
Colonization of Urban Habitats: Tawny Owl Abundance Is Conditioned by Urbanization Structure


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
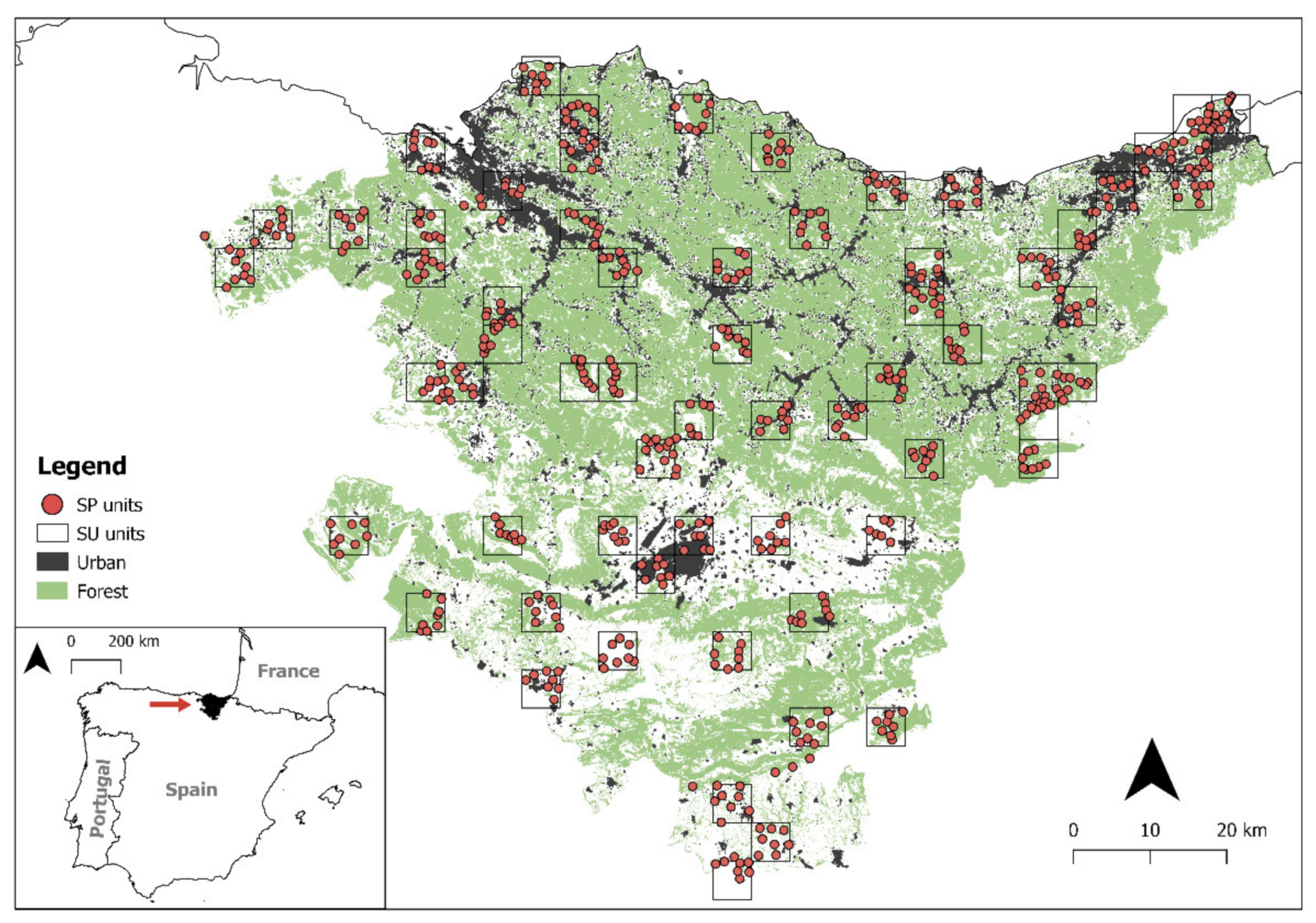
2.1. Study Area
2.2. Data Collection
2.3. Environmental Variables
2.4. Statistical Analysis
3. Results
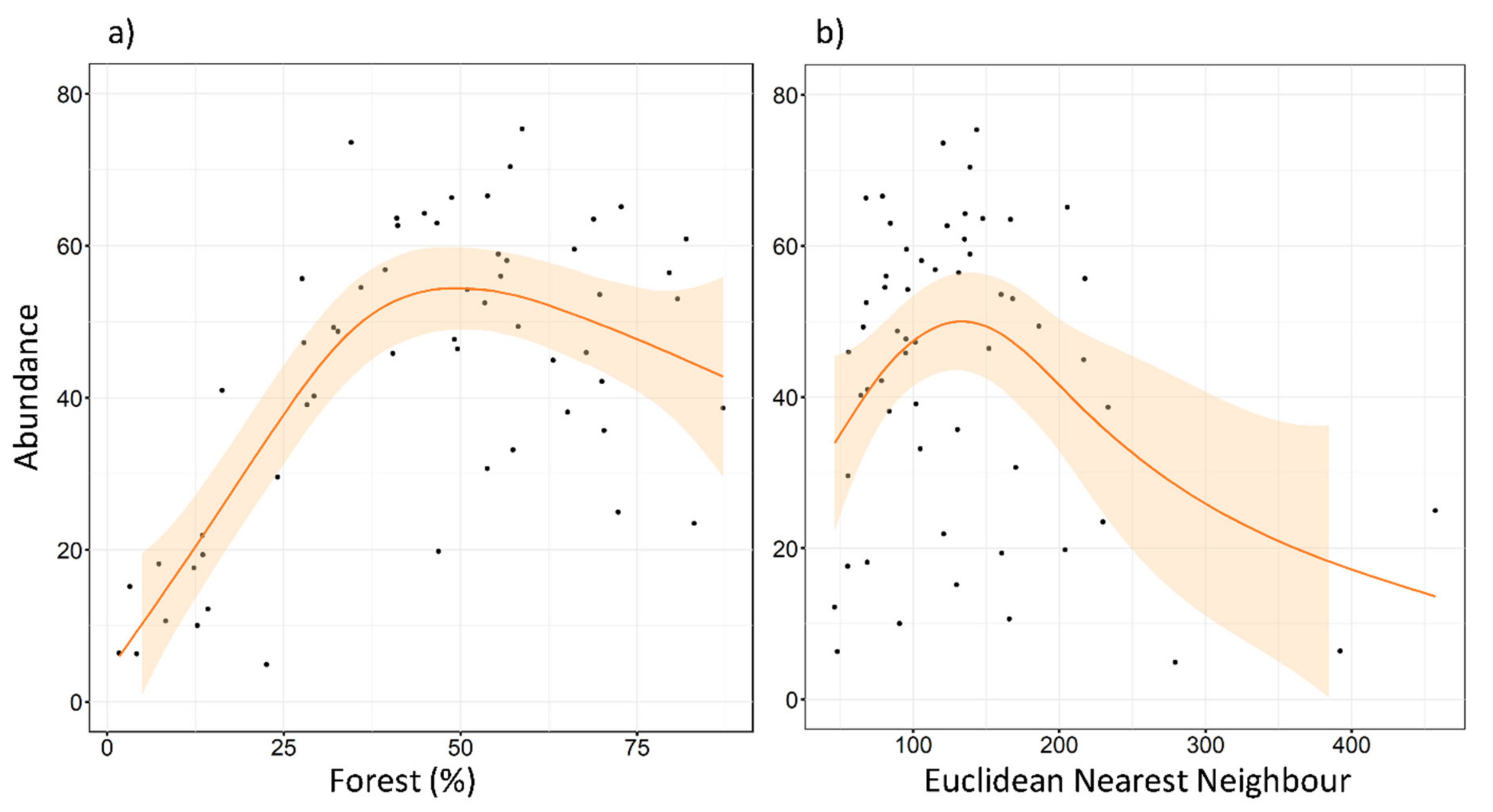
3.1. Local Scale
3.2. Landscape Scale
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (Month) | January | Feburary | March | April | May | June–July | Mean | |
|---|---|---|---|---|---|---|---|---|
| (Playback Sp.) | Strix aluco | Values | ||||||
| SU | Total SPs per SU | SPs Surveyed | SPs Surveyed | SPs Surveyed | SPs Surveyed | SPs Surveyed | SPs Surveyed | SPs Surveyed |
| 30TVN08NW | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 |
| 30TVN68SE | 8 | 5 | 6 | 6 | 6 | 8 | 6 | 6 |
| 30TVN78NW | 8 | 4 | 6 | 5 | 6 | 7 | 7 | 6 |
| 30TVN84NW | 8 | 8 | 8 | 6 | 6 | 8 | 8 | 7 |
| 30TVN88NW | 8 | 5 | 8 | 7 | 6 | 6 | 6 | 6 |
| 30TVN93NW | 8 | 8 | 8 | 6 | 6 | 8 | 7 | 7 |
| 30TVN96NE | 8 | 8 | 8 | 6 | 6 | 6 | 6 | 7 |
| 30TVN96NW | 8 | 8 | 7 | 8 | 6 | 6 | 6 | 7 |
| 30TVN98NW | 8 | 4 | 6 | 6 | 6 | 7 | 7 | 6 |
| 30TVN98SW | 8 | 4 | 8 | 6 | 5 | 6 | 6 | 6 |
| 30TVN99NW | 8 | 7 | 4 | 6 | 8 | 6 | 7 | 6 |
| 30TWN02NE | 8 | 8 | 8 | 7 | 6 | 6 | 6 | 7 |
| 30TWN03NE | 8 | 6 | 8 | 8 | 6 | 8 | 6 | 7 |
| 30TWN04NW | 8 | 8 | 8 | 6 | 6 | 6 | 6 | 7 |
| 30TWN07NW | 8 | 8 | 7 | 6 | 6 | 6 | 5 | 6 |
| 30TWN07SW | 8 | 8 | 8 | 7 | 6 | 6 | 4 | 7 |
| 30TWN09SW | 9 | 5 | 5 | 5 | 7 | 6 | 6 | 6 |
| 30TWN13SE | 8 | 8 | 8 | 6 | 8 | 8 | 8 | 8 |
| 30TWN14NE | 8 | 8 | 8 | 8 | 8 | 8 | 8 | 8 |
| 30TWN16NE | 8 | 8 | 8 | 6 | 5 | 5 | 5 | 6 |
| 30TWN16NW | 8 | 5 | 7 | 4 | 5 | 8 | 6 | 6 |
| 30TWN18NW | 8 | 5 | 6 | 6 | 6 | 7 | 6 | 6 |
| 30TWN18SE | 8 | 5 | 6 | 6 | 7 | 6 | 6 | 6 |
| 30TWN19NW | 8 | 8 | 8 | 7 | 7 | 7 | 7 | 7 |
| 30TWN21SE | 1 | 1 | 1 | 0 | 0 | 0 | 0 | 0 |
| 30TWN24NE | 8 | 7 | 5 | 3 | 6 | 6 | 6 | 6 |
| 30TWN24SW | 8 | 6 | 6 | 2 | 6 | 6 | 7 | 6 |
| 30TWN25NW | 12 | 10 | 9 | 8 | 6 | 4 | 6 | 7 |
| 30TWN26SE | 7 | 7 | 7 | 7 | 7 | 7 | 7 | 7 |
| 30TWN30NE | 8 | 8 | 8 | 6 | 6 | 6 | 8 | 7 |
| 30TWN30SW | 8 | 8 | 8 | 7 | 6 | 7 | 6 | 7 |
| 30TWN31NE | 3 | 3 | 3 | 0 | 0 | 0 | 0 | 1 |
| 30TWN31SW | 8 | 8 | 5 | 8 | 6 | 6 | 6 | 7 |
| 30TWN33SW | 8 | 7 | 6 | 5 | 6 | 5 | 6 | 6 |
| 30TWN34NE | 8 | 6 | 8 | 4 | 7 | 8 | 4 | 6 |
| 30TWN36SE | 8 | 6 | 6 | 6 | 6 | 6 | 6 | 6 |
| 30TWN37SW | 8 | 6 | 6 | 7 | 7 | 8 | 7 | 7 |
| 30TWN38SW | 8 | 7 | 7 | 7 | 6 | 8 | 6 | 7 |
| 30TWN39NE | 8 | 7 | 7 | 8 | 8 | 7 | 7 | 7 |
| 30TWN42SW | 8 | 8 | 8 | 6 | 6 | 6 | 6 | 7 |
| 30TWN43NW | 8 | 8 | 4 | 5 | 6 | 6 | 6 | 6 |
| 30TWN46SE | 8 | 6 | 6 | 7 | 6 | 7 | 4 | 6 |
| 30TWN48NW | 8 | 7 | 7 | 7 | 8 | 8 | 8 | 8 |
| 30TWN52SW | 8 | 8 | 8 | 6 | 6 | 6 | 6 | 7 |
| 30TWN54NW | 6 | 6 | 6 | 6 | 6 | 6 | 6 | 6 |
| 30TWN55NE | 8 | 6 | 6 | 7 | 6 | 6 | 5 | 6 |
| 30TWN56NW | 8 | 6 | 6 | 5 | 5 | 6 | 8 | 6 |
| 30TWN57NE | 8 | 4 | 6 | 6 | 6 | 6 | 8 | 6 |
| 30TWN58SE | 8 | 4 | 6 | 7 | 6 | 6 | 7 | 6 |
| 30TWN59SW | 8 | 6 | 4 | 5 | 6 | 6 | 8 | 6 |
| 30TWN67SW | 8 | 6 | 5 | 6 | 6 | 7 | 6 | 6 |
| 30TWN69SW | 8 | 6 | 5 | 5 | 6 | 6 | 7 | 6 |
| 30TWN75NW | 8 | 6 | 6 | 6 | 4 | 7 | 7 | 6 |
| 30TWN76NE | 8 | 5 | 7 | 6 | 4 | 7 | 7 | 6 |
| 30TWN76NW | 8 | 7 | 5 | 6 | 4 | 7 | 7 | 6 |
| 30TWN76SW | 8 | 5 | 7 | 6 | 5 | 7 | 7 | 6 |
| 30TWN77NE | 8 | 4 | 6 | 5 | 7 | 7 | 7 | 6 |
| 30TWN78NE | 8 | 4 | 6 | 7 | 7 | 7 | 5 | 6 |
| 30TWN78SW | 8 | 4 | 6 | 8 | 6 | 6 | 6 | 6 |
| 30TWN89NE | 8 | 6 | 5 | 6 | 6 | 6 | 6 | 6 |
| 30TWN89SW | 8 | 5 | 7 | 6 | 6 | 6 | 7 | 6 |
| 30TWN99NW | 8 | 6 | 5 | 5 | 4 | 7 | 8 | 6 |
| 30TWN99SW | 8 | 6 | 4 | 6 | 6 | 7 | 7 | 6 |
| 30TWP00NE | 8 | 7 | 7 | 8 | 8 | 8 | 7 | 8 |
| 30TWP10SW | 8 | 7 | 7 | 8 | 8 | 8 | 7 | 8 |
| 30TWP20SE | 8 | 8 | 7 | 7 | 8 | 8 | 8 | 8 |
| 30TWP90SE | 8 | 6 | 4 | 5 | 6 | 8 | 8 | 6 |
| 30TWP90SW | 8 | 6 | 4 | 6 | 4 | 7 | 8 | 6 |
| Total | 527 | 422 | 426 | 402 | 399 | 434 | 424 |
| Models | Formula | AIC | deltaAIC | wAIC |
|---|---|---|---|---|
| m.25 | REG + ALT + FOR + FOR2 + URB + CAI + CLU + ENN + SHAPE + FOR:URB + FOR2:URB + CLM:URB | 3235.88 | 0.00 | 0.28 |
| m.23 | REG + ALT + FOR + FOR2 + URB + CAI + CLU + SHAPE + NP + FOR:URB + FOR2:URB + CLM:URB + NP:URB | 3236.91 | 1.03 | 0.17 |
| m.29 | REG + ALT + FOR + FOR2 + URB + CAI + CLU + ENN + SHAPE + NP + FOR:URB + FOR2:URB + CLM:URB | 3239.94 | 1.06 | 0.16 |
| m.21 | REG + ALT + FOR + FOR2 + URB + CLU + ENN + SHAPE + NP + FOR:URB + FOR2:URB + CLM:URB + NP:URB | 3237.60 | 1.73 | 0.12 |
| m.24 | REG + ALT + FOR + FOR2 + URB + CAI + CLU + ENN + NP + FOR:URB + FOR2:URB + CLM:URB + NP:URB | 3238.09 | 2.21 | 0.092 |
| m.15 (sat) | REG + ALT + FOR + FOR2 + URB + CAI + CLU + ENN + SHAPE + NP + FOR:URB + FOR2:URB + CLM:URB + NP:URB | 3238.81 | 2.93 | 0.064 |
| m.22 | REG + ALT + FOR + FOR2 + URB + CAI + ENN + SHAPE + NP + FOR:URB + FOR2:URB + NP:URB | 3239.66 | 3.78 | 0.042 |
| m.27 | REG + ALT + FOR + FOR2 + URB + CAI + CLU + ENN + SHAPE + NP + FOR:URB + CLM:URB + NP:URB | 3240.10 | 4.22 | 0.034 |
| m.26 | REG + ALT + FOR + FOR2 + URB + CAI + CLU + ENN + SHAPE + NP + CLM:URB + NP:URB | 3241.18 | 5.30 | 0.020 |
| m.28 | REG + ALT + FOR + FOR2 + URB + CAI + CLU + ENN + SHAPE + NP + FOR:URB + FOR2:URB + NP:URB | 3241.41 | 5.53 | 0.018 |
| m.19 | REG + ALT + FOR + URB + CAI + CLU + ENN + SHAPE + NP + FOR:URB + CLM:URB + NP:URB | 3245.02 | 9.15 | 0.003 |
| Models | Formula | AIC | deltaAIC | wAIC |
|---|---|---|---|---|
| mo.11 | REG + ALT + ALT2 + FOR + FOR2 + URB + CAI + CAI2 + ENN + PAF + SHAPE + NP | 1234.39 | 0.00 | 0.22 |
| mo.8 | REG + ALT + ALT2 + FOR + FOR2 + CAI + CAI2 + CLU + ENN + PAF + SHAPE + NP | 1234.74 | 0.35 | 0.18 |
| mo.13 | REG + ALT + ALT2 + FOR + FOR2 + URB + CAI + CAI2 + CLU + ENN + SHAPE + NP | 1235.55 | 1.15 | 0.12 |
| mo.4 | REG + FOR + FOR2 + URB + CAI + CAI2 + CLU + ENN + PAF + SHAPE + NP | 1235.81 | 1.41 | 0.11 |
| mo.2 (sat) | REG + ALT + ALT2 + FOR + FOR2 + URB + CAI + CAI2 + CLU + ENN + PAF + SHAPE + NP | 1236.35 | 1.96 | 0.081 |
| mo.9 | REG + ALT + ALT2 + FOR + FOR2 + URB + CLU + ENN + PAF + SHAPE + NP | 1236.36 | 1.97 | 0.081 |
| mo.3 | ALT + ALT2 + FOR + FOR2 + URB + CAI + CAI2 + CLU + ENN + PAF + SHAPE + NP | 1236.78 | 2.39 | 0.065 |
| mo.5 | REG + ALT + FOR + FOR2 + URB + CAI + CAI2 + CLU + ENN + PAF + SHAPE + NP | 1236.81 | 2.41 | 0.065 |
| mo.10 | REG + ALT + ALT2 + FOR + FOR2 + URB + CAI + CLU + ENN + PAF + SHAPE + NP | 1237.73 | 3.33 | 0.041 |
| mo.15 | REG + ALT + ALT2 + FOR + FOR2 + URB + CAI + CAI2 + CLU + ENN + PAF + SHAPE | 1237.87 | 3.48 | 0.038 |
| mo.12 | REG + ALT + ALT2 + FOR + FOR2 + URB + CAI + CAI2 + CLU + PAF + SHAPE + NP | 1243.04 | 8.65 | 0.0029 |
| mo.7 | REG + ALT + ALT2 + FOR + URB + CAI + CAI2 + CLU + ENN + PAF + SHAPE + NP | 1243.99 | 9.60 | 0.0018 |
References
- McKinney, M.L. Urbanization, biodiversity, and conservation. BioScience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Sol, D.; González-Lagos, C.; Moreira, D.; Maspons, J.; Lapiedra, O. Urbanization tolerance and the loss of avian diversity. Ecol. Lett. 2014, 17, 942–950. [Google Scholar] [CrossRef]
- Lopucki, R.; Klich, D.; Kitowski, I.; Kiersztyn, A. Urban size effect on biodiversity: The need for a conceptual framework for the implementation of urban policy for small cities. Cities 2020, 98, 102590. [Google Scholar] [CrossRef]
- De Lima Filho, J.A.; Vieira, R.J.A.; de Souza, C.A.M.; Ferreira, F.F.; de Oliveira, V.M. Effects of habitat fragmentation on biodiversity patterns of ecosystems with resource competition. Physica A 2021, 564, 125497. [Google Scholar] [CrossRef]
- Hedblom, M.; Murgui, E. Urban Bird Research in a Global Perspective. In Ecology and Conservation of Birds in Urban Environments, 1st ed.; Murgui, E., Hedblom, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 3–10. [Google Scholar] [CrossRef]
- Shanahan, D.F.; Stronhbach, M.W.; Warren, P.S.; Fuller, R.A. The challenges of urban living. In Avian Urban Ecology, 1st ed.; Gil, D., Brumm, H., Eds.; OUP Oxford: Oxford, UK, 2014; pp. 3–20. [Google Scholar] [CrossRef]
- Santos, S.M.; Lourenco, R.; Mira, A.; Beja, P. Relative Effects of Road Risk, Habitat Suitability, and Connectivity on Wildlife Roadkills: The Case of Tawny Owls (Strix aluco). PLoS ONE 2013, 8, e79967. [Google Scholar] [CrossRef] [Green Version]
- Donázar, J.A.; Cortés-Avizanda, A.; Fargallo, J.A.; Margalida, A.; Moleón, M.; Morales-Reyes, Z.; Moreno-Opo, R.; Pérez-García, J.M.; Sánchez-Zapata, J.A.; Zuberogoitia, I.; et al. Roles of Raptors in a Changing World: From Flagships to Providers of Key Ecosystem Services. BioOne 2016, 63, 181–234. [Google Scholar] [CrossRef] [Green Version]
- Chace, J.F.; Walsh, J.J. Urban effects on native avifauna: A review. Landsc. Urban Plan. 2006, 74, 46–69. [Google Scholar] [CrossRef]
- Palomino, D.; Carrascal, L.M. Urban influence on birds at a regional scale: A case study with the avifauna of northern Madrid province. Landsc. Urban Plan. 2006, 77, 276–290. [Google Scholar] [CrossRef]
- Rullman, S.; Marzluff, J.M. Raptor presence along an urban-wildland gradient: Influences of prey abundance and land cover. J. Raptor Res. 2014, 48, 257–272. [Google Scholar] [CrossRef]
- Palomino, D.; Carrascal, L.M. Habitat associations of a raptor community in a mosaic landscape of Central Spain under urban development. Landsc. Urban Plan. 2007, 83, 268–274. [Google Scholar] [CrossRef]
- Martínez, J.A.; Zuberogoitia, I. Habitat references and causes of population decline for barn owls Tyto alba: A multiscale approach. Ardeola 2004, 51, 303–317. [Google Scholar]
- Ivajnšič, D.; Denac, D.; Denac, K.; Pipenbaher, N.; Kaligarič, M. The Scops owl (Otus scops) under human-induces environmental change pressure. Land Use Policy 2020, 99, 104853. [Google Scholar] [CrossRef]
- Solonen, T. Timing of breeding in rural and urban Tawny Owls Strix aluco in southern Finland: Effects of vole abundance and winter weather. J. Ornithol. 2014, 155, 27–36. [Google Scholar] [CrossRef]
- Solonen, T.; Lokki, H.; Sulkava, S. Diet and brood size in rural and urban Northern Goshawks Accipiter gentilis in southern Finland. Avian Biol. Res. 2019, 12, 3–9. [Google Scholar] [CrossRef]
- Anderson, D.L. Landscape Heterogeneity and Diurnal Raptor Diversity in Honduras: The Role of Indigenous Shifting Cultivation. Biotropica 2001, 33, 511–519. [Google Scholar] [CrossRef]
- Fröhlich, A.; Ciach, M. Nocturnal noise and habitat homogeneity limit species richness of owls in an urban environment. Environ. Sci. Pollut. Res. 2019, 26, 17284–17291. [Google Scholar] [CrossRef] [Green Version]
- Ranazzi, L.; Manganaro, A.; Salvati, L. Density fluctuations in urban population of Tawny Owl Strix aluco: A long-term study in Rome, Italy. Ornis Svecica 2002, 12, 63–67. [Google Scholar]
- Fröhlich, A.; Ciach, M. Noise pollution and decreased size of wooded areas reduces the probability of occurrence of Tawny Owl Strix aluco. Ibis 2018, 160, 634–646. [Google Scholar] [CrossRef]
- Redpath, S.M. Habitat Fragmentation and the Individual: Tawny Owl Strix aluco in Woodland Patches. J. Anim. Ecol. 1995, 64, 652–661. [Google Scholar] [CrossRef]
- Sunde, P. What do we know about territorial behaviour and its consequences in Tawny Owls? In Ecology and Conservation of European Forest-Dwelling Raptors, 1st ed.; Zuberogoitia, I., Martínez, J.E., Eds.; Departamento de Agricultura de la Diputación Foral de Bizkaia: Bilbao, Spain, 2011; pp. 253–260. [Google Scholar]
- Martínez, J.A.; Serrano, D.; Zuberogoitia, I. Predictive models of hábitat preferences for the Eurasian Eagle Owl Bubo bubo: A multiscale approach. Ecography 2003, 26, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Euskalmet: Agencia Vasca de Meteorología. El Clima en Euskadi: Zonas Climáticas; Gobierno Vasco: Vitoria-Gasteiz, Spain. Available online: https://www.euskalmet.euskadi.eus/clima/euskadi/ (accessed on 15 March 2019).
- Zuberogoitia, I.; Martínez, J.E.; González-Oreja, J.A.; González de Buitrago, C.; Belamendia, G.; Zabala, J.; Laso, M.; Pagaldai, N.; Jiménez-Franco, M.V. Maximizing detection probability for effective large-scale nocturnal bird monitoring. Divers. Distrib. 2020, 26, 1034–1050. [Google Scholar] [CrossRef]
- Zuberogoitia, I.; Martínez, J.E.; González-Oreja, J.A.; González de Buitrago, C.; Belamendia, G.; Zabala, J.; Laso, M.; Pagaldai, N.; Jiménez-Franco, M.V. Data from: Maximizing detection probability for effective large-scale nocturnal bird monitoring. Divers. Distrib. 2020. Dryad, Dataset. [Google Scholar] [CrossRef]
- Zuberogoitia, I.; Campos, L.F. Censusing owls in large areas: A comparison between methods. Ardeola 1998, 45, 47–53. [Google Scholar]
- Zuberogoitia, I.; Martínez, J.A. Methods for surveying Tawny Owl Strix aluco populations in large areas. Biota 2000, 1, 137–146. [Google Scholar]
- Zuberogoitia, I.; Burgos, G.; González-Oreja, J.A.; Morant, J.; Martínez, J.E.; Zabala, J. Factors affecting spontaneous vocal activity of Tawny Owls Strix aluco and implications for surveying large areas. Ibis 2019, 161, 495–503. [Google Scholar] [CrossRef]
- MacKenzie, D.I.; Nichols, J.D.; Lachman, G.B.; Droege, S.; Royle, J.A.; Langtimm, C.A. Estimating site occupancy rates when detection probabilities are less than one. Ecology 2002, 83, 2248–2255. [Google Scholar] [CrossRef]
- Zabala, J.; Zuberogoitia, I.; Martínez-Climents, J.A.; Martínez, J.E.; Azkona, A.; Hidalgo, S.; Iraeta, A. Occupancy and abundance of Little Owl Athene noctua in an intensively managed forest area in Biscay. Ornis Fennica 2006, 83, 97–107. [Google Scholar]
- Zuberogoitia, I.; Zabala, J.; Martínez, J.E. Bias in little owl population estimates using playback techniques during surveys. Anim. Biodivers. Conserv. 2011, 34, 395–400. [Google Scholar]
- Fiske, I.; Chandler, R. Unmarked: An R package for fitting hierarchical models of wildlife occurrence and abundance. J. Stat. Softw. 2011, 43, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Kéry, M.; Royle, J.A.; Schmid, H. Modeling avian abundance from replicated count using binomial mixture models. Ecol. Appl. 2005, 15, 1450–1461. [Google Scholar] [CrossRef]
- Royle, J.A. N-Mixture Models for Estimating Population Size from Spatially Replicated Counts. Biometrics 2004, 60, 108–115. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 23 July 2021).
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Giam, X.; Olden, J.D. Quantifying variable importance in a multimodel inference framework. Methods Ecol. Evol. 2016, 7, 388–397. [Google Scholar] [CrossRef]
- Krener, P.; Hamstead, Z.; Haase, D.; McPhearson, T.; Frantzeskaki, N.; Andersson, E.; Kabisch, N.; Larondelle, N.; Rall, E.L.; Voigt, A.; et al. Key insights for the future of urban ecosystem services research. Ecol. Soc. 2016, 21, 29. [Google Scholar] [CrossRef] [Green Version]
- Michel, V.T.; Jiménez-Franco, M.V.; Naef-Daenzer, B.; Grüebler, M.U. Intraguild predator drives forest edge avoidance of a mesopredator. Ecosphere 2016, 7, e01229. [Google Scholar] [CrossRef] [Green Version]
- Ranazzi, L.; Manganaro, A.; Ranazzi, R.; Salvati, L. Woodland cover and Tawny Owl Strix aluco density in a Mediterranean urban area. Biota 2000, 1, 27–34. [Google Scholar]
- Burgos, G.; Zuberogoitia, I. A telemetry study to discriminate between home range and territory size in Tawny Owls. Bioacustics 2018, 29, 109–121. [Google Scholar] [CrossRef]
- Rodewald, A.D.; Shustack, D.P. Consumer resource matching in urbanizing landscapes: Are synanthropic species over-matching? Ecology 2008, 89, 515–521. [Google Scholar] [CrossRef] [Green Version]
- Sacchi, R.; Gentilli, A.; Razzetti, E.; Barbieri, F. Effects of building features on density and flock distribution of Feral Pigeons Columba livia var. domestica in an urban environment. Can. J. Zool. 2002, 80, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Millsap, B.A. Survival of Florida Burrowing Owls along an urban-development gradient. J. Raptor Res. 2002, 36, 3–10. [Google Scholar]
- López-Peinado, A.; Lis, A.; Perona, A.M.; López-López, P. Habitat preferences of the tawny owl (Strix aluco) in a special conservancy area of eastern Spain. J. Raptor Res. 2020, 54, 402–413. [Google Scholar] [CrossRef]
- Whytock, R.C.; Fuentes-Montemayor, E.; Watts, K.; Macgregor, N.A.; Call, E.; Mann, J.A.; Park, K.J. Regional land-use and local management create scale-dependent ‘landscapes of fear’ for a common woodland bird. Landsc. Ecol. 2020, 35, 607–620. [Google Scholar] [CrossRef] [Green Version]
- Alaniz, A.J.; Carvajal, M.A.; Fierro, A.; Vergara-Rodríguez, V.; Toledo, G.; Ansaldo, D.; Moreira-Arce, D.; Rojas-Osorio, A.; Vergara, P.M. Remote-sensing estimates of forest structure and dynamics as indicators of habitat quality for Magellanic woodpeckers. Ecol. Indic. 2021, 126, 107634. [Google Scholar] [CrossRef]
- Mak, B.; Francis, R.A.; Chadwick, M.A. Living in the concrete jungle: A review and socio-ecological perspective of urban raptor habitat quality in Europe. Urban Ecosyst. 2021, 1–21. [Google Scholar] [CrossRef]
- Gryz, J.; Krauze-Gryz, D. Influence of habitat urbanization on time of breeding and productivity of tawny owl (Strix aluco). Pol. J. Ecol. 2018, 66, 153–161. [Google Scholar] [CrossRef]
- Møller, A.P.; Díaz, M. Avian preference for close proximity to human habitation and its ecological consequences. Curr. Zool. 2018, 64, 623–630. [Google Scholar] [CrossRef]
- European Commision. European Commision, Europe 2020: A Trategy for Smart, Sustainable and Inclusive Growth. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:52010DC2020&from=EN (accessed on 23 July 2021).
- Gulsrud, N.M.; Ostoić, S.K.; Faehnle, M.; Maric, B.; Paloniemi, R.; Pearlmutter, D.; Simson, A.J. Challenges to Governing Urban Green Infrastructure in Europe—The Case of the European Green Capital Award. In The Urban Forest, 1st ed.; Pearlmutter, D., Calfapietra, C., Samson, R., O’Brien, L., Ostoić, S.K., Sanesi, G., Alonso del Amo, R., Eds.; Springer: Cham, Switzerland, 2017; Volume 7, pp. 235–258. [Google Scholar] [CrossRef]
- Sergio, F.; Newton, I.; Marchesi, L. Top predators and biodiversity. Nature 2005, 436, 192. [Google Scholar] [CrossRef]
- Lodenius, M.; Solonen, T. The use of birds of prey as indicators of metal pollution. Ecotoxicology 2013, 22, 1319–1334. [Google Scholar] [CrossRef] [PubMed]






| Habitat | SP (Mean % ± SD) | SU (Mean % ± SD) | Study Area (%) |
|---|---|---|---|
| Forest | 42.8 ± 26.3 | 45.1 ± 23.1 | 54.6 |
| Urban | 6 ± 11.7 | 8.2 ± 11.7 | 3.1 |
| Variables | Contraction | Description | Type | Range | Unit | Scale |
|---|---|---|---|---|---|---|
| Region | REG | Climatic region (Cantabric, Subcantabric, Mediterranean) | Binary | 0–1–2 | None | LO + LA |
| Altitude | ALT | Altitude above sea level | Continuous | 0–1500 | Meters | LO + LA |
| Forest | FOR | Forested area percentage | Continuous | 0–100 | % | LO + LA |
| Urban | URB | Urbanized area percentage | Continuous | 0–100 | % | LO + LA |
| Mean core area index | CAI | CAI is the percentage that the core area (interior area) takes in a patch. The mean value of the CAI of all the urban patches in each SP or SU is calculated. | Continuous | 0–100 | % | LO + LA |
| Clumpiness index | CLU | Describes how the entire group of urban patches is distributed. Equal to −1 for maximally disaggregated, 0 for randomly distributed, and 1 for maximally aggregated urban patches. | Continuous | −1–1 | None | LO + LA |
| Euclidean nearest mean neighbor | ENN | ENN measures the distance between one urban patch and its nearest urban patch (does not take into account the whole patch group). It is calculated as the mean of ENN of all the urban patches in each SP or SU. | Continuous | >0 | Meters | LO + LA |
| Shape index | SHAPE | SHAPE describes the ratio between the actual perimeter of the urban patch and its hypothetical minimum perimeter. The mean SHAPE value of all urban patches is calculated. | Continuous | ≥1 | None | LO + LA |
| Number of patches | NP | The number of urban patches | Discrete | ≥1 | LO + LA | |
| Perimeter Area Fractal Dimension | PAF | Describes the complexity of an urban patch. Approaches 1 for those with simple shapes (i.e., like a square) and approaches 2 for those that are very irregular. | Continuous | 1 ≤ PAF ≤ 2 | LA |
| Parameters NB | Estimate | SE (Estimate) | Lower 95% CI | Upper 95% CI | RVI |
|---|---|---|---|---|---|
| Local scale: 1 km2 | |||||
| Intercept | 4.77 | 0.44 | 3.91 | 5.63 | |
| REG | −0.76 | 0.11 | −0.98 | −0.54 | 0.99 |
| ALT | 0.45 | 0.08 | 0.28 | 0.61 | 0.99 |
| FOR | 0.07 | 0.08 | −0.08 | 0.23 | 0.99 |
| FOR2 | −0.25 | 0.08 | −0.40 | −0.09 | 0.99 |
| URB | 1.01 | 0.39 | 0.24 | 1.77 | 0.99 |
| CAI | 0.07 | 0.07 | −0.06 | 0.20 | 0.89 |
| CLU | −0.51 | 0.26 | −1.01 | 0.00 | 0.99 |
| ENN | 0.02 | 0.05 | −0.08 | 0.12 | 0.83 |
| SHAPE | −0.08 | 0.06 | −0.21 | 0.05 | 0.92 |
| NP | −0.08 | 0.09 | −0.25 | 0.10 | 0.68 |
| FOR × URB | −0.06 | 0.16 | −0.37 | 0.25 | 0.99 |
| FOR2 × URB | −0.22 | 0.11 | −0.44 | −0.01 | 0.96 |
| URB × CLU | −0.92 | 0.48 | −1.85 | 0.01 | 0.97 |
| URB × NP | 0.03 | 0.06 | −0.08 | 0.14 | 0.51 |
| Landscape scale: 25 km2 | |||||
| Intercept | 3.8 | 1.06 | 1.72 | 5.87 | |
| REG | −0.14 | 0.14 | −0.42 | 0.13 | 0.89 |
| ALT | 0.06 | 0.11 | −0.16 | 0.28 | 0.90 |
| ALT2 | −0.13 | 0.07 | −0.26 | 0.01 | 0.86 |
| FOR | 0.22 | 0.08 | 0.06 | 0.37 | 1 |
| FOR2 | −0.21 | 0.08 | −0.36 | −0.06 | 1 |
| URB | 0.07 | 0.1 | −0.12 | 0.26 | 0.84 |
| CAI | 0.31 | 0.22 | −0.12 | 0.75 | 0.86 |
| CAI2 | −0.19 | 0.13 | −0.43 | 0.06 | 0.78 |
| CLU | 0 | 0.09 | −0.18 | 0.18 | 0.80 |
| ENN | −0.6 | 0.22 | −1.03 | −0.17 | 1 |
| PAF | 0.08 | 0.09 | −0.11 | 0.26 | 1 |
| SHAPE | −0.53 | 0.14 | −0.8 | −0.25 | 1 |
| NP | −0.13 | 0.08 | −0.29 | 0.03 | 0.96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pagaldai, N.; Arizaga, J.; Jiménez-Franco, M.V.; Zuberogoitia, I. Colonization of Urban Habitats: Tawny Owl Abundance Is Conditioned by Urbanization Structure. Animals 2021, 11, 2954. https://doi.org/10.3390/ani11102954
Pagaldai N, Arizaga J, Jiménez-Franco MV, Zuberogoitia I. Colonization of Urban Habitats: Tawny Owl Abundance Is Conditioned by Urbanization Structure. Animals. 2021; 11(10):2954. https://doi.org/10.3390/ani11102954
Chicago/Turabian StylePagaldai, Nerea, Juan Arizaga, María V. Jiménez-Franco, and Iñigo Zuberogoitia. 2021. "Colonization of Urban Habitats: Tawny Owl Abundance Is Conditioned by Urbanization Structure" Animals 11, no. 10: 2954. https://doi.org/10.3390/ani11102954