevated serum melatonin levels during human late pregnancy and labour. J. Obstet. Gynaecol. 1997, 17, 449–451. [Google Scholar] [CrossRef]
- Tain, Y.-L.; Huang, L.-T.; Hsu, C.-N. Developmental Programming of Adult Disease: Reprogramming by Melatonin? Int. J. Mol. Sci. 2017, 18, 426. [Google Scholar] [CrossRef] [Green Version]
- Greenwood, P.L.; Hunt, A.S.; Hermanson, J.W.; Bell, A.W. Effects of birth weight and postnatal nutrition on neonatal sheep: II. Skeletal muscle growth and development. J. Anim. Sci. 2000, 78, 50–61. [Google Scholar] [CrossRef] [Green Version]
- Przybylska, J.; Albera, E.; Kankofer, M. Antioxidants in Bovine Colostrum. Reprod. Domest. Anim. Zuchthyg. 2007, 42, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Marta, K.; Lipko-Przybylska, J. Physiological antioxidative/oxidative status in bovine colostrum and mature milk. Acta Vet. 2008, 58, 231–239. [Google Scholar] [CrossRef]
- Margiloff, L.; Chaplia, L.; Chow, A.; Singhal, P.; Mattana, J. Metal-catalyzed oxidation of immunoglobulin G impairs Fc receptor-mediated binding to macrophages. Free. Radic. Biol. Med. 1998, 25, 780–785. [Google Scholar] [CrossRef]
- Abuelo, Á.; Pérez-Santos, M.; Hernández, J.; Castillo, C. Effect of colostrum redox balance on the oxidative status of calves during the first 3months of life and the relationship with passive immune acquisition. Vet. J. 2014, 199, 295–299. [Google Scholar] [CrossRef] [Green Version]
- Veskoukis, A.S.; Kerasioti, E.; Priftis, A.; Kouka, P.; Spanidis, Y.; Makri, S.; Kouretas, D. A battery of translational biomarkers for the assessment of the in vitro and in vivo antioxidant action of plant polyphenolic compounds: The biomarker issue. Curr. Opin. Toxicol. 2019, 13, 99–109. [Google Scholar] [CrossRef]
- Veskoukis, A.S.; Nikolaidis, M.G.; Kyparos, A.; Kokkinos, D.; Nepka, C.; Barbanis, S.; Kouretas, D. Effects of xanthine oxidase inhibition on oxidative stress and swimming performance in rats. Appl. Physiol. Nutr. Metab. 2008, 33, 1140–1154. [Google Scholar] [CrossRef]
- Veskoukis, A.S.; Kyparos, A.; Nikolaidis, M.G.; Stagos, D.; Aligiannis, N.; Halabalaki, M.; Chronis, K.; Goutzourelas, N.; Skaltsounis, L.; Kouretas, D. The Antioxidant Effects of a Polyphenol-Rich Grape Pomace ExtractIn VitroDo Not CorrespondIn VivoUsing Exercise as an Oxidant Stimulus. Oxidative Med. Cell. Longev. 2012, 2012, 185867. [Google Scholar] [CrossRef] [Green Version]
- Veskoukis, A.S.; Kyparos, A.; Paschalis, V.; Nikolaidis, M.G. Spectrophotometric assays for measuring redox biomarkers in blood. Biomarkers 2016, 21, 208–217. [Google Scholar] [CrossRef]
- Cano, A.; Hernández-Ruíz, J.; García-Cánovas, F.; Acosta, M.; Arnao, M.B. An end-point method for esti-mation of the total antioxidant activity in plant material. Phytochem. Anal. 1998, 9, 196–202. [Google Scholar] [CrossRef]
- Kwong, L.; Hope, J.; Thom, M.; Sopp, P.; Duggan, S.; Bembridge, G.; Howard, C. Development of an ELISA for bovine IL-10. Vet. Immunol. Immunopathol. 2002, 85, 213–223. [Google Scholar] [CrossRef]
- Ciliberti, M.; Albenzio, M.; Inghese, C.; Santillo, A.; Marino, R.; Sevi, A.; Caroprese, M. Peripheral blood mononuclear cell proliferation and cytokine production in sheep as affected by cortisol level and duration of stress. J. Dairy Sci. 2017, 100, 750–756. [Google Scholar] [CrossRef] [Green Version]
- Ciliberti, M.G.; Albenzio, M.; De Palo, P.; Santillo, A.; Caroprese, M. Nexus Between Immune Responses and Oxidative Stress: The Role of Dietary Hydrolyzed Lignin in ex vivo Bovine Peripheral Blood Mononuclear Cell Response. Front. Vet. Sci. 2020, 7, 9. [Google Scholar] [CrossRef]
- Hardeland, R.; Tan, D.-X.; Reiter, R.J. Kynuramines, metabolites of melatonin and other indoles: The resurrection of an almost forgotten class of biogenic amines. J. Pineal Res. 2009, 47, 109–126. [Google Scholar] [CrossRef]
- Reiter, R.J.; Korkmaz, A.; Paredes, S.D.; Manchester, L.C.; Tan, D.-X. Melatonin reduces oxidative/nitrosative stress due to drugs, toxins, metals, and herbicides. Neuro Endocrinol. Lett. 2008, 29, 609–613. [Google Scholar] [PubMed]
- Reiter, R.J.; Tan, D.-X.; Burkhardt, S. Reactive oxygen and nitrogen species and cellular and organismal decline: Amelioration with melatonin. Mech. Ageing Dev. 2002, 123, 1007–1019. [Google Scholar] [CrossRef]
- Marai, I.; El-Darawany, A.; Fadiel, A.; Abdel-Hafez, M. Physiological traits as affected by heat stress in sheep—A review. Small Rumin. Res. 2007, 71, 1–12. [Google Scholar] [CrossRef]
- Davis, F.C.; Mannion, J. Entrainment of hamster pup circadian rhythms by prenatal melatonin injections to the mother. Am. J. Physiol. 1988, 255, R439–R448. [Google Scholar] [CrossRef]
- Klein, D. Evidence for the Placental Transfer of 3H-Acetyl-Melatonin. Nat. New Biol. 1972, 237, 117–118. [Google Scholar] [CrossRef]
- Vermouth, N.T.; Carriazo, C.S.; Gallará, R.V.; Carpentieri, A.R.; Bellavía, S.L. Maternal Coordination of the Daily Rhythm of Malate Dehydrogenase Activity in Testes from Young Rats: Effect of Maternal Sympathetic Denervation of the Pineal Gland and Administration of Melatonin. Chronobiol. Int. 1995, 12, 8–18. [Google Scholar] [CrossRef]
- Tan, D.-X.; Manchester, L.C.; Reiter, R.J.; Qi, W.-B.; Karbownik, M.; Calvo, J.R. Significance of Melatonin in Antioxidative Defense System: Reactions and Products. Biol. Signals Recept. 2000, 9, 137–159. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, A.; Reiter, R.J. Epigenetic regulation: A new research area for melatonin? J. Pineal Res. 2008, 44, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Scumaci, D.; Trimboli, F.; Dell’Aquila, L.; Concolino, A.; Pappaianni, G.; Tammè, L.; Vignola, G.; Luciani, A.; Morelli, D.; Cuda, G.; et al. Proteomics-Driven Analysis of Ovine Whey Colostrum. PLoS ONE 2015, 10, e0117433. [Google Scholar] [CrossRef] [PubMed]
- Bauman, D.E.; Mather, I.H.; Wall, R.J.; Lock, A.L. Major Advances Associated with the Biosynthesis of Milk. J. Dairy Sci. 2006, 89, 1235–1243. [Google Scholar] [CrossRef]
- Abecia, J.; Garrido, C.; Gave, M.; García, A.; López, D.; Luis, S.; Valares, J.; Mata, L. Exogenous melatonin and male foetuses improve the quality of sheep colostrum. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1305–1309. [Google Scholar] [CrossRef] [PubMed]
- Molik, E.; Błasiak, M.; Pustkowiak, H. Impact of Photoperiod Length and Treatment with Exogenous Melatonin during Pregnancy on Chemical Composition of Sheep’s Milk. Animals 2020, 10, 1721. [Google Scholar] [CrossRef]
- Barker, D.J. The developmental origins of well–being. Philos. Trans. R. Soc. B Biol. Sci. 2004, 359, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, L.P.; Borowicz, P.P.; Caton, J.S.; Vonnahme, K.A.; Luther, J.S.; Hammer, C.J.; Maddock Carlin, K.R.; Grazulbilska, A.T.; Redmer, D.A. Developmental programming: The concept, large animal models, and the key role of uteroplacental vascular development1,2. J. Anim. Sci. 2010, 88, E61–E72. [Google Scholar] [CrossRef]
- Gonzalez-Candia, A.; Veliz, M.; Araya, C.; Quezada, S.; Ebensperger, G.; Serón-Ferré, M.; Reyes, R.V.; Llanos, A.J.; Herrera, E.A. Potential adverse effects of antenatal melatonin as a treatment for intrauterine growth restriction: Findings in pregnant sheep. Am. J. Obstet. Gynecol. 2016, 215, 245.e1–245.e7. [Google Scholar] [CrossRef] [PubMed]
- Halliday, R. Immunity and health in young lambs. Vet. Rec. 1978, 103, 489–492. [Google Scholar] [CrossRef]
- Campbell, S.; Siegel, M.; Knowlton, B.J. Sheep immunoglobulins and their transmission to the neonatal lamb. N. Zeal. Vet. J. 1977, 25, 361–365. [Google Scholar] [CrossRef]
- Britti, D.; Massimini, G.; Peli, A.; Luciani, A.; Boari, A. Evaluation of serum enzyme activities as predictors of passive transfer status in lambs. J. Am. Vet. Med. Assoc. 2005, 226, 951–955. [Google Scholar] [CrossRef]
- Rousset, F.; Garcia, E.; Defrance, T.; Peronne, C.; Vezzio, N.; Hsu, D.H.; Kastelein, R.; Moore, K.W.; Banchereau, J. Interleukin 10 is a potent growth and differentiation factor for activated human B lymphocytes. Proc. Natl. Acad. Sci. USA 1992, 89, 1890–1893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doyle, R.J.; Hirst, D.M.; Hutson, J.M. Ab initio potential energy surfaces, bound states, and electronic spectrum of the Ar-SH complex. J. Chem. Phys. 2006, 125, 184312. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Shi, J.; Tian, J.; Tao, J.; Chai, M.; Wang, J.; Xu, Z.; Song, Y.; Zhu, K.; Ji, P.; et al. Exogenous melatonin reduces somatic cell count of milk in Holstein cows. Sci. Rep. 2017, 7, 43280. [Google Scholar] [CrossRef] [Green Version]
- Cosso, G.; Mura, M.C.; Pulinas, L.; Curone, G.; Vigo, D.; Carcangiu, V.; Luridiana, S. Effects of melatonin treatment on milk traits, reproductive performance and immune response in Sarda dairy sheep. Ital. J. Anim. Sci. 2021, 20, 632–639. [Google Scholar] [CrossRef]
- Carrillo-Vico, A.; Reiter, R.J.; Lardone, P.J.; Herrera, J.L.; Fernandez-Montesinos, R.; Guerrero, J.M.; Pozo, D. The modulatory role of melatonin on immune responsiveness. Curr. Opin. Investig. Drugs 2006, 7, 423–431. [Google Scholar]
- Bernard, S.; Macedo, N.; Malpaux, B.; Chemineau, P. Comparison of immune parameters of sheep with naturally high or low plasma concentrations of melatonin. J. Pineal Res. 2001, 31, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Parker, R.J.; Nicol, A.M. The measurement of serum immunoglobulin concentration to estimate lamb colostrum intake. Proc. N. Z. Soc. Anim. Prod. 1990, 50, 275–278. Available online: https://www.nzsap.org/system/files/proceedings/1990/ab90039.pdf (accessed on 31 October 2021).
- Kamada, H.; Nonaka, I.; Ueda, Y.; Murai, M. Selenium Addition to Colostrum Increases Immunoglobulin G Absorption by Newborn Calves. J. Dairy Sci. 2007, 90, 5665–5670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kehrli, M.E.; Harp, J.A. Immunity in the Mammary Gland. Vet. Clin. N. Am. Food Anim. Pract. 2001, 17, 495–516. [Google Scholar] [CrossRef]
- Kimura, K.; Goff, J.P.; Kehrli, M.E. Effects of the Presence of the Mammary Gland on Expression of Neutrophil Adhesion Molecules and Myeloperoxidase Activity in Periparturient Dairy Cows. J. Dairy Sci. 1999, 82, 2385–2392. [Google Scholar] [CrossRef]
- Caroprese, M.; Albenzio, M.; Annicchiarico, G.; Sevi, A. Changes Occurring in Immune Responsiveness of Single- and Twin-Bearing Comisana Ewes during the Transition Period. J. Dairy Sci. 2006, 89, 562–568. [Google Scholar] [CrossRef]
- Theodorou, G.; Fragou, S.; Chronopoulou, R.; Kominakis, A.; Rogdakis, E.; Politis, I. Short Communication: Study of Immune Parameters in Three Greek Dairy Sheep Breeds During the Periparturient Period. J. Dairy Sci. 2007, 90, 5567–5571. [Google Scholar] [CrossRef]
- Kehrli, M.E., Jr.; Nonnecke, B.J.; Roth, J.A. Alterations in bovine neutrophil function during the periparturi-ent period. Am. J. Vet. Res. 1989, 50, 207–214. [Google Scholar]
- Mallard, B.A.; Wagter, L.C.; Ireland, M.J.; Dekkers, J.C. Effects of growth hormone, insulin-like growth factor-I, and cortisol on periparturient antibody response profiles of dairy cattle. Vet. Immunol. Immunopathol. 1997, 60, 61–76. [Google Scholar] [CrossRef]
- Rothel, J.; Hurst, L.; Seow, H.F.; Pepin, M.; Berthon, P.; Corner, L.A.; Wood, P.R. Analysis of ovine IL-1β production in vivo and in vitro by enzyme immunoassay and immunohistochemistry. Vet. Immunol. Immunopathol. 1997, 57, 267–278. [Google Scholar] [CrossRef]
- Yoshioka, M.; Watanabe, A.; Shimada, N.; Murata, H.; Yokomizo, Y.; Nakajima, Y. Regulation of haptoglobin secretion by recombinant bovine cytokines in primary cultured bovine hepatocytes. Domest. Anim. Endocrinol. 2002, 23, 425–433. [Google Scholar] [CrossRef]
- McWaters, P.; Hurst, L.; Chaplin, P.J.; A Collins, R.; Wood, P.R.; Scheerlinck, J.-P. Characterisation of monoclonal antibodies to ovine interleukin-6 and the development of a sensitive capture ELISA. Vet. Immunol. Immunopathol. 2000, 73, 155–165. [Google Scholar] [CrossRef]
- Murata, H.; Shimada, N.; Yoshioka, M. Current research on acute phase proteins in veterinary diagnosis: An overview. Vet. J. 2004, 168, 28–40. [Google Scholar] [CrossRef]
- Winter, P.; Colditz, I. Immunological responses of the lactating ovine udder following experimental challenge with Staphylococcus epidermidis. Vet. Immunol. Immunopathol. 2002, 89, 57–65. [Google Scholar] [CrossRef]
- Lust, J.A.; Donovan, K.A.; Kline, M.P.; Greipp, P.R.; Kyle, R.A.; Maihle, N.J. Isolation of an mRNA encoding a soluble form of the human interleukin-6 receptor. Cytokine 1992, 4, 96–100. [Google Scholar] [CrossRef]
- Caroprese, M.; Ciliberti, M.G.; Albenzio, M.; Annicchiarico, G.; Sevi, A. Dietary polyunsaturated fatty acids from flaxseed affect immune responses of dairy sheep around parturition. Vet. Immunol. Immunopathol. 2015, 168, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Fernández, A.; Marteles, D.; de Arcaute, M.R.; Lacasta, D.; Conde, T.; Loste, A. Relationship between Pro-Inflammatory Cytokines, IL-10 Anti-Inflammatory Cytokine and Serum Proteins in Healthy Lambs and with Diarrhea. Pak. Vet. J. 2016, 36, 63–67. [Google Scholar]
- Mocellin, S.; Marincola, F.; Rossi, C.R.; Nitti, D.; Lise, M. The multifaceted relationship between IL-10 and adaptive immunity: Putting together the pieces of a puzzle. Cytokine Growth Factor Rev. 2004, 15, 61–76. [Google Scholar] [CrossRef]
- Chadio, S.; Katsafadou, A.; Kotsampasi, B.; Michailidis, G.; Mountzouris, K.C.; Kalogiannis, D.; Christodoulou, V.; Mantzaris, G.J. Effects of maternal undernutrition during late gestation and/or lactation on colostrum synthesis and immunological parameters in the offspring. Reprod. Fertil. Dev. 2016, 28, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Sordillo, L.; Shafer-Weaver, K.; DeRosa, D. Immunobiology of the Mammary Gland. J. Dairy Sci. 1997, 80, 1851–1865. [Google Scholar] [CrossRef]
- Elenkov, I.J.; Chrousos, G.P. Stress Hormones, Th1/Th2 patterns, Pro/Anti-inflammatory Cytokines and Susceptibility to Disease. Trends Endocrinol. Metab. TEM 1999, 10, 359–368. [Google Scholar] [CrossRef]
- Ohtsuka, H.; Kudo, K.; Mori, K.; Nagai, F.; Hatsugaya, A.; Tajima, M.; Tamura, K.; Hoshi, F.; Koiwa, M.; Kawamura, S.-I. Acute Phase Response in Naturally Occurring Coliform Mastitis. J. Vet. Med. Sci. 2001, 63, 675–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunz-Ebrecht, S.R.; Mohamed-Ali, V.; Feldman, P.J.; Kirschbaum, C.; Steptoe, A. Cortisol responses to mild psychological stress are inversely associated with proinflammatory cytokines. Brain Behav. Immun. 2003, 17, 373–383. [Google Scholar] [CrossRef]
- Duncan, L.M.; Meegan, L.S.; Unanue, E.R. IL-1 gene expression in lymphoid tissues. J. Immunol. 1991, 146, 565–571. [Google Scholar] [PubMed]
- Riollet, C.; Rainard, P.; Poutrel, B. Kinetics of cells and cytokines during immune-mediated inflammation in the mammary gland of cows systemically immunized with Staphylococcus aureus α-toxin. Inflamm. Res. 2000, 49, 486–496. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouroutzika, E.; Ciliberti, M.G.; Caroprese, M.; Theodosiadou, E.; Papadopoulos, S.; Makri, S.; Skaperda, Z.-V.; Kotsadam, G.; Michailidis, M.-L.; Valiakos, G.; et al. Association of Melatonin Administration in Pregnant Ewes with Growth, Redox Status and Immunity of Their Offspring. Animals 2021, 11, 3161. https://doi.org/10.3390/ani11113161
Bouroutzika E, Ciliberti MG, Caroprese M, Theodosiadou E, Papadopoulos S, Makri S, Skaperda Z-V, Kotsadam G, Michailidis M-L, Valiakos G, et al. Association of Melatonin Administration in Pregnant Ewes with Growth, Redox Status and Immunity of Their Offspring. Animals. 2021; 11(11):3161. https://doi.org/10.3390/ani11113161
Chicago/Turabian StyleBouroutzika, Efterpi, Maria Giovanna Ciliberti, Mariangela Caroprese, Ekaterini Theodosiadou, Serafeim Papadopoulos, Sotiria Makri, Zoi-Vasiliki Skaperda, Georgios Kotsadam, Marios-Lazaros Michailidis, George Valiakos, and et al. 2021. "Association of Melatonin Administration in Pregnant Ewes with Growth, Redox Status and Immunity of Their Offspring" Animals 11, no. 11: 3161. https://doi.org/10.3390/ani11113161