
Natural Habitat Design for Zoo-Housed Elasmobranch and Teleost Fish Species Improves Behavioural Repertoire and Space Use in a Visitor Facing Exhibit


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Subjects and General Housing Conditions
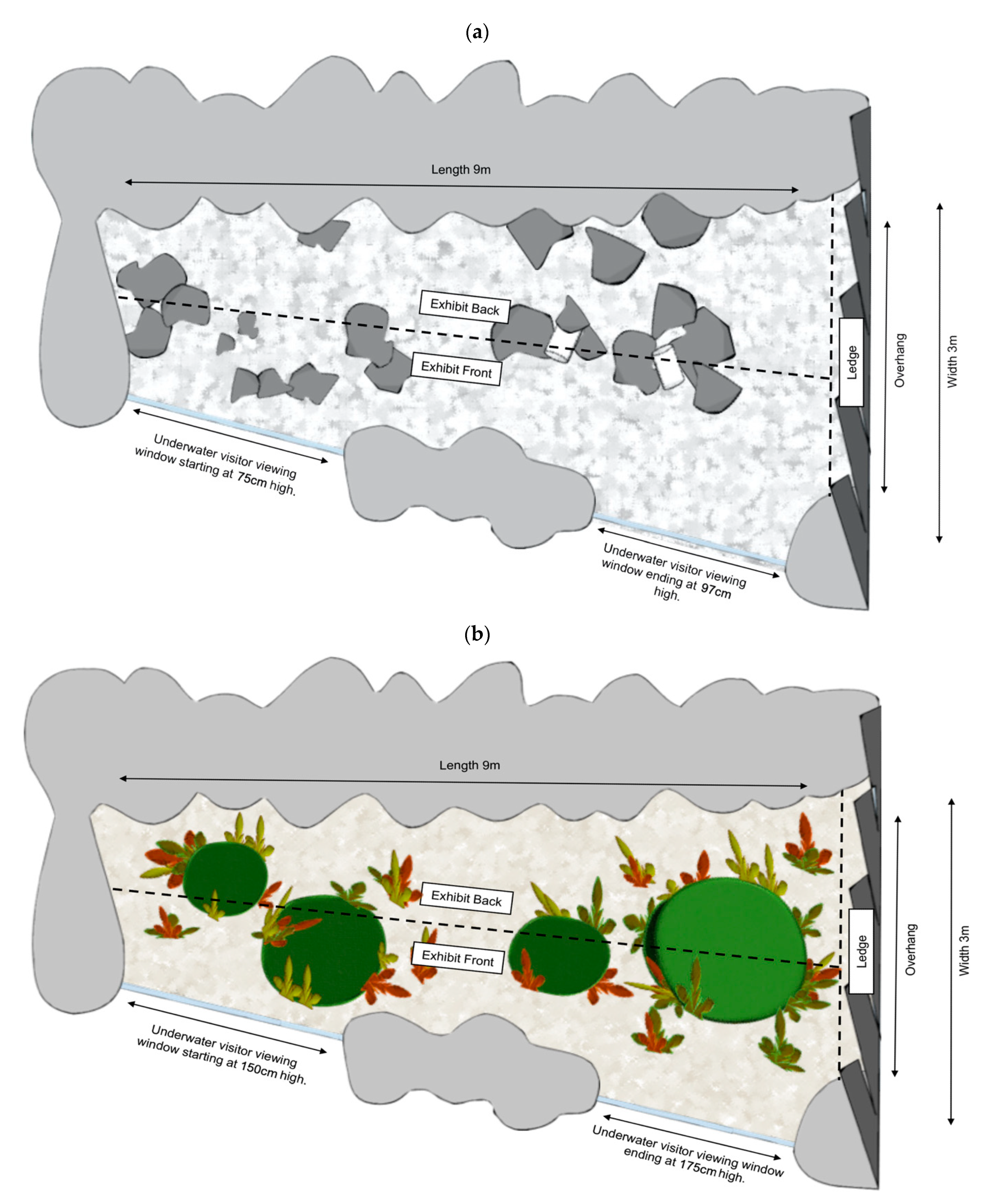
2.2. Tank Renovations (Treatments)
- Pre-renovation: tank had minimal environmental complexity with very little overhead cover, no seaweed theming, thick shell grit substrate and high visitor exposure through lack of barrier at water’s surface. The height of the glass on the visitor viewing side reached a height of 75 cm at the lowest point, rising to 97 cm at the highest (towards the ledge overhang) as the visitor ground sloped downward. The only structures within the tank included 40/60 cm × 40/60 cm size garden rocks, some clumped together, and two PVC pipes wedged in between two rock clumps providing cover for only small fish.
- Post-renovation: the tank was renovated to incorporate more naturalistic environmental features including the addition of four themed structures; a large swim through cave, two small enclosed caves with a front entrance and a large bommie structure providing overhead cover, finer coral sand substrate and multiple seaweed clusters. The glass wall was elevated to a height of 150 cm rising to 175 cm at the far side blocking direct visitor contact with water.
2.3. Behavioural Observations
2.4. Analysis
3. Results
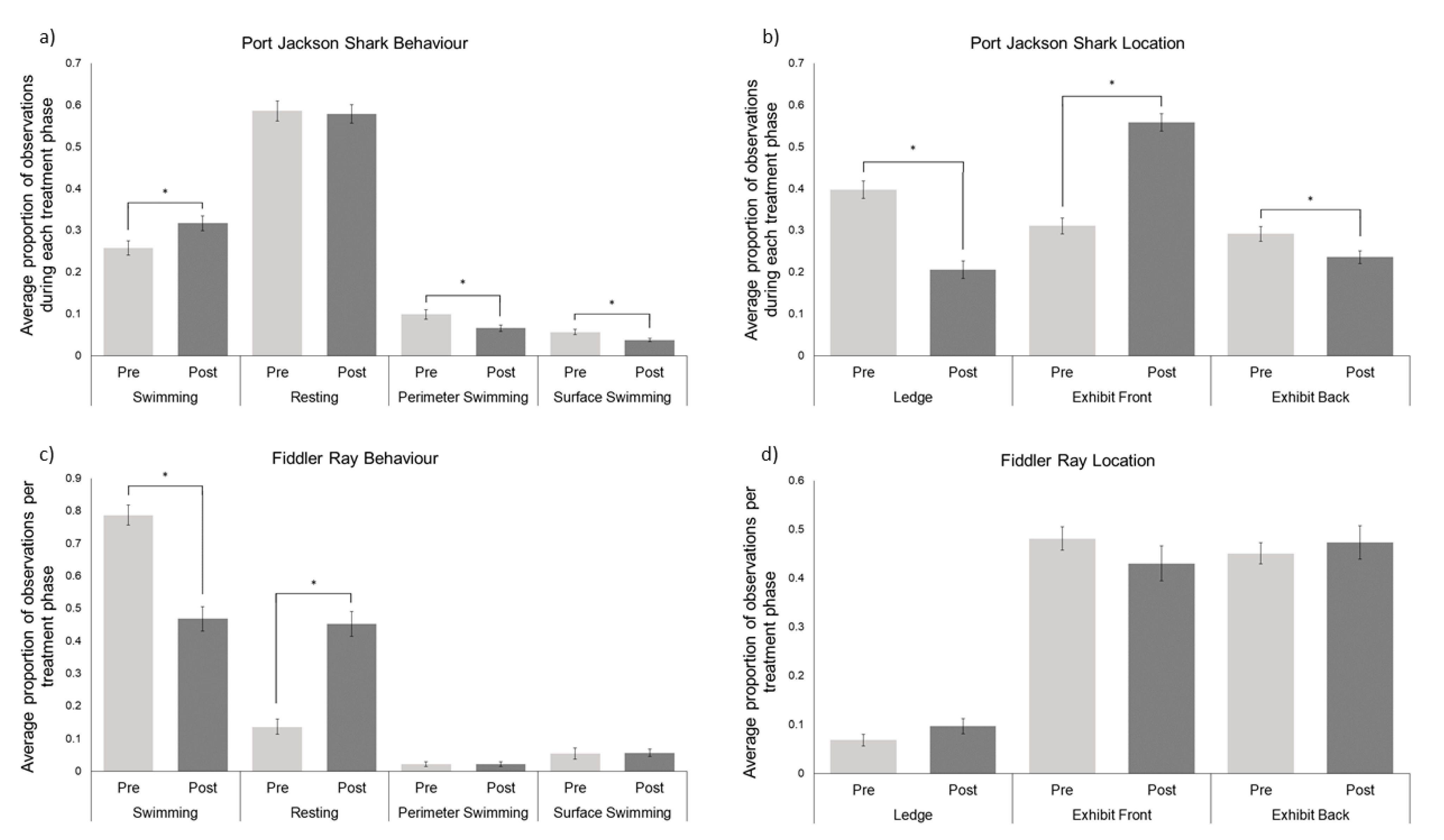
3.1. Elasmobranchs
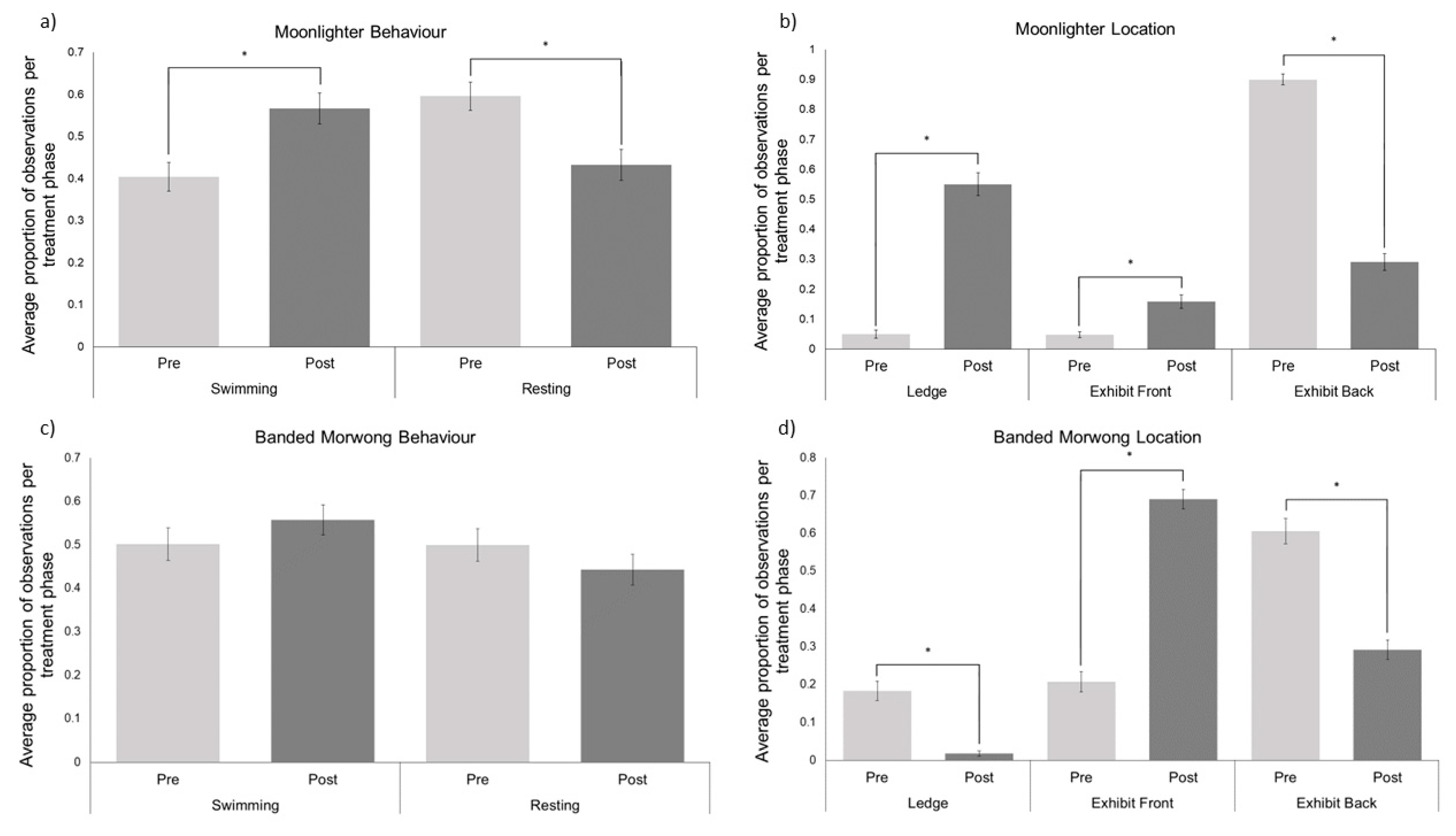
3.2. Teleost
4. Discussion
4.1. Port Jackson Sharks
4.2. Southern Fiddler Ray
4.3. Moonlighter and Banded Morwong
5. Limitations and Future Research
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kleiman, D.G.; Thompson, K.V.; Baer, C.K. Wild Mammals in Captivity: Principles and Techniques for Zoo Management; University of Chicago Press: Chicago, IL, USA, 2010. [Google Scholar]
- Melfi, V. There are big gaps in our knowledge, and thus approach, to zoo animal welfare: A case for evidence-based zoo animal management. Zoo Biol. Publ. Affil. Am. Zoo Aquar. Assoc. 2009, 28, 574–588. [Google Scholar] [CrossRef]
- Tribe, A.; Booth, R. Assessing the role of zoos in wildlife conservation. Hum. Dimens. Wildl. 2003, 8, 65–74. [Google Scholar] [CrossRef]
- Fraser, D. Understanding Animal Welfare: The Science in its Cultural Context; Wiley-Blackwell: Chichester, UK, 2008. [Google Scholar]
- Ward, S.J.; Sherwen, S.; Clark, F.E. Advances in applied zoo animal welfare science. J. Appl. Anim. Welf. Sci. 2018, 21, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broom, D.M. A history of animal welfare science. Acta Biotheor. 2011, 59, 121–137. [Google Scholar] [CrossRef] [PubMed]
- Fàbregas, M.C.; Guillén-Salazar, F.; Garcés-Narro, C. Do naturalistic enclosures provide suitable environments for zoo animals? Zoo Biol. 2012, 31, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Fife-Cook, I.; Franks, B. Positive welfare for fishes: Rationale and areas for future study. Fishes 2019, 4, 31. [Google Scholar] [CrossRef] [Green Version]
- Mason, G.J. Species differences in responses to captivity: Stress, welfare and the comparative method. Trends Ecol. Evol. 2010, 25, 713–721. [Google Scholar] [CrossRef] [Green Version]
- Alligood, C.; Leighty, K. Putting the “E” in SPIDER: Evolving trends in the evaluation of environmental enrichment efficacy in zoological settings. Anim. Behav. Cogn. 2015, 2, 200–217. [Google Scholar] [CrossRef]
- Arechavala-Lopez, P.; Diaz-Gil, C.; Saraiva, J.L.; Moranta, D.; Castanheira, M.F.; Nuñez-Velázquez, S.; Ledesma-Corvi, S.; Mora-Ruiz, M.; Grau, A. Effects of structural environmental enrichment on welfare of juvenile seabream (Sparus aurata). Aquac. Rep. 2019, 15, 100224. [Google Scholar] [CrossRef]
- Fanson, K.; Wielebnowski, N. Effect of housing and husbandry practices on adrenocortical activity in captive Canada lynx (Lynx canadensis). Anim. Welf. 2013, 22, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Mallapur, A.; Chellam, R. Environmental influences on stereotypy and the activity budget of Indian leopards (Panthera pardus) in four zoos in Southern India. Zoo Biol. Publ. Affil. Am. Zoo Aquar. Assoc. 2002, 21, 585–595. [Google Scholar] [CrossRef]
- Mellor, D. Positive animal welfare states and encouraging environment-focused and animal-to-animal interactive behaviours. N. Z. Vet. J. 2015, 63, 9–16. [Google Scholar] [CrossRef]
- Sherwen, S.L.; Hemsworth, P.H. The visitor effect on zoo animals: Implications and opportunities for zoo animal welfare. Animals 2019, 9, 366. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Bai, Q.; Xu, X.; Guo, H.; Zhang, X. Effects of environmental enrichment on the welfare of juvenile black rockfish Sebastes schlegelii: Growth, behavior and physiology. Aquaculture 2020, 518, 734782. [Google Scholar] [CrossRef]
- Fraser, D. Assessing animal welfare: Different philosophies, different scientific approaches. Zoo Biol. Publ. Affil. Am. Zoo Aquar. Assoc. 2009, 28, 507–518. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, A.B.; Bashaw, M.J.; Maple, T.L. Scientific Foundations of Zoos and Aquariums: Their Role in Conservation and Research; Cambridge University Press: Cambridge, UK, 2019. [Google Scholar]
- Špinka, M.; Wemelsfelder, F. Environmental challenge and animal agency. Anim. Welf. 2011, 2, 27–44. [Google Scholar]
- Shepherdson, D.J. Environmental enrichment: Past, present and future. Int. Zoo Yearb. 2003, 38, 118–124. [Google Scholar] [CrossRef]
- Young, R.J. Environmental Enrichment for Captive Animals; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Hutchins, M.; Smith, B. Characteristics of a world-class zoo or aquarium in the 21st century. Int. Zoo Yearb. 2003, 38, 130–141. [Google Scholar] [CrossRef]
- Barber, J.C. Programmatic approaches to assessing and improving animal welfare in zoos and aquariums. Zoo Biol. Publ. Affil. Am. Zoo Aquar. Assoc. 2009, 28, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Maple, T.L. Toward a science of welfare for animals in the zoo. J. Appl. Anim. Welf. Sci. 2007, 10, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.J.; Vicino, G.A.; Sheftel, J.; Lauderdale, L.K. Behavioral diversity as a potential indicator of positive animal welfare. Animals 2020, 10, 1211. [Google Scholar] [CrossRef]
- Chamove, A.S.; Anderson, J.R.; Morgan-Jones, S.C.; Jones, S.P. Deep woodchip litter: Hygiene, feeding, and behavioral enhancement in eight primate species. Int. J. Study Anim. Probl. 1982, 3, 308–318. [Google Scholar]
- Rose, P.; Robert, R. Evaluating the activity patterns and enclosure usage of a little-studied zoo species, the sitatunga (Tragelaphus spekii). J. Zoo Aquar. Res. 2013, 1, 14–19. [Google Scholar]
- Rose, P.E.; Nash, S.M.; Riley, L.M. To pace or not to pace? A review of what abnormal repetitive behavior tells us about zoo animal management. J. Vet. Behav. 2017, 20, 11–21. [Google Scholar] [CrossRef]
- Mellor, D.J.; Hunt, S.; Gusset, M. Caring for wildlife: The world zoo and aquarium animal welfare strategy. WAZA Exec. Off. 2015, 1, 1–87. [Google Scholar]
- Swaisgood, R.R.; Shepherdson, D.J. Scientific approaches to enrichment and stereotypies in zoo animals: What’s been done and where should we go next? Zoo Biol. Publ. Affil. Am. Zoo Aquar. Assoc. 2005, 24, 499–518. [Google Scholar] [CrossRef]
- Balcombe, J. What a Fish Knows: The Inner Lives of Our Underwater Cousins; Scientific American/Farrar, Straus and Giroux: New York, NY, USA, 2016. [Google Scholar]
- Mather, J.A. Ethics and care: For animals, not just mammals. Animals 2019, 9, 1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, P.E.; Brereton, J.E.; Rowden, L.J.; de Figueiredo, R.L.; Riley, L.M. What’s new from the zoo? An analysis of ten years of zoo-themed research output. Palgrave Commun. 2019, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Braithwaite, V.A.; Huntingford, F.A. Fish and welfare: Do fish have the capacity for pain perception and suffering? Animal Welf. 2004, 13, S87–S92. [Google Scholar]
- Brown, C. Fish intelligence, sentience and ethics. Anim. Cogn. 2015, 18, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.; Laland, K.; Krause, J. Fish Cognition and Behavior; John Wiley & Sons: Hoboken, NJ, USA, 2011; Volume 21. [Google Scholar]
- Walker, M.; Diez-Leon, M.; Mason, G. Animal welfare science: Recent publication trends and future research priorities. Int. J. Consum. Stud. 2014, 27, 80–100. [Google Scholar]
- Sneddon, L.U.; Braithwaite, V.A.; Gentle, M.J. Do fishes have nociceptors? Evidence for the evolution of a vertebrate sensory system. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 1115–1121. [Google Scholar] [CrossRef] [Green Version]
- Braithwaite, V.A.; Huntingford, F.; van den Bos, R. Variation in emotion and cognition among fishes. J. Agric. Environ. Ethics 2013, 26, 7–23. [Google Scholar] [CrossRef]
- Cerqueira, M.; Millot, S.; Castanheira, M.F.; Félix, A.S.; Silva, T.; Oliveira, G.; Oliveira, C.; Martins, C.; Oliveira, R.F. Cognitive appraisal of environmental stimuli induces emotion-like states in fish. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Brown, C. Familiarity with the test environment improves escape responses in the crimson spotted rainbowfish, Melanotaenia duboulayi. Anim. Cogn. 2001, 4, 109–113. [Google Scholar] [CrossRef]
- Odling-Smee, L.; Braithwaite, V.A. The role of learning in fish orientation. Fish Fish. 2003, 4, 235–246. [Google Scholar] [CrossRef]
- da Silva, A.; Lima, M.R.; Meletti, P.C.; Jerep, F.C. Impact of environmental enrichment and social group size in the aggressiveness and foraging activity of Serrapinnus notomelas. Appl. Anim. Behav. Sci. 2020, 224, 104943. [Google Scholar] [CrossRef]
- Grosenick, L.; Clement, T.S.; Fernald, R.D. Fish can infer social rank by observation alone. Nature 2007, 445, 429–432. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.I.; Galhardo, L.; Noble, C.; Damsgård, B.; Spedicato, M.T.; Zupa, W.; Beauchaud, M.; Kulczykowska, E.; Massabuau, J.-C.; Carter, T. Behavioural indicators of welfare in farmed fish. Fish Physiol. Biochem. 2012, 38, 17–41. [Google Scholar] [CrossRef] [Green Version]
- Sadoul, B.; Geffroy, B. Measuring cortisol, the major stress hormone in fishes. J. Fish Biol. 2019, 94, 540–555. [Google Scholar] [CrossRef] [Green Version]
- van de Nieuwegiessen, P.G.; Boerlage, A.S.; Verreth, J.A.; Schrama, J.W. Assessing the effects of a chronic stressor, stocking density, on welfare indicators of juvenile African catfish, Clarias gariepinus Burchell. Appl. Anim. Behav. Sci. 2008, 115, 233–243. [Google Scholar] [CrossRef]
- Näslund, J.; Johnsson, J.I. Environmental enrichment for fish in captive environments: Effects of physical structures and substrates. Fish Fish. 2016, 17, 1–30. [Google Scholar] [CrossRef]
- Bshary, R.; Wickler, W.; Fricke, H. Fish cognition: A primate’s eye view. Anim. Cogn. 2002, 5, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.J.; Paull, G.C.; Tyler, C.R. Effects of environmental enrichment on survivorship, growth, sex ratio and behaviour in laboratory maintained zebrafish Danio rerio. J. Fish Biol. 2019, 94, 86–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woodward, M.A.; Winder, L.A.; Watt, P.J. Enrichment increases aggression in zebrafish. Fishes 2019, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Delicio, H.C.; Barreto, R.E.; Normandes, E.B.; Luchiari, A.C.; Marcondes, A.L. A place preference test in the fish Nile tilapia. J. Exp. Anim. Sci. 2006, 43, 141–148. [Google Scholar] [CrossRef]
- Jones, N.A.; Spence, R.; Jones, F.A.; Spence-Jones, H.C. Shade as enrichment: Testing preferences for shelter in two model fish species. J. Fish Biol. 2019, 95, 1161–1165. [Google Scholar] [CrossRef]
- Sullivan, M.; Lawrence, C.; Blache, D. Why did the fish cross the tank? Objectively measuring the value of enrichment for captive fish. Appl. Anim. Behav. Sci. 2016, 174, 181–188. [Google Scholar] [CrossRef]
- Neto, J.F.; Giaquinto, P.C. Environmental enrichment techniques and tryptophan supplementation used to improve the quality of life and animal welfare of Nile tilapia. Aquac. Rep. 2020, 17, 100354. [Google Scholar] [CrossRef]
- Blanchett, M. Investigating the Use of GIS for Monitoring Visitor Effects and Enclosure Use of Captive Birds in a Mixed Species, Free-Ranging Aviary at Lowry Park Zoo, Tampa FL. Master’s Thesis, University of Guelph, Guelph, ON, Canada, 2018. [Google Scholar]
- Dancer, A.M.; Burn, C.C. Visitor effects on zoo-housed Sulawesi crested macaque (Macaca nigra) behaviour: Can signs with ‘watching eyes’ requesting quietness help? Appl. Anim. Behav. Sci. 2019, 211, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Kathleen, M.; Tomborg, C. Sources of stress in captivity. Appl. Anim. Behav. Sci. 2007, 102, 262–302. [Google Scholar]
- Larsen, M.J.; Sherwen, S.L.; Rault, J.-L. Number of nearby visitors and noise level affect vigilance in captive koalas. Appl. Anim. Behav. Sci. 2014, 154, 76–82. [Google Scholar] [CrossRef]
- Sherwen, S.L.; Magrath, M.J.; Butler, K.L.; Hemsworth, P.H. Little penguins, Eudyptula minor, show increased avoidance, aggression and vigilance in response to zoo visitors. Appl. Anim. Behav. Sci. 2015, 168, 71–76. [Google Scholar] [CrossRef]
- Smith, K.N.; Kuhar, C.W. Siamangs (Hylobates syndactylus) and white-cheeked gibbons (Hylobates leucogenys) show few behavioral differences related to zoo attendance. J. Appl. Anim. Welf. Sci. 2010, 13, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Suárez, P.; Recuerda, P.; Arias-de-Reyna, L. Behaviour and welfare: The visitor effect in captive felids. Anim. Welf. 2017, 26, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Wells, D.L. A note on the influence of visitors on the behaviour and welfare of zoo-housed gorillas. Appl. Anim. Behav. Sci. 2005, 93, 13–17. [Google Scholar] [CrossRef]
- Chamove, A.S.; Hosey, G.R.; Schaetzel, P. Visitors excite primates in zoos. Zoo Biol. 1988, 7, 359–369. [Google Scholar] [CrossRef]
- Glatston, A.R.; Geilvoet-Soeteman, E.; Hora-Pecek, E.; Van Hooff, J. The influence of the zoo environment on social behavior of groups of cotton-topped tamarins, Saguinus oedipus oedipus. Zoo Biol. 1984, 3, 241–253. [Google Scholar] [CrossRef]
- Mansour, A.A.; Zakaria, A.-H.; Fraser, A.F. Effect of enclosure quality on reactivity and welfare of captive Soemmerring’s gazelle (Gazella soemmerringii). J. Appl. Anim. Welf. Sci. 2000, 3, 335–343. [Google Scholar] [CrossRef]
- Carrier, J.C.; Musick, J.A.; Heithaus, M.R. Biology of Sharks and Their Relatives; CRC press: Boca Raton, FL, USA, 2012. [Google Scholar]
- Hart, N.S.; Lisney, T.J.; Collin, S.P. Visual communication in elasmobranchs. Commun. Fishes 2006, 2, 337–392. [Google Scholar]
- McLaughlin, R.H. The Ecology of Heterodont Sharks; University of New South Wales: Sydney, Australia, 1969. [Google Scholar]
- Powter, D.M.; Gladstone, W. Habitat-Mediated Use of Space by Juvenile and Mating Adult Port Jackson Sharks, Heterodontus portusjacksoni, in Eastern Australia1. Pac. Sci. 2009, 63, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kadar, J.; Ladds, M.; Mourier, J.; Day, J.; Brown, C. Acoustic accelerometry reveals diel activity patterns in premigratory Port Jackson sharks. Ecol. Evol. 2019, 9, 8933–8944. [Google Scholar] [CrossRef] [Green Version]
- Martins, C.L. Stress, Survival and Movement Following Fishing Gear Capture in Chondrichthyan Species. Ph.D. Thesis, Monash University, Clayton, Australia, 2017. [Google Scholar]
- Glennon, V.; Chisholm, L.; Whittington, I. Three unrelated species, 3 sites, same host monogenean parasites of the southern fiddler ray, Trygonorrhina fasciata, in South Australia: Egg hatching strategies and larval behaviour. Parasitology 2006, 133, 55–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izzo, C.; Gillanders, B.M. Initial assessment of age, growth and reproductive parameters of the southern fiddler ray Trygonorrhina fasciata (Müller & Henle, 1841) from South Australia. Pan-Am. J. Aquat. Sci. 2008, 3, 321–327. [Google Scholar]
- Shepherd, S.; Teale, J.; Muirhead, D. Cleaning symbiosis among inshore fishes at Althorpe Island, South Australia and elsewhere. Trans. R. Soc. S. Aust. 2005, 129, 193–201. [Google Scholar]
- Leum, L.; Choat, J. Density and distribution patterns of the temperate marine fish Cheilodactylus spectabilis (Cheilodactylidae) in a reef environment. Mar. Biol. 1980, 57, 327–337. [Google Scholar] [CrossRef]
- Shepherd, S.A.; Baker, J.L.; Brown, A. Reef fishes of NE Kangaroo Island: A nursery role for Eastern Cove? Report to Wildlife Conservation Fund; Department for Environment and Heritage, and Natural Resources Management Board: Kangaroo Island, Australia, 2008. [Google Scholar]
- Casamitjana, J. Aquatic Zoos. A Critical Study of UK Public Aquaria in the Year 2004; The Captive Animals’ Protection Society: Manchester, UK, 2004. [Google Scholar]
- Biasetti, P.; Florio, D.; Gili, C.; de Mori, B. The Ethical Assessment of Touch Pools in Aquariums by Means of the Ethical Matrix. J. Agric. Environ. Ethics 2020, 33, 337–353. [Google Scholar] [CrossRef]
- Grossman, R. Stereotypical surface breaking behaviour in captive rays (Genus: Raja) at the London aquarium. In Proceedings of the 7th Annual Symposium on Zoo Research, Twycross Zoo, Warwickshire, UK, 7–8 July 2005; pp. 175–187. [Google Scholar]
- Scott, G.; Hull, S.; Rollinson, D. Surface breaking behaviour in a population of captive rays Raja: The expression of a need to forage? Aquar. Sci. Conserv. 1998, 2, 161–169. [Google Scholar] [CrossRef]
- McLaughlin, R.; O’Gower, A. Life history and underwater studies of a heterodont shark. Ecol. Monogr. 1971, 41, 271–289. [Google Scholar] [CrossRef]
- Last, P.R.; Stevens, J.D.; Swainston, R.; Davis, G. Sharks and Rays of Australia; CSIRO: Melbourne, Australia, 2009.
- Ritar, A.J.; Pribadi, T.A. Hormonally Induced Spawning, Embryonic Development, and Larval Rearing of the Southern Temperate Banded Morwong, Cheilodactylus spectabilis. J. World Aquac. Soc. 2006, 37, 397–406. [Google Scholar] [CrossRef] [Green Version]
- Bissonette, J.A. Small sample size problems in wildlife ecology: A contingent analytical approach. Wildl. Biol. 1999, 5, 65–71. [Google Scholar]
- Ward, S.J.; Hosey, G. The need for a convergence of agricultural/laboratory and zoo-based approaches to animal welfare. J. Appl. Anim. Welf. Sci. 2020, 23, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Clubb, R.; Mason, G.J. Natural behavioural biology as a risk factor in carnivore welfare: How analysing species differences could help zoos improve enclosures. Appl. Anim. Behav. Sci. 2007, 102, 303–328. [Google Scholar] [CrossRef] [Green Version]
- de Azevedo, C.S.; Cipreste, C.F.; Young, R.J. Environmental enrichment: A GAP analysis. Appl. Anim. Behav. Sci. 2007, 102, 329–343. [Google Scholar] [CrossRef]
- Leonardi, R.; Buchanan-Smith, H.M.; Dufour, V.; MacDonald, C.; Whiten, A. Living together: Behavior and welfare in single and mixed species groups of capuchin (Cebus apella) and squirrel monkeys (Saimiri sciureus). Am. J. Primatol. 2010, 72, 33–47. [Google Scholar] [CrossRef]
- Clark, F.E.; Melfi, V.A. Environmental enrichment for a mixed-species nocturnal mammal exhibit. Zoo Biol. 2012, 31, 397–413. [Google Scholar] [CrossRef]
- Hogan, L.A.; Lisle, A.T.; Johnston, S.D.; Robertson, H. Non-invasive assessment of stress in captive numbats, Myrmecobius fasciatus (Mammalia: Marsupialia), using faecal cortisol measurement. Gen. Comp. Endocrinol. 2012, 179, 376–383. [Google Scholar] [CrossRef]
- Menargues, A.; Urios, V.; Mauri, M. Welfare assessment of captive Asian elephants (Elephas maximus) and Indian rhinoceros (Rhinoceros unicornis) using salivary cortisol measurement. Anim. Welf. 2008, 17, 305–312. [Google Scholar]
- Narayan, E.; Hero, J.-M.; Evans, N.; Nicolson, V.; Mucci, A. Non-invasive evaluation of physiological stress hormone responses in a captive population of the greater bilby Macrotis lagotis. Endanger. Species Res. 2012, 18, 279–289. [Google Scholar] [CrossRef] [Green Version]
- Rajagopal, T.; Archunan, G.; Sekar, M. Impact of zoo visitors on the fecal cortisol levels and behavior of an endangered species: Indian blackbuck (Antelope cervicapra L.). J. Appl. Anim. Welf. Sci. 2011, 14, 18–32. [Google Scholar] [CrossRef]
- Van der Weyde, L.; Martin, G.; Paris, M. Monitoring stress in captive and free-ranging African wild dogs (Lycaon pictus) using faecal glucocorticoid metabolites. Gen. Comp. Endocrinol. 2016, 226, 50–55. [Google Scholar] [CrossRef] [Green Version]
- Whitten, P.L.; Stavisky, R.; Aureli, F.; Russell, E. Response of fecal cortisol to stress in captive chimpanzees (Pan troglodytes). Am. J. Primatol. 1998, 44, 57–69. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Behaviour Type/Label | Description | Sampling Method |
|---|---|---|
| Active | Swimming below the water surface at a consistent/steady pace throughout the tank (not in a repetitive pattern). | Scan |
| Resting | Being motionless on the sand or structure, at rest | Scan |
| Perimeter Swimming | Moving against the perimeter of the tank in a horizontal position, meaning the ventral surface of the animal is in contact with the glass/tank walls. | Scan |
| Surface Swimming | Moving with a portion of body out of the water | Scan |
| Spy Hopping | Vigorously propelling the body out of the water in a vertical motion. At minimum, the head breaches the water but can extend to half the body length. Typically occurs at the edges of the tank. | All-occurrence |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lawrence, K.; Sherwen, S.L.; Larsen, H. Natural Habitat Design for Zoo-Housed Elasmobranch and Teleost Fish Species Improves Behavioural Repertoire and Space Use in a Visitor Facing Exhibit. Animals 2021, 11, 2979. https://doi.org/10.3390/ani11102979
Lawrence K, Sherwen SL, Larsen H. Natural Habitat Design for Zoo-Housed Elasmobranch and Teleost Fish Species Improves Behavioural Repertoire and Space Use in a Visitor Facing Exhibit. Animals. 2021; 11(10):2979. https://doi.org/10.3390/ani11102979
Chicago/Turabian StyleLawrence, Kristie, Sally L. Sherwen, and Hannah Larsen. 2021. "Natural Habitat Design for Zoo-Housed Elasmobranch and Teleost Fish Species Improves Behavioural Repertoire and Space Use in a Visitor Facing Exhibit" Animals 11, no. 10: 2979. https://doi.org/10.3390/ani11102979