
Discrimination of Acoustic Stimuli and Maintenance of Graded Alarm Call Structure in Captive Meerkats



Abstract
:Simple Summary
Abstract
1. Introduction
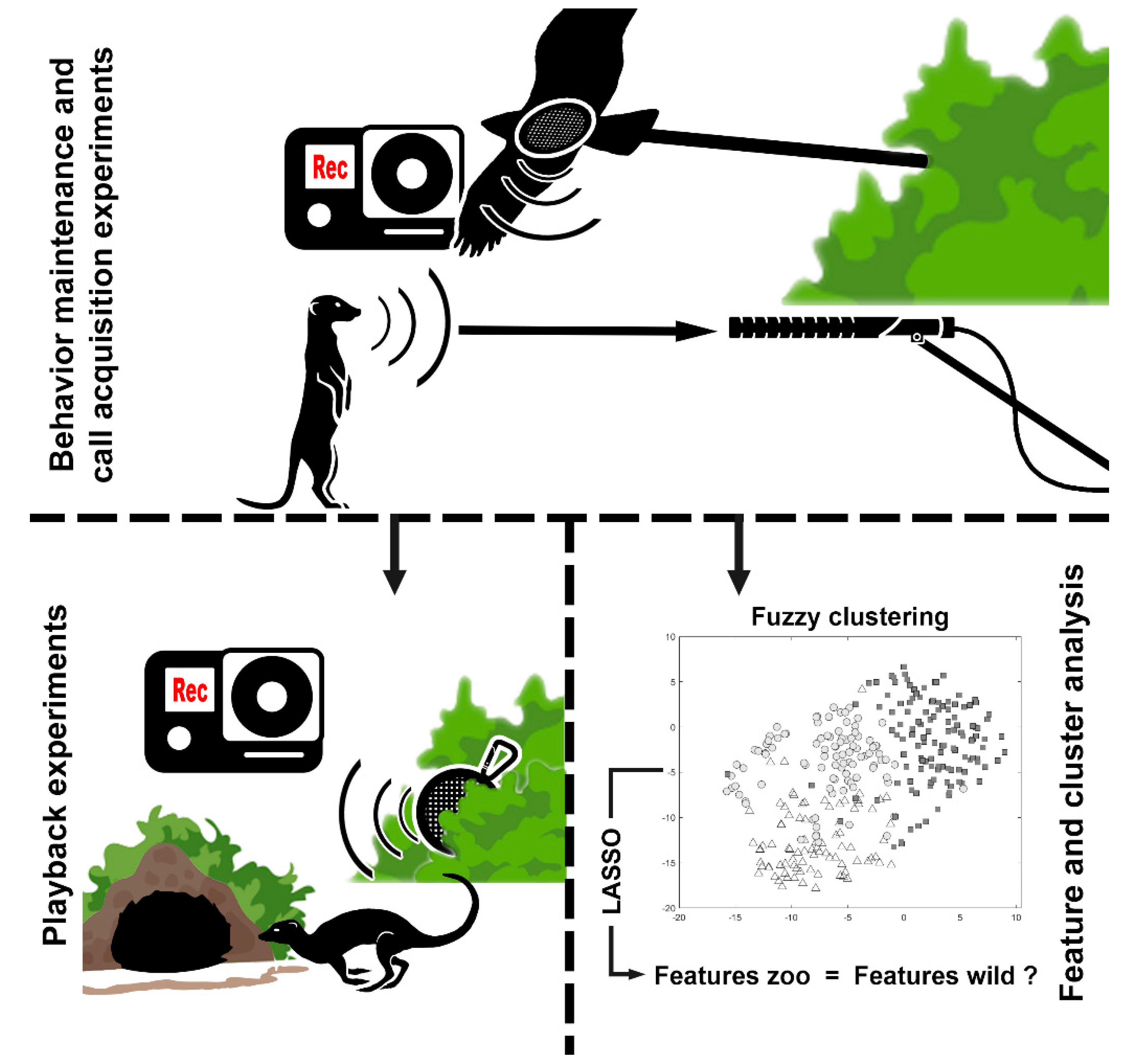
2. Materials and Methods
2.1. Study Site
2.2. Experimental Background
2.2.1. Behavior Maintenance and Call Acquisition Experiments
2.2.2. Playback Experiments
2.2.3. Feature and Cluster Analysis
2.3. Behavior Analysis
3. Results
3.1. Behavior Maintenance on Acoustic Stimuli
3.2. Playback Experiments
3.3. Feature and Cluster Analysis
4. Discussion
4.1. Meerkats Can Discriminate Acoustic Stimuli in Terms of Their Potential Significance
4.2. Feature and Cluster Analysis
4.3. Relevance for Animal Welfare and Conservation
4.4. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Binding, S.; Farmer, H.; Krusin, L.; Cronin, K. Status of animal welfare research in zoos and aquariums: Where are we, where to next? JZAR 2020, 8, 166–174. [Google Scholar] [CrossRef]
- Wolfensohn, S.; Shotton, J.; Bowley, H.; Davies, S.; Thompson, S.; Justice, W.S.M. Assessment of welfare in zoo animals: Towards optimum quality of life. Animals 2018, 8, 110. [Google Scholar] [CrossRef] [Green Version]
- Kleiman, D.G.; Thompson, K.V.; Baer, C.K. Wild Mammals in Captivity: Principles and Techniques for Zoo Management, 2nd ed.; University of Chicago Press: Chicago, IL, USA, 2010; ISBN 9780226440118. [Google Scholar]
- Carlstead, K.; Shepherdson, D. Effects of environmental enrichment on reproduction. Zoo Biol. 1994, 13, 447–458. [Google Scholar] [CrossRef]
- Altman, J.D. Animal activity and visitor learning at the zoo. Anthrozoos Multidiscip. J. Interact. People Anim. 1998, 11, 12–21. [Google Scholar] [CrossRef]
- Carr, N. An analysis of zoo visitors’ favourite and least favourite animals. Tour. Manag. Perspect. 2016, 20, 70–76. [Google Scholar] [CrossRef]
- Bradbury, J.W.; Vehrencamp, S.L. Principles of Animal Communication, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2011; ISBN 9780878930456. [Google Scholar]
- Gerhardt, H.C.; Huber, F. Acoustic Communication in Insects and Anurans: Common Problems and Diverse Solutions; University of Chicago Press: London, UK, 2002; ISBN 0226288331. [Google Scholar]
- Sun, G.-Z.; Huang, B.; Guan, Z.-H.; Geissmann, T.; Jiang, X.-L. Individuality in male songs of wild black crested gibbons (Nomascus concolor). Am. J. Primatol. 2011, 73, 431–438. [Google Scholar] [CrossRef]
- Clink, D.J.; Bernard, H.; Crofoot, M.C.; Marshall, A.J. Investigating individual vocal signatures and small-scale patterns of geographic variation in female bornean gibbon (Hylobates muelleri) great calls. Int. J. Primatol. 2017, 38, 656–671. [Google Scholar] [CrossRef]
- Jansen, D.A.W.A.M.; Cant, M.A.; Manser, M.B. Segmental concatenation of individual signatures and context cues in banded mongoose (Mungos mungo) close calls. BMC Biol. 2012, 10, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mumm, C.A.; Urrutia, M.C.; Knörnschild, M. Vocal individuality in cohesion calls of giant otters, Pteronura brasiliensis. Anim. Behav. 2014, 88, 243–252. [Google Scholar] [CrossRef]
- Schneider, S.; Dierkes, P.W. Localize Animal Sound Events Reliably (LASER): A new software for sound localization in zoos. JZBG 2021, 2, 146–163. [Google Scholar] [CrossRef]
- Janik, V.M.; Sayigh, L.S. Communication in bottlenose dolphins: 50 years of signature whistle research. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2013, 199, 479–489. [Google Scholar] [CrossRef]
- Kalan, A.K.; Boesch, C. Audience effects in chimpanzee food calls and their potential for recruiting others. Behav. Ecol. Sociobiol. 2015, 69, 1701–1712. [Google Scholar] [CrossRef]
- Manser, M.B. The acoustic structure of suricates’ alarm calls varies with predator type and the level of response urgency. Proc. Biol. Sci. 2001, 268, 2315–2324. [Google Scholar] [CrossRef] [Green Version]
- Blumstein, D.T. The evolution, function, and meaning of marmot alarm communication. In Advances in the Study of Behavior; Slater, P., Snowdon, C., Rosenblatt, J., Milinski, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 371–401. ISBN 9780120045372. [Google Scholar]
- Seyfarth, R.M.; Cheney, D.L.; Marler, P. Monkey responses to three different alarm calls: Evidence of predator classification and semantic communication. Science 1980, 210, 801–803. [Google Scholar] [CrossRef] [PubMed]
- Fichtel, C. Monkey Alarm Calling: It Ain’t all Referential, or is It? AB&C 2020, 7, 101–107. [Google Scholar] [CrossRef]
- Volodina, E.; Volodin, I.A. Bioacoustics in zoos: A review of applications and perspectives. Int. Zoo News 1999, 46, 208–213. [Google Scholar]
- Moberg, G.P. Biological response to stress: Key to assessment of animal well-being? In Animal Stress; Moberg, G.P., Ed.; Springer: New York, NY, USA, 1985; pp. 27–49. ISBN 978-1-4614-7544-6. [Google Scholar]
- Clinchy, M.; Sheriff, M.J.; Zanette, L.Y. Predator-induced stress and the ecology of fear. Funct. Ecol 2013, 27, 56–65. [Google Scholar] [CrossRef]
- Zanette, L.Y.; White, A.F.; Allen, M.C.; Clinchy, M. Perceived predation risk reduces the number of offspring songbirds produce per year. Science 2011, 334, 1398–1401. [Google Scholar] [CrossRef] [PubMed]
- Eggers, S.; Griesser, M.; Nystrand, M.; Ekman, J. Predation risk induces changes in nest-site selection and clutch size in the Siberian jay. Proc. Biol. Sci. 2006, 273, 701–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boinski, S.; Gross, T.S.; Davis, J.K. Terrestrial predator alarm vocalizations are a valid monitor of stress in captive brown capuchins (Cebus apella). Zoo Biol. 1999, 18, 295–312. [Google Scholar] [CrossRef]
- Kershenbaum, A.; Blumstein, D.T.; Roch, M.A.; Akçay, Ç.; Backus, G.; Bee, M.A.; Bohn, K.; Cao, Y.; Carter, G.; Cäsar, C.; et al. Acoustic sequences in non-human animals: A tutorial review and prospectus. Biol. Rev. Camb. Philos. Soc. 2016, 91, 13–52. [Google Scholar] [CrossRef] [Green Version]
- Wadewitz, P.; Hammerschmidt, K.; Battaglia, D.; Witt, A.; Wolf, F.; Fischer, J. Characterizing Vocal Repertoires--Hard vs. Soft Classification Approaches. PLoS ONE 2015, 10, e0125785. [Google Scholar] [CrossRef] [PubMed]
- Tibshirani, R. Regression shrinkage and selection via the lasso. J. R. Stat. Soc. Ser. B Stat. Methodol. 1996, 58, 267–288. [Google Scholar] [CrossRef]
- Hollén, L.I.; Clutton-Brock, T.; Manser, M.B. Ontogenetic changes in alarm-call production and usage in meerkats (Suricata suricatta): Adaptations or constraints? Behav. Ecol. Sociobiol. 2008, 62, 821–829. [Google Scholar] [CrossRef] [Green Version]
- Mateo, J.M. Early auditory experience and the ontogeny of alarm-call discrimination in Belding’s ground squirrels (Spermophilus beldingi). J. Comp. Psychol. 1996, 110, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishnan, U.; Coss, R.G. Age differences in the responses to adult and juvenile alarm calls by bonnet macaques (Macaca radiata). Ethology 2000, 106, 131–144. [Google Scholar] [CrossRef] [Green Version]
- Hanson, M.T.; Coss, R.G. Age Differences in the Response of California Ground Squirrels (Spermophilus beecheyi) to Conspecific Alarm Calls. Ethology 2001, 107, 259–275. [Google Scholar] [CrossRef]
- McCowan, B.; Franceschini, N.V.; Vicino, G.A. Age differences and developmental trends in alarm peep responses by squirrel monkeys (Saimiri sciureus). Am. J. Primatol. 2001, 53, 19–31. [Google Scholar] [CrossRef]
- Hollén, L.I.; Manser, M.B. Ontogeny of alarm call responses in meerkats, Suricata suricatta: The roles of age, sex and nearby conspecifics. Anim. Behav. 2006, 72, 1345–1353. [Google Scholar] [CrossRef]
- Manser, M.B.; Bell, M.B.; Fletcher, L.B. The information that receivers extract from alarm calls in suricates. Proc. Biol. Sci. 2001, 268, 2485–2491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myles, S.; Montrose, V.T. The effects of olfactory stimulation on the behaviour of captive meerkats (Suricata suricatta). JZAR 2015, 3, 37–42. [Google Scholar] [CrossRef]
- Hollén, L.I.; Manser, M.B. Persistence of alarm-call behaviour in the absence of predators: A comparison between wild and captive-born meerkats (suricata suricatta). Ethology 2007, 113, 1038–1047. [Google Scholar] [CrossRef] [Green Version]
- Friard, O.; Gamba, M. BORIS: A free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 2016, 7, 1325–1330. [Google Scholar] [CrossRef]
- Kroodsma, D.E.; Byers, B.E.; Goodale, E.; Johnson, S.; Liu, W.-C. Pseudoreplication in playback experiments, revisited a decade later. Anim. Behav. 2001, 61, 1029–1033. [Google Scholar] [CrossRef] [Green Version]
- Manser, M.B. Meerkats-Identifying cognitive mechanisms underlying meerkat coordination and communication: Experimental designs in their natural habitat. In Field and Laboratory Methods in Animal Cognition: A Comparative Guide; Bueno-Guerra, N., Amici, F., Eds.; Cambridge University Press: Cambridge, UK, 2018; ISBN 978-1-108-41394-7. [Google Scholar]
- de Faria, C.M.; de Souza Sá, F.; Costa, D.D.L.; Da Silva, M.M.; Da Silva, B.C.; Young, R.J.; de Azevedo, C.S. Captive-born collared peccary (Pecari tajacu, Tayassuidae) fails to discriminate between predator and non-predator models. Acta Ethol. 2018, 21, 175–184. [Google Scholar] [CrossRef]
- Blumstein, D.T.; Daniel, J.C. The loss of anti-predator behaviour following isolation on islands. Proc. Biol. Sci. 2005, 272, 1663–1668. [Google Scholar] [CrossRef] [Green Version]
- Stein, B.E.; London, N.; Wilkinson, L.K.; Price, D.D. Enhancement of perceived visual intensity by auditory stimuli: A psychophysical analysis. J. Cogn. Neurosci. 1996, 8, 497–506. [Google Scholar] [CrossRef]
- Thornton, A.; McAuliffe, K. Teaching in wild meerkats. Science 2006, 313, 227–229. [Google Scholar] [CrossRef] [Green Version]
- Deecke, V.B.; Janik, V.M. Automated categorization of bioacoustic signals: Avoiding perceptual pitfalls. J. Acoust. Soc. Am. 2006, 119, 645–653. [Google Scholar] [CrossRef] [Green Version]
- Townsend, S.W.; Charlton, B.D.; Manser, M.B. Acoustic cues to identity and predator context in meerkat barks. Anim. Behav. 2014, 94, 143–149. [Google Scholar] [CrossRef]
- August, P.V.; Anderson, J.G.T. Mammal sounds and motivation-structural rules: A test of the hypothesis. J. Mammal. 1987, 68, 1–9. [Google Scholar] [CrossRef]
- Ouattara, K.; Zuberbühler, K.; N’goran, E.K.; Gombert, J.-E.; Lemasson, A. The alarm call system of female Campbell’s monkeys. Anim. Behav. 2009, 78, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Morris, V.; Pitcher, B.J.; Chariton, A. A cause for alarm: Increasing translocation success of captive individuals through alarm communication. Front. Conserv. Sci. 2021, 2, 1–9. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Part of the Study | Stimulus | Alarm Call | Local to the Zoo | Iterations |
|---|---|---|---|---|
| Behavior maintenance and call acquisition experiments | Common buzzard * (Buteo buteo) | Aerial | Yes | 6/6 * |
| Red kite (Milvus milvus) | Aerial | Yes | 7 | |
| Robin (Erithacus rubecula) | Yes | 7 | ||
| Blue tit (Cyanistes caeruleus) | Yes | 9 | ||
| Cuckoo (Cuculus canorus) | Yes | 7 | ||
| Common raven (Corvus corax) | Aerial | Yes | 8 | |
| Jackal buzzard (Buteo rufofuscus) | Aerial | No | 6 | |
| Common jackal (Canis aureus) | Terrestrial | No | 9 | |
| domestic dog (Weimaraner) | Terrestrial | (Yes) | 3 | |
| Playback experiments | Meerkat alarm call (Suricata suricatta) | Aerial | 10 | |
| Robin (Erithacus rubecula) | Yes | 10 | ||
| Feature and cluster analysis | Common buzzard * (Buteo buteo) | Aerial and Terrestrial | 6 * |
| Behavior Category | Behavior |
|---|---|
| environment-directed behavior | guard |
| observe | |
| self- and intraspecies- directed behavior | foraging |
| rest | |
| eat | |
| others | |
| flight | flight |
| walk | walk |
| n.v. | not visible |
| Aerial Alarm Call | Terrestrial Alarm Call |
|---|---|
| Duration | Duration |
| F0 | F0 |
| F1 | ΔF0-F1 |
| Harmonic ratio | Harmonic ratio |
| Spectral flatness | DFA1 |
| DFA2 | |
| DFA1maloc | |
| DFA2maloc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schneider, S.; Goettlich, S.; Diercks, C.; Dierkes, P.W. Discrimination of Acoustic Stimuli and Maintenance of Graded Alarm Call Structure in Captive Meerkats. Animals 2021, 11, 3064. https://doi.org/10.3390/ani11113064
Schneider S, Goettlich S, Diercks C, Dierkes PW. Discrimination of Acoustic Stimuli and Maintenance of Graded Alarm Call Structure in Captive Meerkats. Animals. 2021; 11(11):3064. https://doi.org/10.3390/ani11113064
Chicago/Turabian StyleSchneider, Sebastian, Sarah Goettlich, Charlette Diercks, and Paul Wilhelm Dierkes. 2021. "Discrimination of Acoustic Stimuli and Maintenance of Graded Alarm Call Structure in Captive Meerkats" Animals 11, no. 11: 3064. https://doi.org/10.3390/ani11113064