
The Response of Layer Hen Productivity and Egg Quality to an Additional Limestone Source When Offered Diets Differing in Calcium Concentrations and the Inclusion of Phytase

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Design, Protocol, and Animal Management
2.3. Egg Quality Assessment
2.4. Chemical Analysis of Diets, Limestone Supplement, and Excreta
2.5. Statistical Analysis
3. Results
3.1. Dietary Composition
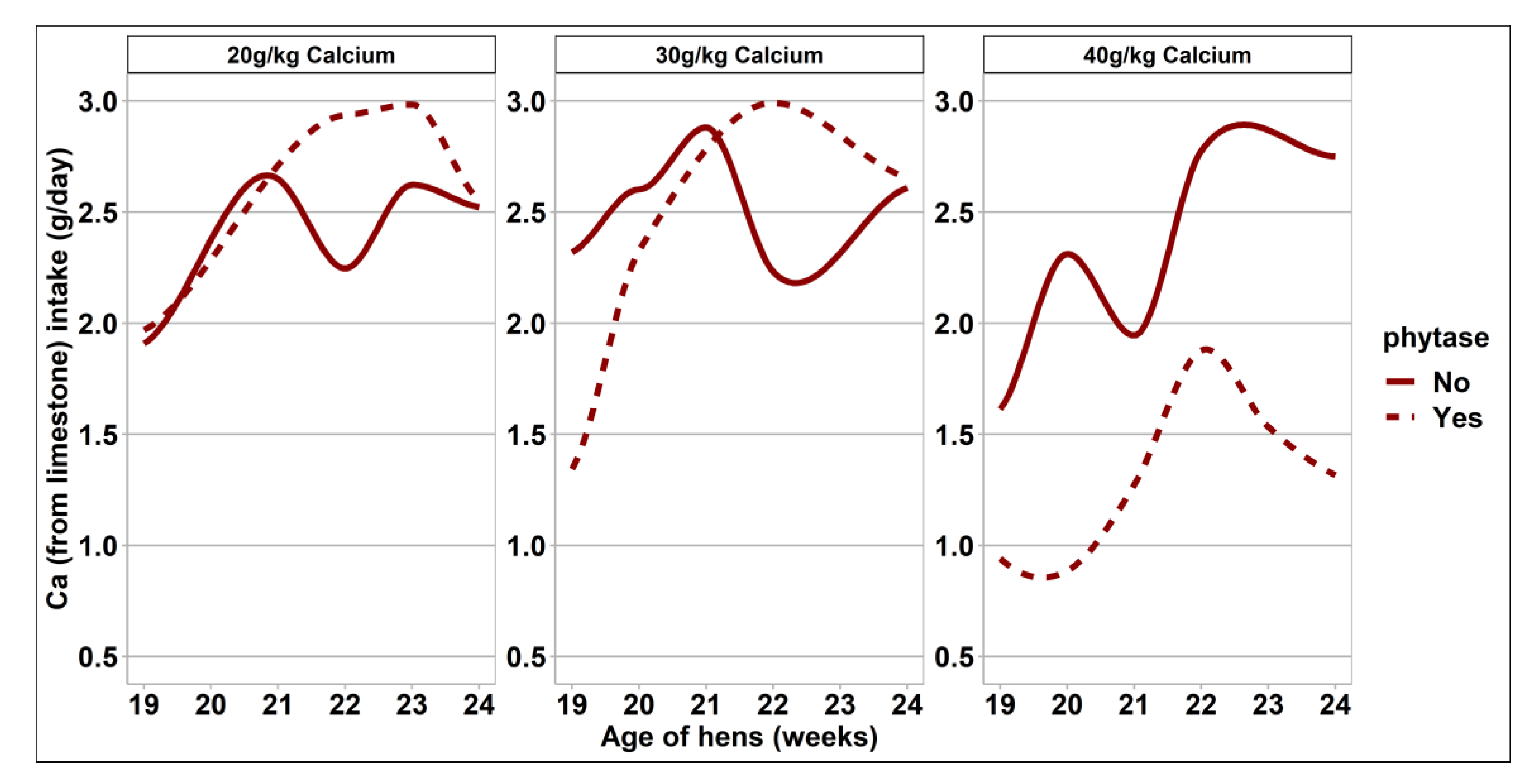
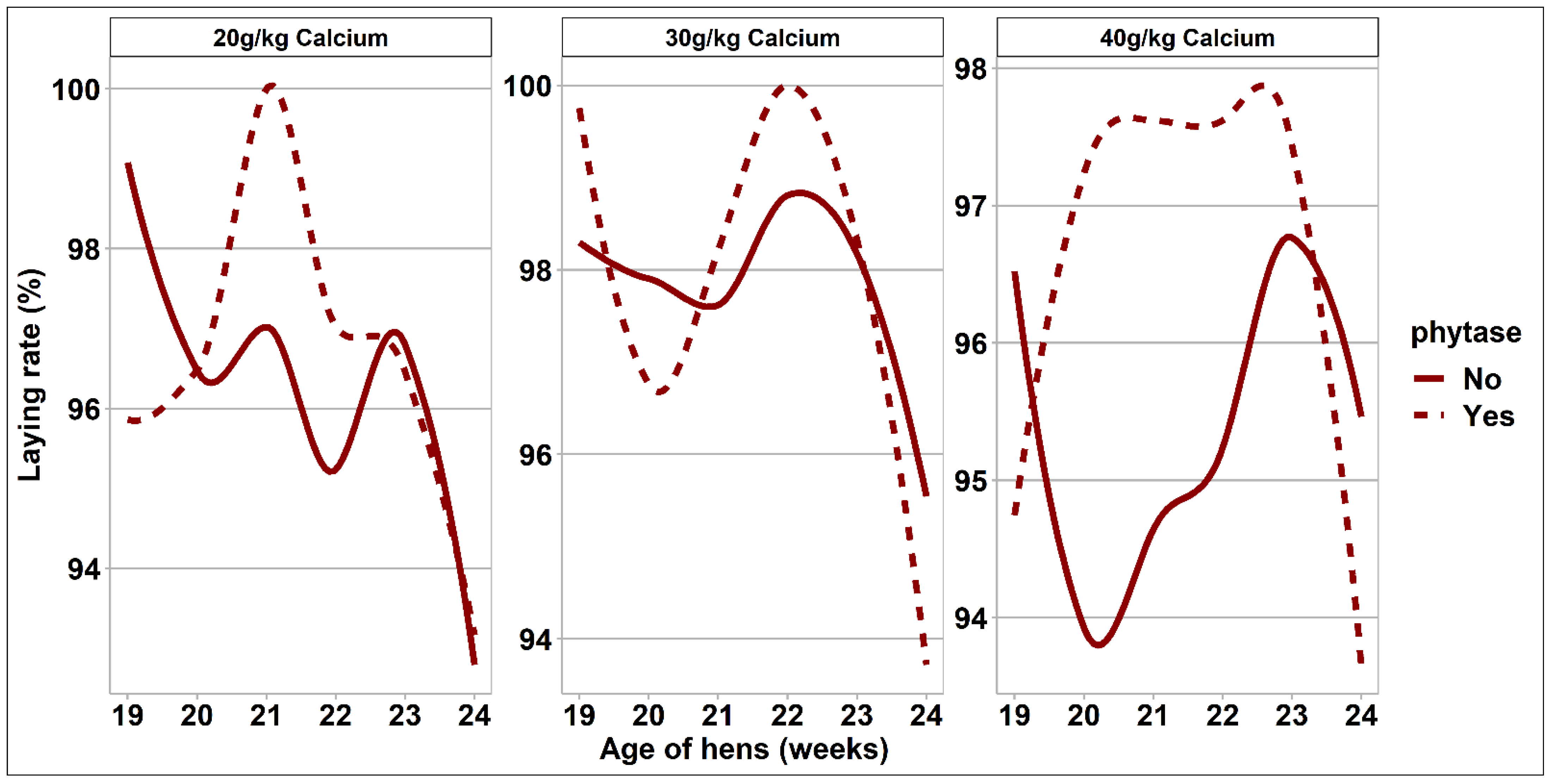
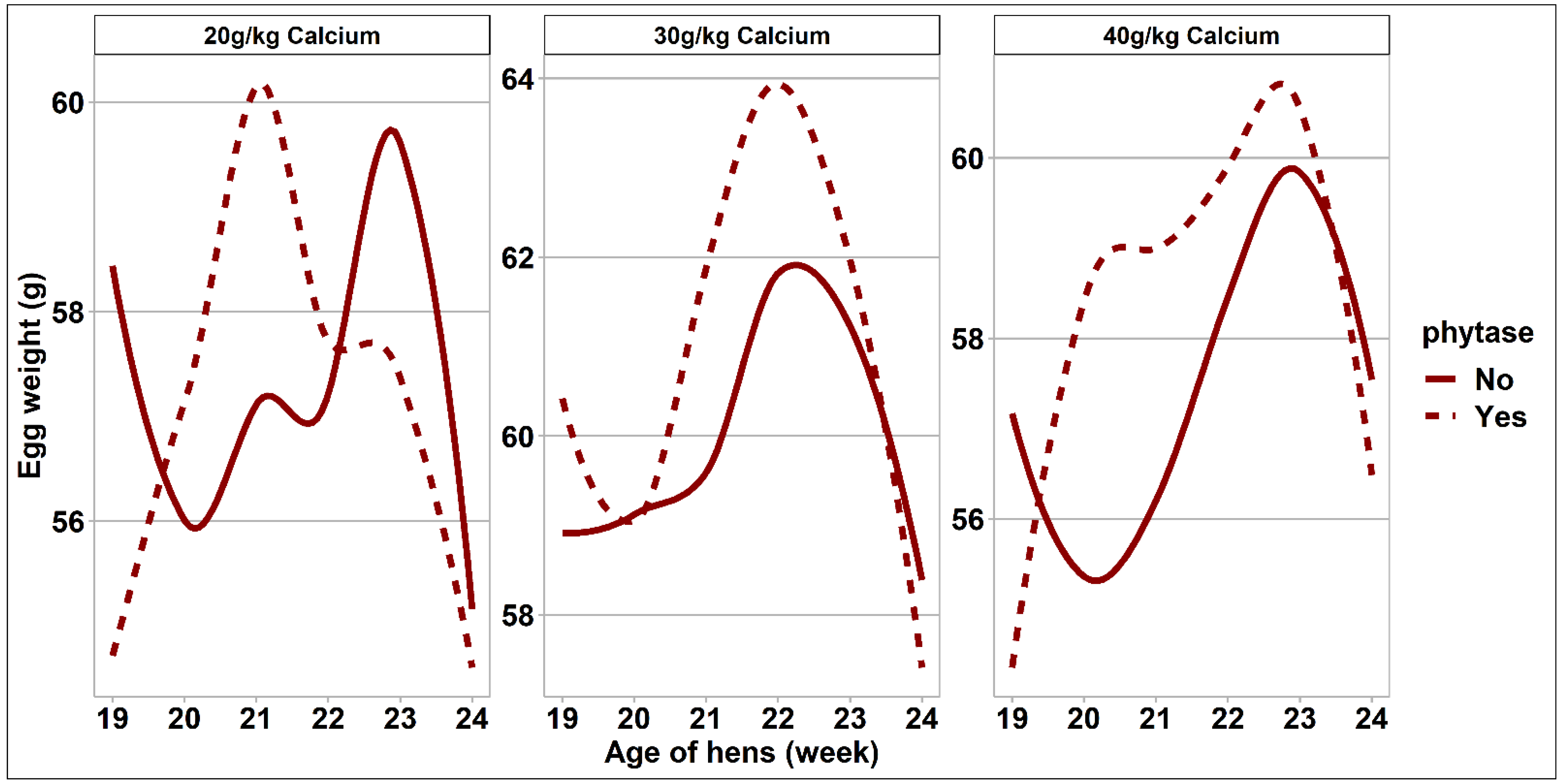
3.2. Bird Performance and Egg Quality Measurements
3.3. Nutrient Digestibility and Retention
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shafey, T.M.; McDonald, M.W.; Dingle, J.G. Effects of dietary calcium and available phosphorus concentration on digesta pH and on the availability of calcium, iron, magnesium and zinc from the intestinal contents of meat chickens. Brit. Poult. Sci. 1991, 32, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Morgan, N.K.; Walk, C.L.; Bedford, M.R.; Burton, E.J. The effect of dietary calcium inclusion on broiler gastrointestinal pH: Quantification and method optimization. Poult. Sci. 2014, 93, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Taylor, T.C. The availability of the calcium and phosphorus of plant materials for animals. Proc. Nutr. Soc. 1965, 24, 105–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selle, P.H.; Cowieson, A.J.; Ravindran, V. Consequences of calcium interactions with phytate and phytase for poultry and pigs. Livest. Sci. 2009, 124, 126–141. [Google Scholar] [CrossRef]
- Jiang, S.; Cui, L.; Shi, C.; Ke, X.; Luo, J.; Hou, J. Effects of dietary energy and calcium levels on performance, egg shell quality and bone metabolism in hens. Vet. J. 2013, 198, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Cufadar, Y.; Olgun, O.; Yildiz, A.Ö. The effect of dietary calcium concentration and particle size on performance, eggshell quality, bone mechanical properties and tibia mineral contents in moulted laying hens. Brit. Poult. Sci. 2011, 52, 761–768. [Google Scholar] [CrossRef]
- Holcombe, D.J.; Roland, D.A.; Harms, R.H. The ability of hens to adjust calcium intake when given a choice of diets containing two levels of calcium. Poult. Sci. 1975, 54, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Rose, S.P.; Kyriazakis, I. Diet selection of pigs and poultry. Proc. Nutr. Soc. 1991, 50, 87–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, S.J.; Selle, P.H.; Bedford, M.R.; Cowieson, A.J. Exploiting calcium-specific appetite in poultry nutrition. World Poult. Sci. J. 2011, 67, 587–598. [Google Scholar] [CrossRef]
- Joshua, I.G.; Mueller, W.J. The development of a specific appetite for calcium in growing broiler chicks. Brit. Poult. Sci. 1979, 20, 481–490. [Google Scholar] [CrossRef]
- Kyriazakis, I.; Tolkamp, B.J.; Emmans, G. Diet selection and animal state: An integrative framework. Proc. Nutr. Soc. 1999, 58, 765–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, S.J.; Selle, P.H.; Bedford, M.R.; Cowieson, A.J. Separate feeding of calcium improves performance and ileal nutrient digestibility in broiler chicks. Anim. Prod. Sci. 2014, 54, 172–178. [Google Scholar] [CrossRef]
- Snow, J.L.; Douglas, M.W.; Parsons, C.M. Phytase effects on amino acid digestibility in molted laying hens. Poult. Sci. 2003, 82, 474–477. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.S.; Namkung, H.; Paik, I.K. Effects of phytase supplementation on performance, egg quality and phosphorus excretion of laying hens fed different levels of dietary calcium and nonphytate phosphorus. Poult. Sci. 2003, 82, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Brennan, S.C.; Davies, T.S.; Schepelmann, M.; Riccardi, D. Emerging roles of the extracellular calcium-sensing receptor in nutrient sensing: Control of taste modulation and intestinal hormone secretion. Brit. J. Nutr. 2014, 111, S16–S22. [Google Scholar] [CrossRef] [Green Version]
- National Health and Medical Research Council (Australia); Australian Research Council. Australian Code for the Care and Use of Animals for Scientific Purposes; National Health and Medical Research Council: Australia, 2013. [Google Scholar]
- ISA Brown. Nutrition Management Guide; Institut de Selection Animale, BV: Boxmeer, The Netherlands, 2011. [Google Scholar]
- Şekeroǧlu, A.; Altuntaş, E. Effects of egg weight on egg quality characteristics. J. Sci. Food Agr. 2009, 89, 379–383. [Google Scholar] [CrossRef]
- Hill, F.W.; Anderson, D.L. Comparison of metabolizable energy and productive energy determinations with growing chicks. J. Nutr. 1958, 64, 587–603. [Google Scholar] [CrossRef]
- SAS University Edition; SAS Institute Inc.: Cary, NC, USA, 2015.
- Kassambara, A. Rstatix: Pipe-Friendly Framework for Basic Statistical Tests; R Package Version 0.7.0. 2020; Available online: https://CRAN.R-project.org/package=rstatix (accessed on 9 July 2021).
- Meunier-Salaün, M.C.; Faure, J.M. On the feeding and social behaviour of the laying hen. Appl. Anim. Behav. Sci. 1984, 13, 129–141. [Google Scholar] [CrossRef]
- Cransberg, P.H.; Parkinson, G.B.; Wilson, S.; Thorp, B.H. Sequential studies of skeletal calcium reserves and structural bone volume in a commercial layer flock. Brit. Poult. Sci. 2001, 42, 260–265. [Google Scholar] [CrossRef]
- Ziaei, N.; Shivazad, M.; Mirhadi, S.A.; Gerami, A. Effects of reduced calcium and phosphorous diets supplemented with phytase on laying performance of hens. Pak. J. Biol. Sci. 2009, 12, 792–797. [Google Scholar] [CrossRef] [Green Version]
- Leeson, S.; Summers, J.D. Influence of nutritional modification on skeletal size of leghorn and broiler breeder pullets. Poult. Sci. 1984, 63, 1222–1228. [Google Scholar] [CrossRef] [PubMed]
- Röhe, I.; Ruhnke, I.; Knorr, F.; Mader, A.; Boroojeni, F.G.; Löwe, R.; Zentek, J. Effects of grinding method, particle size, and physical form of the diet on gastrointestinal morphology and jejunal glucose transport in laying hens. Poult. Sci. 2014, 93, 2060–2068. [Google Scholar] [CrossRef] [PubMed]
- Ruhnke, I.; Röhe, I.; Krämer, C.; Boroojeni, F.G.; Knorr, F.; Mader, A.; Schulze, E.; Hafeez, A.; Neumann, K.; Löwe, R.; et al. The effects of particle size, milling method, and thermal treatment of feed on performance, apparent ileal digestibility, and pH of the digesta in laying hens. Poult. Sci. 2015, 94, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Pitargue, F.M.; Jung, H.; Han, G.P.; Choi, H.S.; Kil, D.Y. Effect of superdosing phytase on productive performance and egg quality in laying hens. Asian Austral. J. Anim. 2017, 30, 994–998. [Google Scholar] [CrossRef] [Green Version]
- Fernández, S.R.; Chárraga, S.; Ávila-Gonzalez, E. Evaluation of a new generation phytase on phytate phosphorus release for egg production and tibia strength in hens fed a corn-soybean meal diet. Poult. Sci. 2019, 98, 2087–2093. [Google Scholar] [CrossRef]
- Driver, J.P.; Pesti, G.M.; Bakalli, R.I.; Edwards, J.H.M. Effects of calcium and nonphytate phosphorus concentrations on phytase efficacy in broiler chicks. Poult. Sci. 2005, 84, 1406–1417. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Dietary Treatments | |||
|---|---|---|---|
| Calcium (g/kg Diet) | 40 | 30 | 20 |
| Ingredients (g/kg) | |||
| Wheat | 208 | 261 | 314 |
| Soybean meal (48% CP) | 266 | 255 | 244 |
| Soybean oil | 39 | 24 | 9.5 |
| Sorghum (9.2% CP) | 350 | 350 | 350 |
| Limestone | 101 | 74 | 47 |
| Dicalcium phosphorus | 6.9 | 6.5 | 6.1 |
| Salt | 2.4 | 2.3 | 2.3 |
| Sodium bicarbonate | 2.5 | 2.5 | 2.5 |
| L-Lysine HCl | 0.8 | 1.0 | 1.1 |
| DL-Methionine | 2.2 | 2.1 | 2.1 |
| L-Tryptophan | 0.3 | 0.3 | 0.3 |
| Threonine | 0.3 | 0.3 | 0.4 |
| Phytase 1 | (−/+) | (−/+) | (−/+) |
| Layer premix 2 | 1 | 1 | 1 |
| Celite | 20 | 20 | 20 |
| TOTAL | 1000 | 1000 | 1000 |
| Dietary Treatments | ||||
|---|---|---|---|---|
| Calcium (g/kg) | Units | 40 | 30 | 20 |
| Phytase (−/+) | (−/+) | (−/+) | (−/+) | |
| Calculated | ||||
| AME | MJ/kg | 11.5 | 11.5 | 11.5 |
| Crude protein (N × 6.25) | g/kg | 183 | 183 | 184 |
| Crude fibre | % | 2.1 | 2.1 | 2.2 |
| Calcium | g/kg | 40.0 | 30.0 | 20.0 |
| Phosphorus | g/kg | 4.8 | 4.8 | 4.8 |
| npP | g/kg | 2.5 | 2.5 | 2.5 |
| Calcium:npP | Ratio | 16.0 | 12.0 | 8.0 |
| Chloride | g/kg | 2.0 | 2.0 | 2.0 |
| Sodium | g/kg | 1.7 | 1.7 | 1.7 |
| TDC lysine | g/kg | 8.5 | 8.5 | 8.5 |
| TDC methionine | g/kg | 4.4 | 4.4 | 4.4 |
| TDC cysteine | g/kg | 2.6 | 2.6 | 2.7 |
| TDC total sulfur amino acids | g/kg | 7.0 | 7.0 | 7.1 |
| TDC threonine | g/kg | 6 | 6 | 6 |
| TDC tryptophan | g/kg | 1.9 | 1.9 | 1.9 |
| TDC valine | g/kg | 7.7 | 7.7 | 7.7 |
| Analyzed | ||||
| Phytase FYT/kg (−/+) | 106/3588 | 99/3436 | 137/4061 | |
| Ca (−/+ phytase) | g/kg | 46.0/50.8 | 33.4/33.2 | 25.8/32.0 |
| P | g/kg | 4.6 | 4.8 | 4.8 |
| Crude protein | g/kg | 19.9 | 19.6 | 20.3 |
| Gross Energy | MJ/kg | 15.6 | 15.8 | 15.7 |
| Dry Matter | g/kg | 91.0 | 90.7 | 90.4 |
| Calcium (g/kg) | Phytase | Initial Body Weight (g) | Final Body Weight (g) | Body Weight Change (%) | Feed Intake (g/day) | Ca Intake (from Feed, g/hen/day) | Ca Intake from Limestone (g/hen/day) | Total Ca Intake | Rate of Lay (%) | Egg Weight (g/hen/day) | Egg Mass (g/hen/day) | FCR |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 40 | − | 1890 | 1952 | 3.29 | 118 | 5.43 a | 2.45 | 7.88 | 98 | 62 | 61 | 1.947 |
| 40 | + | 1857 | 1930 | 3.93 | 117 | 5.94 b | 1.25 | 7.19 | 96 | 62 | 60 | 1.955 |
| 30 | − | 1857 | 1924 | 3.66 | 117 | 3.91 c | 2.54 | 6.45 | 98 | 62 | 61 | 1.918 |
| 30 | + | 1893 | 1936 | 2.27 | 120 | 3.99 c | 2.50 | 6.49 | 98 | 63 | 61 | 1.963 |
| 20 | − | 1854 | 1859 | 0.27 | 116 | 2.97 d | 2.81 | 5.78 | 98 | 61 | 60 | 1.940 |
| 20 | + | 1862 | 1881 | 1.02 | 116 | 3.72 c | 2.61 | 6.33 | 97 | 60 | 59 | 1.980 |
| SEM | 23 | 25 | 0.870 | 2.38 | 0.099 | 0.549 | 0.553 | 0.704 | 0.871 | 0.871 | 0.040 | |
| 40 | 1874 | 1941 a | 3.61 a | 117 | 5.68 | 1.85 | 7.53 a | 97 | 62 | 60 | 1.95 | |
| 30 | 1875 | 1930 a | 2.97 a | 119 | 3.95 | 2.52 | 6.47 ab | 98 | 63 | 61 | 1.94 | |
| 20 | 1858 | 1870 b | 0.64 b | 116 | 3.34 | 2.71 | 6.06 b | 98 | 61 | 59 | 1.96 | |
| SEM | 17 | 18 | 0.62 | 1.68 | 0.070 | 0.392 | 0.391 | 0.498 | 0.616 | 0.616 | 0.028 | |
| − | 1867 | 1911 | 2.40 | 117 | 4.10 | 2.60 | 6.70 | 98 | 62 | 61 | 1.93 | |
| + | 1871 | 1916 | 2.41 | 118 | 4.55 | 2.12 | 6.67 | 97 | 62 | 60 | 1.97 | |
| SEM | 13 | 14 | 0.503 | 1.37 | 0.057 | 0.320 | 0.320 | 0.407 | 0.503 | 0.503 | 0.02 | |
| p−values | ||||||||||||
| Ca | 0.725 | 0.014 | 0.004 | 0.599 | 0.001 | 0.196 | 0.031 | 0.590 | 0.128 | 0.105 | 0.886 | |
| Phytase | 0.849 | 0.842 | 0.994 | 0.666 | 0.001 | 0.024 | 0.938 | 0.345 | 0.784 | 0.457 | 0.344 | |
| Interaction | 0.349 | 0.657 | 0.393 | 0.635 | 0.007 | 0.275 | 0.532 | 0.603 | 0.706 | 0.670 | 0.886 |
| Variable | Low Limestone Grit Intake (n = 22) | High Limestone Grit Intake (n = 22) | SEM | p-Value |
|---|---|---|---|---|
| Limestone grit intake (g/day) * | 0.039 (0.013) | 6.97 (1.14) | - | - |
| Initial body weight (g) | 1841 | 1913 | 23.5 | 0.038 |
| Final body weight (g) | 1920 | 1911 | 18.9 | 0.744 |
| Feed intake (g/hen/day) | 114 | 114 | 1.86 | 0.855 |
| Ca intake (from feed, g/hen/day) | 4.57 | 3.91 | 0.226 | 0.043 |
| Rate of lay (%) | 91 | 97 | 1.52 | 0.024 |
| Egg weight (g/hen/day) | 57 | 61 | 1.34 | 0.067 |
| Egg mass (g/hen/day) | 53 | 59 | 1.87 | 0.030 |
| FCR | 2.01 | 1.94 | 0.036 | 0.182 |
| Calcium | Phytase | Egg Weight (g) | Albumen Height (mm) | Albumen Weight (g) | Haugh Unit | Yolk Height (mm) | Yolk Weight (g) | Yolk Color | Egg Breaking Force (g) | Shell Weight (g) | Shell Weight (%) | Shell Thickness (mm) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 40 | − | 63.5 | 9.8 | 37.9 | 98 | 16.9 | 15.4 | 11.8 | 4667 | 6.62 | 10.4 | 0.401 |
| 40 | + | 64.2 | 10.4 | 39.2 | 100 | 17.3 | 15.4 | 11.7 | 4781 | 6.78 | 10.5 | 0.406 |
| 20 | − | 62.4 | 9.7 | 37.6 | 97 | 17.1 | 14.9 | 11.2 | 4472 | 6.50 | 10.4 | 0.388 |
| 20 | + | 64.3 | 9.6 | 38.7 | 96 | 16.3 | 15.3 | 11.6 | 4448 | 6.32 | 9.9 | 0.384 |
| 30 | − | 60.9 | 9.8 | 37.3 | 98 | 17.1 | 14.9 | 10.9 | 4203 | 5.80 | 9.4 | 0.340 |
| 30 | + | 59.3 | 9.6 | 35.3 | 97 | 18.4 | 14.3 | 11.1 | 4246 | 5.74 | 9.7 | 0.337 |
| SEM | 1.13 | 0.40 | 0.97 | 1.79 | 0.629 | 0.269 | 0.180 | 170 | 0.17 | 0.26 | 0.01 | |
| 40 | 63.8 a | 10.11 | 38.6 | 98.9 | 17.1 | 15.4 a | 11.7 a | 4724 a | 6.70 a | 10.4 a | 0.403 a | |
| 30 | 63.3 a | 9.64 | 38.1 | 96.9 | 16.7 | 15.1 a | 11.4 a | 4460 ab | 6.41 a | 10.2 a | 0.386 b | |
| 20 | 60.1 b | 9.70 | 36.3 | 97.7 | 17.7 | 14.6 b | 11.0 b | 4224 b | 5.77 b | 9.6 b | 0.339 c | |
| SEM | 0.796 | 0.282 | 0.684 | 1.270 | 0.445 | 0.190 | 0.128 | 121 | 0.123 | 0.181 | 0.004 | |
| − | 62.3 | 9.79 | 37.6 | 97.7 | 17.1 | 15.1 | 11.3 | 4442 | 6.31 | 6.31 | 0.376 | |
| + | 62.6 | 9.85 | 37.7 | 98.0 | 17.3 | 15.0 | 11.5 | 4495 | 6.29 | 6.28 | 0.376 | |
| SEM | 0.650 | 0.231 | 0.559 | 1.03 | 0.34 | 0.13 | 0.10 | 97.9 | 0.09 | 0.100 | 0.003 | |
| p−values | ||||||||||||
| Ca | 0.003 | 0.449 | 0.053 | 0.555 | 0.260 | 0.010 | 0.002 | 0.010 | 0.001 | 0.006 | 0.001 | |
| Phytase | 0.735 | 0.861 | 0.883 | 0.842 | 0.568 | 0.784 | 0.279 | 0.705 | 0.874 | 0.866 | 0.892 | |
| Interaction | 0.314 | 0.496 | 0.168 | 0.570 | 0.265 | 0.165 | 0.476 | 0.917 | 0.614 | 0.220 | 0.737 |
| Ca (g/kg) | Phytase | AME | AMEn | N Retention | Dry Matter Retention |
|---|---|---|---|---|---|
| 40 | − | 13.5 | 13.4 | 0.535 | 0.648 |
| 40 | + | 13.5 | 13.3 | 0.527 | 0.739 |
| 30 | − | 13.7 | 13.6 | 0.501 | 0.731 |
| 30 | + | 13.7 | 13.6 | 0.530 | 0.695 |
| 20 | − | 13.6 | 13.5 | 0.577 | 0.704 |
| 20 | + | 13.5 | 13.4 | 0.587 | 0.697 |
| SEM | 0.304 | 0.296 | 0.038 | 0.029 | |
| 40 | 13.5 | 13.4 | 0.531 | 0.694 | |
| 30 | 13.7 | 13.6 | 0.515 | 0.713 | |
| 20 | 13.6 | 13.4 | 0.582 | 0.700 | |
| SEM | 0.215 | 0.210 | 0.027 | 0.020 | |
| − | 13.6 | 13.5 | 0.538 | 0.694 | |
| + | 13.6 | 13.4 | 0.548 | 0.710 | |
| SEM | 0.176 | 0.171 | 0.250 | 0.017 | |
| p−values | |||||
| Ca | 0.785 | 0.763 | 0.189 | 0.789 | |
| Phytase | 0.831 | 0.793 | 0.744 | 0.495 | |
| Interaction | 0.983 | 0.984 | 0.885 | 0.084 |
| Ca (g/kg) | Phytase | Na | Ca | P | Cu | Fe | Mg | K | Mn |
|---|---|---|---|---|---|---|---|---|---|
| 40 | − | 0.666 | 0.529 ac | 0.447 | 0.334 c | 0.350 | 0.209 | 0.304 ab | 0.143 |
| 40 | + | 0.710 | 0.740 abc | 0.363 | 0.239 bc | 0.517 | 0.399 | 0.435 c | 0.386 |
| 30 | − | 0.694 | 0.777 ab | 0.357 | 0.060 b | 0.434 | 0.402 | 0.308 ab | 0.182 |
| 30 | + | 0.664 | 0.551 c | 0.232 | 0.338 c | 0.364 | 0.338 | 0.279 b | 0.192 |
| 20 | − | 0.674 | 0.849 b | 0.406 | 0.378 ac | 0.537 | 0.349 | 0.405 ac | 0.313 |
| 20 | + | 0.689 | 0.821 b | 0.389 | 0.497 a | 0.486 | 0.406 | 0.363 abc | 0.319 |
| SEM | 0.024 | 0.085 | 0.057 | 0.053 | 0.072 | 0.079 | 0.037 | 0.055 | |
| 40 | 0.688 | 0.634 | 0.405 | 0.286 | 0.433 | 0.304 | 0.369 | 0.265 | |
| 30 | 0.679 | 0.664 | 0.294 | 0.199 | 0.399 | 0.370 | 0.294 | 0.187 | |
| 20 | 0.682 | 0.835 | 0.398 | 0.438 | 0.511 | 0.377 | 0.384 | 0.316 | |
| SEM | 0.017 | 0.060 | 0.041 | 0.038 | 0.051 | 0.056 | 0.026 | 0.040 | |
| − | 0.678 | 0.718 | 0.403 | 0.257 | 0.440 | 0.320 | 0.339 | 0.213 | |
| + | 0.688 | 0.704 | 0.328 | 0.358 | 0.456 | 0.381 | 0.359 | 0.299 | |
| SEM | 0.014 | 0.049 | 0.033 | 0.031 | 0.042 | 0.046 | 0.021 | 0.032 | |
| p−values | |||||||||
| Ca | 0.919 | 0.064 | 0.130 | 0.001 | 0.285 | 0.595 | 0.038 | 0.093 | |
| Phytase | 0.611 | 0.840 | 0.117 | 0.029 | 0.790 | 0.355 | 0.515 | 0.072 | |
| Interaction | 0.301 | 0.044 | 0.645 | 0.007 | 0.222 | 0.289 | 0.045 | 0.071 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruhnke, I.; Akter, Y.; Sibanda, T.Z.; Cowieson, A.J.; Wilkinson, S.; Maldonado, S.; Singh, M.; Hughes, P.; Caporale, D.; Bucker, S.; et al. The Response of Layer Hen Productivity and Egg Quality to an Additional Limestone Source When Offered Diets Differing in Calcium Concentrations and the Inclusion of Phytase. Animals 2021, 11, 2991. https://doi.org/10.3390/ani11102991
Ruhnke I, Akter Y, Sibanda TZ, Cowieson AJ, Wilkinson S, Maldonado S, Singh M, Hughes P, Caporale D, Bucker S, et al. The Response of Layer Hen Productivity and Egg Quality to an Additional Limestone Source When Offered Diets Differing in Calcium Concentrations and the Inclusion of Phytase. Animals. 2021; 11(10):2991. https://doi.org/10.3390/ani11102991
Chicago/Turabian StyleRuhnke, Isabelle, Yeasmin Akter, Terence Zimazile Sibanda, Aaron J. Cowieson, Stuart Wilkinson, Stephanie Maldonado, Mini Singh, Patrick Hughes, Dylana Caporale, Stephan Bucker, and et al. 2021. "The Response of Layer Hen Productivity and Egg Quality to an Additional Limestone Source When Offered Diets Differing in Calcium Concentrations and the Inclusion of Phytase" Animals 11, no. 10: 2991. https://doi.org/10.3390/ani11102991